

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Hepatic encephalopathy (HE) is a neuropsychiatric syndrome of both acute liver failure and chronic liver disease with symptoms ranging from subtle fluctuating cognitive impairment to coma [1]. It is characterized by personality or mood changes, abnormal movements, intellectual impairment, and a depressed level of consciousness [1]. Overt HE occurs in 30–40% of patients with cirrhosis, whereas minimal HE affects up to 80% of patients with cirrhosis [1,2]. Although the incidence of HE is high, there is still a paucity in the understanding of precise pathogenesis of HE, which calls for basic research into the pathogenic mechanism of HE.
For HE research, a wide range of animals have been used, including large animals (dogs, goats, pigs, and rabbits) and rodent (rats and mice) [3]. Hepatotoxin models have been widely used to study HE caused by acute liver failure [3–5]. Meanwhile, for the research of HE in chronic liver disease, bile duct ligation (BDL) has been used [3,6–11]. BDL in rodents provides a model of secondary biliary fibrosis, which is characterized by proliferation of bile ductules, cholestasis, portal inflammation, and fibrosis [12,13]. Rats are particularly adapted to BDL model because they do not have a gall bladder; however, BDL is also used in mice although marked dilation of the gall bladder occurs after BDL [12,14].
BDL can be performed by double ligation of the common bile duct with or without transection at the midpoint between the two ligatures [6–11,13,14]. However, in the case of dissection of the bile duct between the ligatures, there is the risk of bile leaks if one knot is not secure, which results in severe peritonitis and mortality [14]. In C57BL/6 mice, double ligation of the common bile duct without transection successfully induces inflammatory liver injury and fibrosis with a low mortality rate; liver enzymes such as alanine aminotransferase (ALT) and aspartate transaminase (AST) peak 10–14 days after BDL and periportal fibrosis and perisinusoidal fibrosis appear 14 days after BDL [14].
Recently, mice are rapidly overtaking rats as the major model of choice in biological research; mice accounted for about 20% of test animals in the 1970s and 1980s, and then grew to around 50% in recent years [15]. This shift may be related to the availability of techniques for genetic modification of mice [15]. Among mice, C57BL/6 mice are the most commonly used mouse in biomedical research [15]. A wide array of genetically engineered mutant mice on the C57BL/6 background have provided extensive insights into specific genes and pathways involved in different liver diseases; therefore, C57BL/6 mice remain the most widely used strain in liver research [15]. However, whether BDL in C57BL/6 mice is useful for HE research has not been established. Our aims in this study were to evaluate the suitability of a BDL model using C57BL/6 mouse for the study of HE and to determine which behavioral test is appropriate for the identification of HE in this model.
Go to : 
MATERIALS AND METHODS
Animals
All procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Yonsei University Health System (no. 2018-0068). Twelve to fourteen-week-old male C57BL/6 mice (Orient Bio, Korea) were used for this study. Mice were randomly assigned to sham group (n = 12) or BLD group (n = 14). For the 10% possibility of mortality, two more mice were assigned to BDL group. Animals were housed under a 12-h light/dark cycle in a temperature-controlled room with ad libitum access to food and water.
Surgery for bile duct ligation
All surgery for BDL was performed as described in a previous study with slight modifications [14]. Sham-operated mice were laparotomized without BDL.
1) Anesthesia was induced by inhalation of 4 vol% isoflurane, and maintained with 1.5–2.0 vol% isoflurane in 100% oxygen at a flow rate of 1 L/min.
2) The mouse was placed on a 37°C heated hot plate. The abdominal fur of the mouse was removed with an electric fur shaver and the eyes were protected by using an eye ointment.
3) The abdominal skin was sterilized with betadine followed by 70% ethanol (2–3 times) using cotton swabs.
4) The abdomen was opened by a midline laparotomy approximately 2 cm in length with a surgical scissor, and the connective tissue on top of the peritoneum was dissected using a pair of scissors as a spreader.
5) The peritoneum was cut along the linea alba to open the peritoneal cavity.
6) The cavity was enlarged by inserting a holding suture in the sternum, raising the filament of the suture, and fixing it on top of the circuit. The peritoneal cavity was enlarged with a retractor.
7) The liver was lifted with a moistened (0.9% NaCl solution) cotton swab so that the ventral side of it stuck to the diaphragm and the hilum was clearly visible. The bile duct was exposed by caudal movement of the gut.
