

 PDF
PDF Citation
Citation Print
Print


Introduction
Mesenchymal stem cells (MSCs) are multipotent, self-renewable, and spindle shape cells (1) which were firstly isolated from guinea-pig bone marrow and spleen by Friedenstein et al. in 1970 (2). Now, MSCs can be isolated from cord blood, umbilical cord, bone marrow, adipose tissue, and many other tissues (3, 4). The single connecting feature of all these tissues is the presence of perivascular space around the blood vessels where pericytes come off and differentiate into MSCs after vessels got inflamed or injured. According to their common origin, pericytes and MSCs both are negative for CD34, CD45, CD56 and positively express CD29, CD44, CD73, CD90, CD105, CD146 (5-7). MSCs are functionally defined based on their capacity to produce molecules that provide a regenerative and immunoregulatory milieu. Moreover, these multipotent cells can differentiate into a closely related family of cells in vitro or following transplantation and subsequently recover the damages. However, recent findings have reported that MSCs do not significantly differentiate into other lineages in vivo under normal condition (8). Consequently in a new paradigm, Arnold Caplan coined “Medical Signaling Cell” for MSCs to precisely express that mesenchymal stem cells dock at the sites of injury and produce regenerative and immunoregulatory components (9, 10). Afterwards, a great number of clinical trials have exploited MSC-based modalities in clinical conditions such as Type 1 diabetes mellitus, Rheumatoid Arthritis (RA), Multiple Sclerosis (MS), Crohn’s disease, Lupus, kidney transplant, Graft versus Host Disease (GvHD) and many cancers (clinicaltrials.gov).
Unlike other stem cells (embryonic stem cells as well as induced pluripotent stem cells), MSCs have a dynamic immunomodulatory profile which lies on their ability to sense the injury or inflammation and then switch to the required response (11). For instance, MSCs produce transforming growth factor beta (TGF-β), indoleamine 2,3-dioxygenase (IDO), nitric oxide (NO), soluble HLA-G, and prostaglandin E2 (PGE2) in response to high concentration of interferon gamma (IFNγ), tumor necrosis factor alpha (TNF-α), and toll like receptor 3 (TLR3) agonists (11-13). This reaction activates CD4+CD25hiFoxP3+ regulatory T cells (Tregs) while inhibits natural killer cell and T cell responses (11-13). In contrast, low level of inflammatory agents and TLR4 agonists, decrease IDO, NO, PGE2 production and increase expression of pro-inflammatory cytokines (CXCL9, CXCL10, MIP-1α, MIP-1β, CCL5), indicating their double-edged sword function within the immune system (14). Accumulating investigations have demonstrated that these factors are mainly delivered through direct secretions of soluble molecules and release of extracellular vesicles particularly exosomes.
Go to : 
Exosomes
As shown in Table 1 (15-20), exosomes are one of the nanoscale cell-derived extracellular vesicles (EV) with 30∼100 nm diameter (15). Exosomes were first thought to be required for discarding unnecessary proteins out of the cells and termed as “garbage cans” (21). Later, they were introduced as gulps of cytosol delivering bioactive molecules especially proteins and small RNAs (like microRNAs) in intercellular communications. Recent investigation have shown that exosomes transfer bioactive molecules among cells and therefore is able to play an essential role in pathological conditions including Alzheimer’s disease (22), transmissible spongiform encephalopathies (23), Parkinson’s disease (24), and amyotrophic lateral sclerosis (25) or could be employed as nanoscale platforms for drug delivery.
Table 1
Commonalities and diversities of EVs
| Features\Type of vesicle | Exosome | Ectosome or Microvesicle or Microparticle | Apoptotic body |
|---|---|---|---|
| The most frequent biomarkers | CD9, CD63, CD81 (15), Alix (16), Tsg101 (16, 17) | No common marker (18) | Annexin V, Phosphatidylserine, Thrombospondin, C3b (19) |
| Size (nm) | 30∼100 | 100∼1000 | 500∼4000 |
| Stemed from | Intraluminal microvesicles which are originated from endosomal pathway, pass through plasma membrane via exocytosis | Plasma membrane budding of activated cells including human neutrophils, tumor cells, erythrocytes, and polymorphonuclear leukocytes (20) | Plasma membrane of apoptotic cells containing cell fragments |
![]()
Constitutive shedding of exosomes is mostly reported for immune cells (i.e. MSCs, lymphocytes, mast cells and dendritic cells) and non-immune cells particularly tumor cells (26, 27). Additionally, interaction between T lymphocytes and antigen presenting cells strongly induce the release of the exosomes bearing TCR/CD3/zeta complex (28) and MHC II-peptide (29), respectively. It was lately demonstrated that in hypoxic preconditioning and presence of lipopolysaccharide, MSCs significantly enhance the release of exosomes which shuttle anti-inflammatory agents to polarize macrophages into the M2 phenotype (30, 31).