8) The bile duct was separated from the portal vein and hepatic artery using micro-serrations forceps (Fig. 1A).
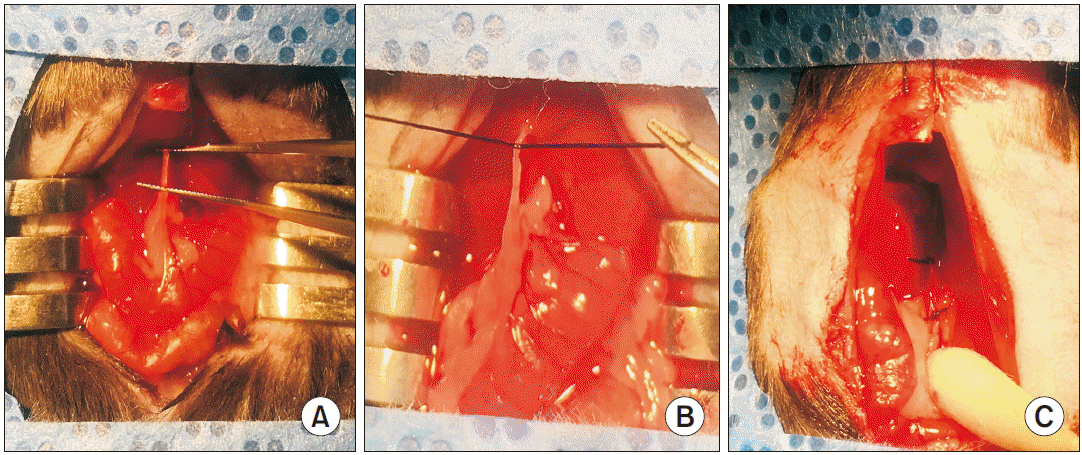
9) A 5–0 silk suture was placed around the bile duct and secured with two surgical knots (Fig. 1B). A second cranial ligation was added in the same manner but did not dissect the bile duct in-between (Fig. 1C).
10) The ends of the sutures were cut, the sternum was lowered, and the retractor was removed.
11) A 0.9% NaCl solution (0.3–0.5 ml) was applied to the peritoneal cavity and the abdominal organs were replaced in their physiological positions.
12) The peritoneum and skin were closed with simple continuous suture with 5–0 silk, and the operation area was sterilized with betadine followed by 70% ethanol (2–3 times) using cotton swabs.
13) Intraperitoneal injection of tramadol 0.25 mg/kg was performed immediately after surgery.
Behavioral tests
All behavioral tests were performed as described in a previous study with the modifications described below [16]. Symptoms of minimal HE include anxiety and impaired memory [1,2]. Therefore, open filed test and elevated plus maze test were performed to test anxiety, and novel object recognition test and passive avoidance test were performed to test memory. For environmental acclimation, mice were placed into a behavior room for 1 h prior to the test. After each trial, the apparatus was cleaned with distilled water and then cleaned with 70% ethanol and the inter-trial interval was 3 min. Behavioral data were automatically analyzed using SMART Video Tracking system, which tracks nose, tail, and center of the mouse’s body (Panlab Harvard Apparatus, Spain) with SMART v2.5.21 software.
Open field test (13 days after surgery)
After acclimation of the behavior room in the home cage, the mouse was gently placed in the center of an open field chamber (40 × 40 × 40 cm, JEUNGDO Bio & Plant Co., Ltd., Korea) and allowed to move freely for 10 min. The total distance moved and time spent in the center were recorded and analyzed to evaluate general locomotor activity and level of anxiety, respectively.
Elevated plus maze (14 days after surgery)
The maze consisted of two open arms (31 × 6 × 1 cm) and two enclosed arms (31 × 6 × 15 cm) with a central open square area (5 × 5 × 1 cm), and was elevated by 50 cm (JEUNGDO Bio & Plant Co., Ltd.). Mice were individually placed in the center facing an open arm and allowed to explore for 5 min. Duration of open arm and closed arm entries were recorded. Entry was defined as movement of all paws into an arm. The percentage of time spent in open arms was measured.
Novel object recognition test (14 days after surgery)
To evaluate recognition memory, the novel object recognition test was performed in the same chamber of open filed test. After 24 h of open filed test, the individual mouse, which was habituated to the chamber during open field test, was presented with two equal sample objects for familiarization and allowed to explore freely for 10 min. After 1 h, one of the familiar objects was replaced with a novel object and free exploration was allowed for 10 min [17]. Recognition memory was expressed as the percentage of novel object exploration time, which was calculated as (novel object exploration time/[familiar object exploration time + novel object exploration time]) × 100%.
Passive avoidance test (13 and 14 days after surgery)
To evaluate learning/memory as a result of a fear response, the passive avoidance test was performed in a 41 × 21 × 30 cm plastic chamber consisting of light and dark compartments separated by a guillotine door (JEUNGDO Bio & Plant Co., Ltd.). The stainless steel grid on the floor of the dark compartment was capable of administering an electric shock. In the training trial (13 days after surgery), each mouse was placed in the light compartment and allowed to explore the chamber for 10 s. The door between the two compartments was opened 10 s after the mouse was placed on the light compartment and the mouse was allowed to explore and move freely into dark compartment. When the mouse entered the dark compartment completely, the door was closed and an electric foot shock (0.5 mA) was applied for 3 s through the steel grid on the floor. Twenty-four hours after the training trial (14 days after surgery), retention trial was performed in the same chamber. Identical to the training trial, the mouse was placed in the light compartment and then 10 s later, the door was opened. The time to enter the dark compartment (transfer latency) was recorded. If the mouse did not enter the dark side within 300 s, transfer latency was recorded as 300 s.
Laboratory tests (14 days after surgery)
After behavioral tests, whole blood obtained by cardiac puncture and ammonia levels were measured using the PocketChem BA PA-4140 (Arkay Inc., Japan) according to the manufacturer’s instructions. Analysis of ALT level was performed in serum by the Korea Animal Medical Science Institute (KAMSI, Korea) using Hitachi 7180 chemistry analyzer (Hitachi High-Technologies Co., Japan).
Liver histology (14 days after surgery)
Livers were fixed in 4% paraformaldehyde overnight at 4°C, embedded in paraffin, and then sectioned to obtain 4-μm slices. Hematoxylin and eosin (H&E) and Masson’s trichrome staining were performed and stained slides were examined using an Olympus BX53 microscope (Olympus, Japan).
Statistical analysis