Generally, extracellular vesicles affect the neighboring cells via two different strategies: 1) fusion dependent 2) fusion independent. The first mechanism takes place when vesicle is taken up by a recipient cell. There are two types of fusions: 1) direct fusion of the extracellular vesicle with the plasma membrane of recipient cell that requires receptor-ligand interactions 2) Back fusion which confers to the endocytosis of vesicle by the recipient cell and subsequently incorporation of vesicle with endosome’s membrane (32, 33). In fusion independent manner, the vesicle and recipient cells interact with each other, like when exosomes bearing MHC-peptide are being specifically attached to the T cell receptor (TCR) on T cells (34).
Go to :
MSC-Derived Exosome Therapy: Cons and Pros
As shown in Table 2 (35-46), recent findings depicted that MSC-derived exosomes deliver various cytoplasmatic constituents of the MSC secretomes, relevant to the stemness, angiogenesis, and particularly inflammatory factors. They were first used in 2010 for regeneration of tissues in a mouse model of myocardial ischemia/reperfusion injury (47) and are extensively on investigations at present. As a cell-free therapeutic tools provided a potential opportunity to unlock existing boundaries of cell therapy for autoimmune diseases. Exosomes introduce several advantages including capacity to cross the barriers (blood-brain barrier, capillaries) (48) and small enough to evade from being cleared by reticuloendothelial system (RES) (49). In addition to their general characteristics, MSC-derived exosomes are normally hypo-immunogenic due to the lack of MHC-II and low expression of MHC-I similar to their parental cells (50). It has also been revealed that MSCs can produce higher amount of exosomes compared with human cell lines like human embryonic kidney (HEK) and human acute monocytic leukemia (THP-1); Therefore, MSCs appear to be one of the best sources for high amount of immunoregulatory exosomes (51, 52).
Table 2
The content of mesenchymal-derived exosomes
| Component(s) | Function(s) | Source(s) | Reference(s) |
|---|---|---|---|
| hCAP-18/LL-37 | Antibacterial | hMSC | (35) |
| SOX2, Nanog, POU5F1A/B | Critical role in multipotency of MSC | hMSC | (36) |
| Transforming Growth Factor β1 (TGF-β1) | Inhibits lymphocyte proliferation and differentiation | hMSC | (37, 38) |
| Interleukin 6 (IL-6) | IL-10 and inducer | hMSC | (38) |
| Acute phase reaction enhancer | |||
| Interleukin 10 (IL-10) | T cell proliferation suppressor | pMSC | (39) |
| Regulatory T cell inducer | |||
| Hepatocyte Growth Factor (HGF) | Decrease antigen presenting capacity of Dendritic cells and suppress T cell response | hMSC | (40, 41) |
| PGE-2 | T cell proliferation suppressor | hMSC | (42) |
| NK cell proliferation suppressor | |||
| miR-155 | Pro and anti-inflammatory miRNA based depending on stimulant involved | mMSC | (43) |
| miR-146a | Anti-inflammatory miRNA | hMSC | (36) |
| miR-181c | Negatively affects NF-κB importin on nuclear membrane | hMSC | (44) |
| miR-17 | Anti NLRP3 inflammasome signaling pathway | hMSC, mMSC | (36, 45) |
| Targets E-selectin | |||
| miR-21 | Anti TLR4 signaling pathway | hMSC | (36, 46) |
| miR-10a | Endothelial inflammation and senescence regulator | hMSC | (36) |
| miR-92a | Targets MKK4 kinase upon TLR signaling pathway in macrophages | hMSC | (36) |
| miR-99b | Downregulates expression of IL-1β, IL-6, and IL-12 by targeting either TNF-α and TNF-α superfamily member 4 receptor genes | hMSC | (36) |
![]()
Despite therapeutic advantages of MSC-derived exosomes, quantification of their protein profile and more importantly durability of cryopreserved exosomes as well as their in vivo tracking are still effortful. Regarding frozen exosomes, Cosenza et al. (53) analyzed functions of cryopreserved MSC-derived exosomes compared to freshly isolated exosomes and demonstrated that storage destabilize the integrity of membrane, leading to leakage of its content, aggregation of vesicles, and decrease in their immunosuppressive capacity. MSC-derived exosomes obviously express both the common MSC markers, such as CD44, CD90, and CD73 and markers of exosomes, CD9, CD63, and CD81 (51). These markers are used to track exosomes especially for drug delivery. Alternatively, cell tracing dyes can also help to chase exosomes in vivo. For instance, intravenous (i.v.) administration of Vybrant Cell Tracer DiD- or DiI-labeled exosomes to the mouse model of acute kidney injury, were used to monitor accumulation of MSC-derived exosomes. In this investigation, exosomes were mostly found in kidney, spleen, liver, and lung as filtering organs of the body (54). Apart from utilization of fluorescent molecules to chase organ localization of EVs, lower sensitivity of fluorophores negatively affect pharmacokinetic analysis of their applications (especially for quantification of exosome accumulated in a target tissue) (55). Radiolabeling with iodine-125 and bioluminescence emitted from luciferase provide a more practical solution for whole-body imaging of exosome-treated animals with significantly higher sensitivity of detection (56, 57). Also, stability of B16-BL6 melanoma cell-derived exosomes in blood circulation were evaluated using exosomes expressing a fusion protein of Gaussia luciferase-Lactadherin, a reporter protein attached to one of the exosome’s trophic protein, respectively. The results indicated exosomal half-life of about 2 minutes following i.v. injection. This finding suggests short half-life of exosomes is because of their fast uptake by liver and spleen resident macrophages (56).
Go to :
MSC-Derived Exosome Therapy for Autoimmune Diseases
Utilization of the MSC-derived exosomes for autoimmune diseases therapy is at early stages. Most of studies are focused on immunomodulatory effects of these small vesicles in animal models. Here we will discuss the roles of these exosomes in the treatment of autoimmune diseases.
Go to :
Multiple Sclerosis
Multiple Sclerosis (MS) is a chronic autoimmune disease characterized by neuroinflammation and demyelination in the central nervous system (CNS) (58). Numerous studies have revealed activation and recruitment of autoaggressive CD4+ and CD8+ T cells into the CNS as the main pathogenic players of MS (59, 60). Disease-modifying treatments (DMTs) especially IFN-β are commonly prescribed to treat the affected individuals (61, 62). Despite their beneficial impacts, DMTs increase the risk of adverse effects including lymphopenia, lymphadenopathy, dyspnea and lipoatrophy (63-65). Hence, identification and evaluation of alternative approaches with lower side effects are necessary.
We have already demonstrated that MSCs and their activity have been altered after DMT consumption in relapsing-remitting MS patients (66). Indeed, patients own stem cells may have more deficiencies which not efficiently prevent for MS initiation and progression (67). However, MSCs can differentiate into neural cells and release neurotrophic molecules, indicating their strong neuroprotective effects for treating MS (68). Accumulating evidence has shown that MSCs can regulate autoreactive cells in a paracrine manner through secretion of inhibitory cytokines or molecules as well (69-71). MSC-derived exosomes can also induce peripheral tolerance toward autoreactive T cells via transferring the tolerogenic molecules to the autoreactive immune cells.