Data are expressed as mean ± standard deviation or median (1Q, 3Q) of independent experiments. Normality of distribution was assessed with D’Agostino-Pearson test. Independent t test was used for parametric data (ALT, ammonia, total distance moved and time spent in the center [open field test], time spent in open arms [elevated plus maze test], and percentage of novel object exploration time [novel object recognition test]), and Mann–Whitney U test was used for nonparametric data (transfer latency [passive avoidance test]). Body weight was analyzed by two-way ANOVA with repeated measures in one factor. P < 0.05 was considered statistically significant, and all analyses were performed with GraphPad Prism Software v7.0 (GraphPad Software, USA).
Go to :
RESULTS
Histological changes of the liver in the BDL model
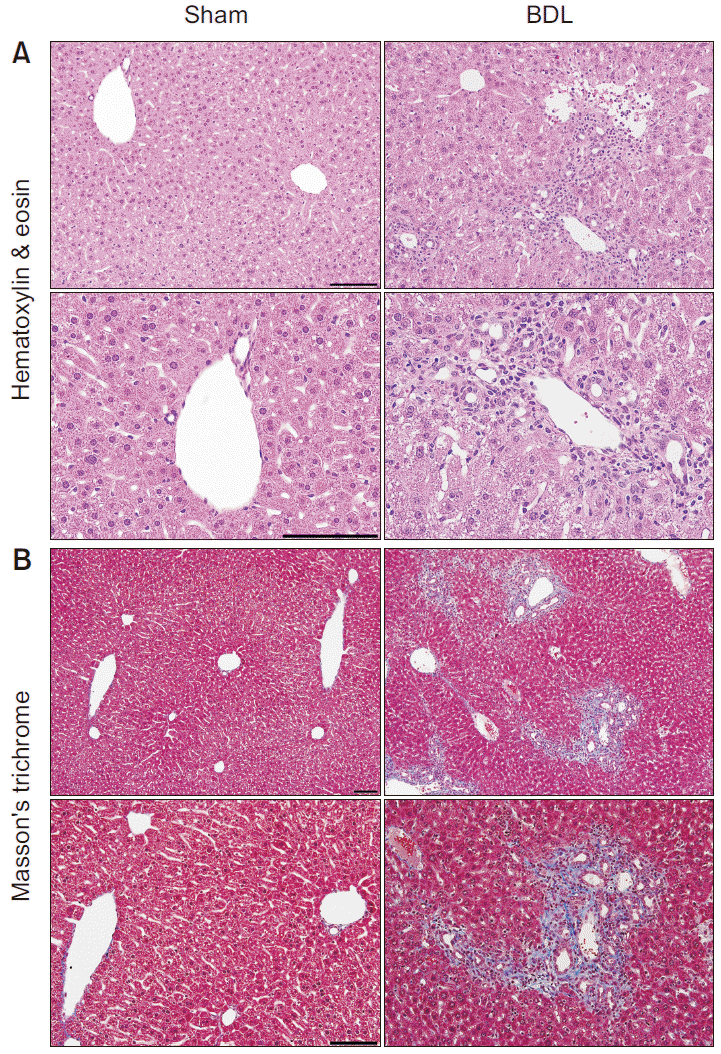
To assess histological changes and liver fibrosis, H&E and Masson’s trichrome staining were performed 14 days after surgery (Fig. 2). In H&E staining, mononuclear cell infiltration around the portal vein and massive cytoplasmic degeneration with vacuolated cytoplasm was observed in BDL group. Moreover, pyknotic nuclei and hepatocyte necrosis were evident in widened areas, especially surrounding the portal vein in BDL group (Fig. 2A). In Masson’s trichrome staining, marked fibrotic areas (stained blue) around the portal vein were revealed in BDL group (Fig. 2B).
Alteration of physiological and biochemical parameters in the BDL model
Although body weights were similar between the groups before surgery (28.9 ± 1.4 vs. 29.9 ± 1.2 g in sham group and BDL group, respectively, P = 0.416), the body weight was significantly less in BDL group 14 days after surgery compared to the sham group (23.1 ± 1.9 vs. 29.3 ± 1.6 g, respectively, P < 0.001; Fig. 3A). The level of ALT was more than 25-fold higher in BDL group than in sham group (1,013 ± 599 vs. 37 ± 14 U/L, respectively, P = 0.007; Fig. 3B). In addition, BDL group showed a significant increase in blood ammonia levels compared to sham group (297 ± 82 vs. 101 ± 48 µg/dl, respectively, P < 0.001; Fig. 3C).
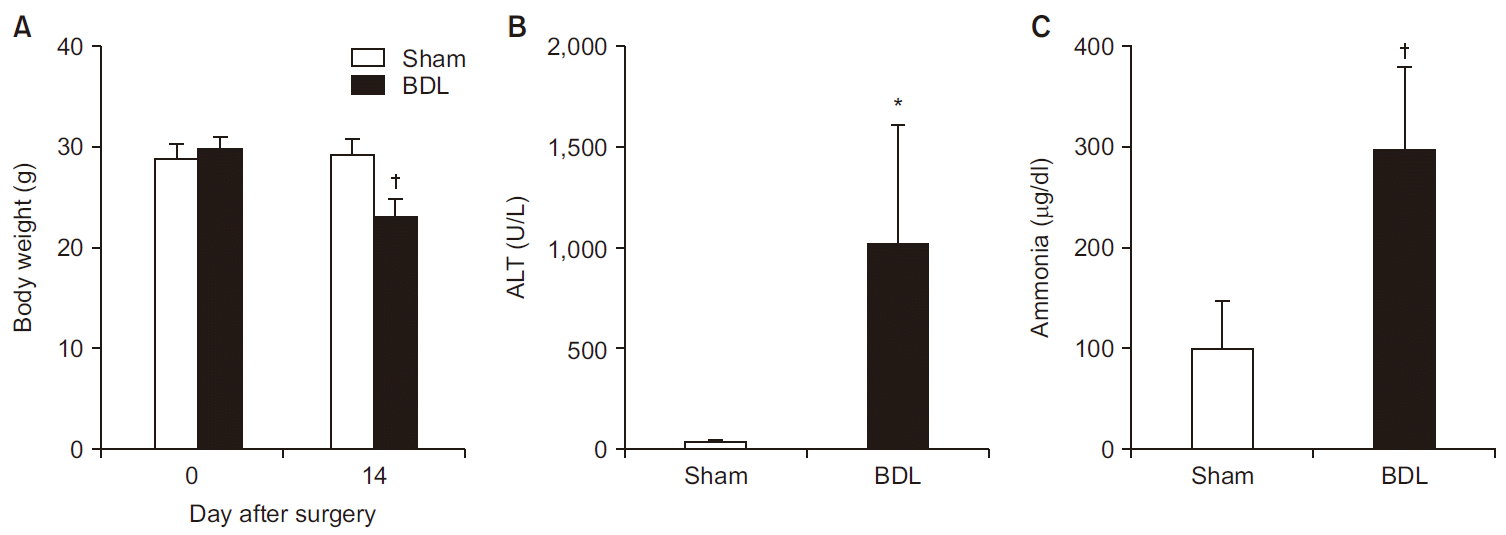 | Fig. 3(A) Changes in body weight after 14 days of surgery (n = 8 per group). Level of (B) serum ALT (n = 5 per group) and (C) blood ammonia (n = 7 per group) after 14 days of surgery. Values are presented as mean ± SD. ALT: alanine aminotransferase, BDL: bile duct ligation. *P < 0.01, †P < 0.001 vs. sham group.
|
No alteration of general activity and anxiety in BDL model
General locomotor activity (total distance moved) measured by open field test were not different between the groups (2,732 ± 525 vs. 2,343 ± 389 cm in sham group and BDL group, respectively, P = 0.114; Fig. 4A). The level of anxiety assessed by open field test (time in center) and elevated plus maze (time in open arms) were similar between the groups. In open field test, 10.9% of time was spent in the center in sham group and 8.5% of time in BDL group (P = 0.503; Fig. 4A). In elevated plus maze test, 19.9% of time was spent in open arms in sham group and 20.1% of time in BDL group (P = 0.977; Fig. 4B).
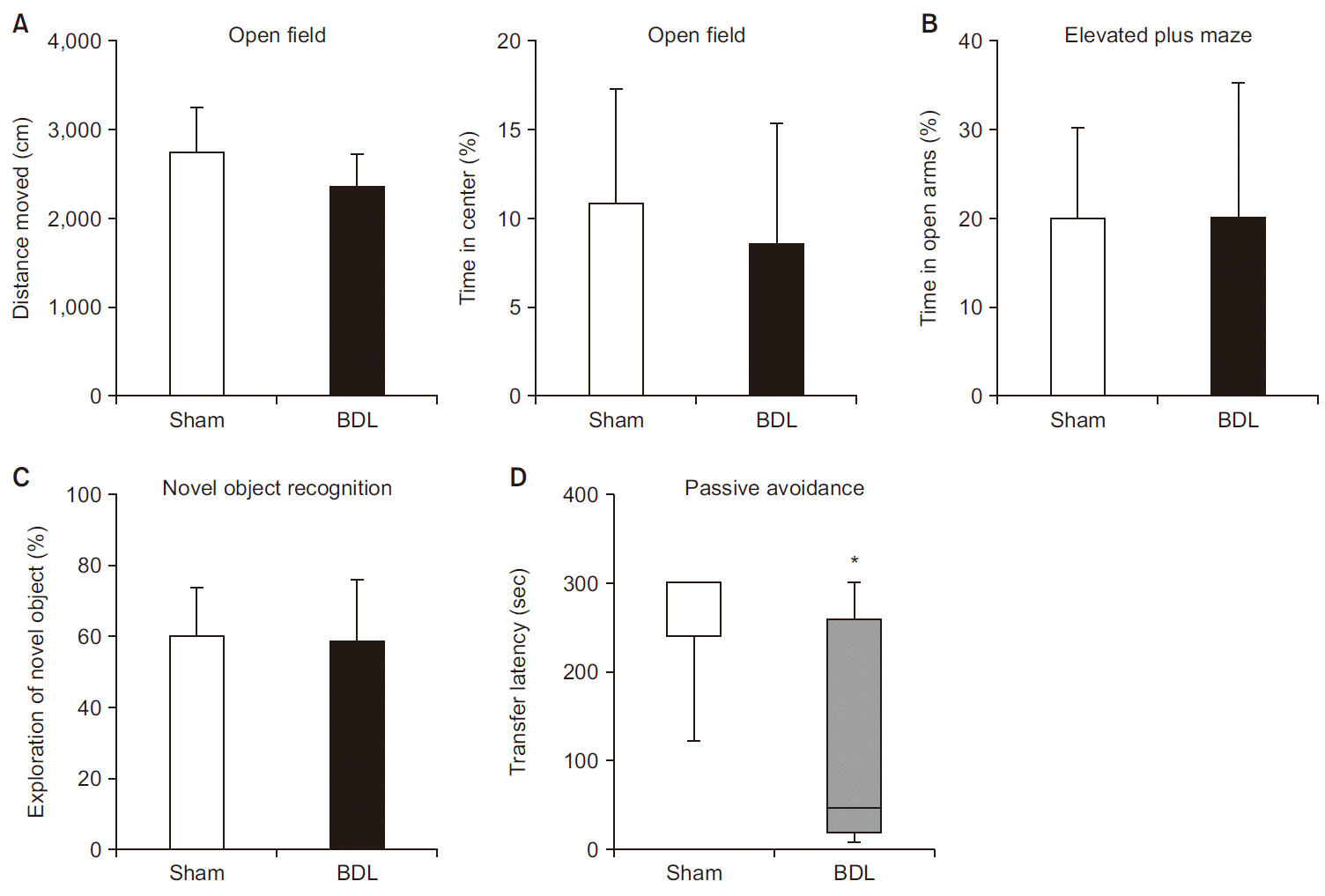 | Fig. 4(A) Total distance moved (left) and percentage of time spent in the center zone (right) in the open field test (n = 8 per group). (B) Percentage of time spent in the open arms in the elevated plus maze test (n = 8 per group). (C) Percentage of novel object exploration time in the novel object recognition test (n = 8 per group). (D) Transfer latency to enter the dark compartment in the passive avoidance test (n = 8 per group). Transfer latency is expressed using a box plot with median (solid line), interquartile range (box), plus maximum and minimum values (whiskers). Other values are presented as mean ± SD. BDL: bile duct ligation. *P < 0.05 vs. sham group.
|
Impairment of learning/memory in the BDL model
Learning/memory was assessed by novel object recognition test and passive avoidance test. In the novel object recognition test, the time spent exploring novel objects was similar for the two groups (59.9 ± 14.3 vs. 58.5 ± 17.8% in sham group and BDL group, respectively, P = 0.571; Fig. 4C). However, in the passive avoidance test, transfer latency to enter the dark compartment was shorter in BDL group than in sham group (48.5 [20.8, 259] vs. 300 [241.5, 300] s, respectively, median [1Q, 3Q], P = 0.018), which indicates learning/memory impairment in BDL mice (Fig. 4D).
Go to :
DISCUSSION