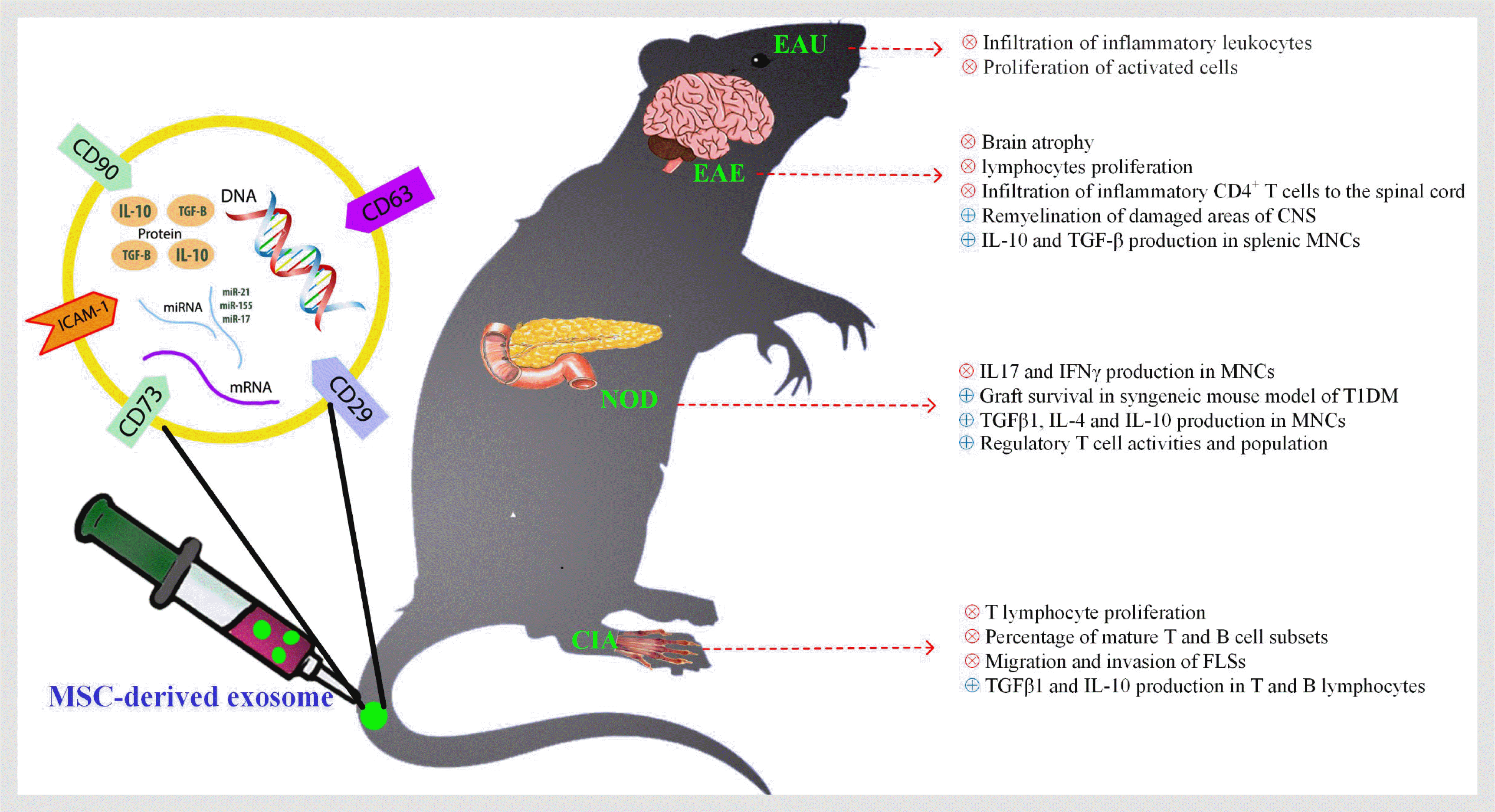
Since exosomes are lipid-bound nano-vesicles, they can cross the blood-brain barrier. Therefore, incubation of exosomes in presence of neurotrophic and remyelination factors elevate their therapeutic profits. Additionally, expression of receptor(s) with ability to recognize neural markers in the MSCs increase efficiency of the delivery system. In this case, Zhuang et al. (72) have administered exosomes bearing anti-inflammatory compounds such as curcumin to the EAE mouse and demonstrated that clinical symptoms of the diseases were improved.
In 2012, Mokarizadeh et al. (73) showed that MSC-derived extracellular vesicles exert immunomodulatory effects on splenic mononuclear cells (MNCs) of EAE mouse. These small vesicles not only reduced autoreactive lymphocytes proliferation, but also potentially induced MNCs to secrete significant amount of IL-10. They also demonstrated that MSC-derived vesicles contain PD-L1, galectin-1, and TGF-β, which help them with their immunoregulatory functions. This investigation proposed that MSC-derived vesicles including exosomes are capable of transferring anti-inflammatory molecules into autoreactive cells and re-inducing a self-tolerance. Later in 2018, Laso-García et al. (74) revealed that i.v. administration of MSC-derived exosomes to a progressive model of MS, Theiler’s murine encephalomyelitis virus (TMEV)-induced demyelinating disease, recovered the disabilities observed before treatment. They indicated that administrated MVs prompt remyelination, reduce the brain atrophy, and infiltration of inflammatory CD4+ T cells to the spinal cord of TMEV-infected mice.
Recently, Li et al. (75) demonstrated that MSC-derived exosomes polarize microglia cells mainly into the M2 phenotype and consequently alleviate clinical scores of EAE. In this study, the balance of M1/M2 in EAE rates significantly deviated toward M2 phenotype and their cytokines profile (IL-10 and TGF-β), while frequency and activity of M1 cells were decreased. Interestingly for the first time, Hosseini Shamili et al. (76) bioconjugated the MSC-derived exosomes to an aptamer which targets oligodendrocyte markers. The armed exosome improved proliferation of oligodendroglia cell line (OLN93) in vitro and reduced both inflammatory responses and demyelinated lesions in the CNS of EAE mice. These methodologies may provide a new perspective toward indirect applications of MSCs in the treatment of MS.
Go to :
Rheumatoid Arthritis
Rheumatoid arthritis (RA) is an inflammatory disease with unknown etiology that causes articular destruction and subsequently functional loss in joints. RA immunopathogenesis is characterized by disordered innate and adaptive immunity that result in immune complex-mediated complement activation, autoreactive lymphocyte response against self-antigens, and also dysregulated cytokine networks (77, 78).
As with MS, there are various biological disease-modifying anti-rheumatic drugs (bDMARDs) available to control the symptoms of RA. For example, blockade of proinflammatory cytokines including GM-CSF, TNF-α, and IL-6 with antibodies or soluble decoy receptors have been mainly utilized for patients suffering from RA for many years (79). Along with beneficial effects of immunosuppressive and immunomodulatory drugs, they present various side effects including nausea, liver problems, lymphopenia, high risk of infection, and etc. (79). There is indeed a crucial need for new treatment modules with fewer side effects. MSC-derived exosomes have been considered to exert immunosuppressive activities, supporting their application in rheumatic diseases. Although the use of exosomes derived from various origins have been studied extensively to date (80, 81), to date, the role of MSC-derived exosomes is still in the beginning.
In 2018, Cosenza et al. (53) showed that MSC-derived exosomes reveal anti-inflammatory impacts on T and B lymphocytes in collagen-induced arthritis (CIA) mice model. In this study, MSCs-derived exosomes effectively inhibit T lymphocyte proliferation in a dose-dependent manner and decreased the percentage of mature T and B cell subsets. Interestingly, exosomes increased regulatory T cell populations much better than MSCs and MSC-derived microvesicles. Further investigations confirmed that microvesicles derived from MSCs are less effective to induce TGF-β and IL-10 production in T and B lymphocytes compared to MSCs alone (82). In 2018, Chen et al. (83) demonstrated that modified MSC-derived exosomes which contain miRNA-150, target the matrix metalloproteinase 14 (MMP14) and thereafter, decrease migration and invasion of fibroblast-like synoviocytes (FLS). They also indicated that injection of these engineered exosomes to the CIA mouse can effectively reduce the hind paw thickness and the clinical arthritic scores of animals. Another study evaluated beneficial impacts of human umbilical cord mesenchymal stem cells-derived exosomes (hUCMSC-derived exosomes) on bone destruction in CIA rats. Interestingly, the treatment reduced recruitment of inflammatory cell to the joints and prevent from joint synovial hyperplasia. The authors proposed that the underlying mechanism could be due to regulating of the imbalance of RANKL/OPG. To be clear, they showed that RANKL was decreased in serum and synovial tissues of the CIA rats, while the treatment raised the OPG concentration (84). In consensus with this study, another group demonstrated that intra-articular injection of hUCMSC-derived exosomes significantly decreased autoreactive infiltering chemokines CCL2 and CXCL12 in serum and synovial fluid (85).
Go to :
Type 1 Diabetes Mellitus (Insulin-Dependent Diabetes Mellitus)
Type 1 diabetes mellitus (T1DM) is an incurable and chronic autoimmune disease which is classically identified by reduced level of insulin due to autoimmune destruction of insulin-producing pancreatic β cell, leading to hyperglycemia (86). Although underlying mechanism of disease is unknown, disruption of immune tolerance in autoreactive B cells, CD4+ T cells and particularly CD8+ cytotoxic T lymphocytes (CTL) are believed to contribute to the pathogenesis of type 1 diabetes. As a result, diabetic patients need exogenous insulin regiment to control their blood glucose level and avoid from consequences caused by hyperglycemia like fatigue, cataract, and also diabetic coma. Repetitive injection of insulin point to the fact that insulin titer is not stable in affected subjects and patients require a durable cure for diabetes.
In order to develop novel therapies for T1DM, researchers have taken different approaches so far. Since first whole pancreas transplantation in 1966, long-term normoglycemia has been achieved with a graft survival rate of 50 to 70 % after 5 years (87, 88). But sequential complications including pancreatitis, thrombosis, and pseudocyst appeared in graft are major disadvantages of this surgery (89). Eventually, transplantation of endocrine part of pancreas (known as islet transplantation) is a well-developed procedure to minimize complications of insulin therapy and pancreas transplantation (90). last two methods prevent from the disease progression for a limited period of time, but can not correctify autoimmune response against the β cells. The cure must be in the region where autoimmune response is originated.
Various strategies have been applied to recover immune tolerance at critical points of regulatory pathways. Of the different approaches, anti-CD3 antibodies nonspecifically induce T cell anergy by co-stimulation blockade. Thereafter, beta cells are preserved in disease established non-obese diabetic (NOD) mice (91). Traditional αCD3 antibodies like OKT3 were provided from mouse and could not be administered continuously for chronic diseases. This drawback is overwhelmed by humanizing the antibody sequence (92). Moreover, mitogenic effect of CD3 specific antibodies on T cells, owning to their cross-linking ability to bind to the Fc receptors on natural killer cells and monocytes, brings about a notable cytokine release (93). This problem can be solved via engineering of C-terminal sequence of antibody to stop its binding possibility to Fcγ receptors.
MSC therapy has also shown antidiabetic effects to restore immune response balance in pancreas. Ezquer et al. (94) injected GFP expressing MSCs to diabetic mouse and reported normalized Th1/Th2 response in diabetic group compared to control. They also traced labeled MSCs after 7 and 65 days to be sure if MSCs differentiate into beta cells and lead to normoglycemia. Surprisingly, they found MSCs in heart and secondary lymphoid organs (like Payer’s patch, inguinal lymph node, and pancreatic lymph nodes) not in pancreas. It suggests that the antidiabetic effect of MSC is due to its endocrine activity not differentiation capacity. In 2016, Nakano et al. (95) have used MSC-derived exosomes to diminish cognitive impairments of diabetic rats by repairing damaged astrocytes and more importantly neurons. Furthermore, administration of MSC-derived exosomes simultaneously with islet transplantation, suppressed proliferation of PBMCs, improved regulatory T cell activities and increased graft survival in syngeneic mouse model of T1DM (96). Similar to previous studies, a group of scientists also evaluated immunomodulatory effects of MSC-derived exosomes compared with control and reported significant increase of regulatory T cell population among splenic MNCs, accompanied by upregulation of IL-4, IL-10, TGF-β plus downregulation of IL-17 and IFNγ cytokines after treatment. Accordingly treatment stabilized the blood glucose and vanished the signs of diabetes (97).
Go to :
Uveitis
Uveitis is an autoimmune disorder which causes severe visual disability (98). Current clinical treatment includes corticosteroids, other immunosuppressive drugs, and newly developed biologics. The long-term utilization of each is followed by serious systemic side effects and the local risk of cataracts and glaucoma (99, 100). Hence, most investigation intended to find localized and specific therapies. Although, it has been recently revealed that i.v. injection of MSCs improved experimental autoimmune uveitis (EAU) in animal model of uveitis (101-103), the underlying mechanisms of MSCs is not clear yet. In 2017, Bai et al. (104) showed that, human umbilical cord MSC-derived exosomes can reduce the severity of EAU. They demonstrated that MSC-derived exosomes inhibited the infiltration of inflammatory leukocytes to the eyes and these outcomes could, at least partly, be due to expression of CD73 on the MSCs. As expected, exosomes bearing CD73 interact with CD39 on activated immune cells and induce production of adenosine which result in proliferation inhibition. This study is the only study on the role of exosomes uveitis, indicating the importance of further investigation in this field.
Go to :
Conclusions
Until last decade, the contribution of MSC-derived exosomes in the control of inflammatory responses has been less clear. As a functional vesicle derived from MSCs, exosomes have shown same beneficial impacts of the cell of origin, to suppress several autoreactive cells in autoimmune diseases. Moreover, although mesenchymal-derived exosomes expose immunomodulatory functions by self, it seems that engineering these vesicles through adding anti-inflammatory molecules and certain receptors, they can specifically target tissue/organ of interest. This attitude might consider as a potentially novel candidate to treat other autoimmune disorders like spondylitis ankylosing and scleroderma as well.
With respect to remaining challenges, more investigations are required to elucidate unclear aspects of cell-free therapy using MSC-derived exosomes in ongoing studies.
Go to :
 XML Download
XML Download