This study demonstrated that liver fibrosis was induced and learning/memory impairment appeared 14 days after BDL in C57BL/6 mice. Therefore, this 14 day-BDL model in C57BL/6 mouse can be used to study HE and the passive avoidance test can be used to identify HE in this model.
HE is subdivided into type A, which is caused by acute liver failure, type B, which is predominantly due to portosystemic bypass or shunting without associated intrinsic liver disease, and type C, which is caused by cirrhosis [1]. To create type A HE rodent models, various types of hepatotoxic substances has been used: thioacetamide, acetaminophen, and carbon tetrachloride in rats [3,18–20], and azoxymethane in mice [3–5]. Among 6,307 patients who visited the liver clinic of a single community hospital in Korea, 3,957 (62.7%) patients were diagnosed with chronic hepatitis, whereas 528 (8.4%) were diagnosed with acute hepatitis [21]. Therefore, a type C HE model may be more appropriate than a type A HE model in Korea in that type A HE has different manifestations from type C HE such as a prominent increase in intracranial pressure with cerebral edema [1].
Since BDL model has features of chronic liver disease and is technically relatively simple and quick to perform [13,14], it can be considered good experimental models for type C HE [3]. In fact, several studies have used BDL model in mice for HE research [6–11]. BDL in mice induces inflammatory liver injury and fibrosis through obstructive cholestasis [13,14]. After BDL, ductular reaction is induced by the proliferation of biliary epithelial cells, while hepatocytes in areas of proliferated bile ducts undergo necrosis and apoptosis [13,14]. As the degree of fibrosis after BDL can be dependent on the genetic background, sex, and age of the mice [12,13], confirmation of liver fibrosis is crucial in each mouse strain that is planned for use. In the present study, double ligation of the common bile duct without transection was applied in male C57BL/6 mice, and inflammatory liver injury and fibrosis were observed 14 days after BDL with mortality rate of 7% (1 of 14). Therefore, a 14-day BDL model can be applied in male C57BL/6 mouse for research on chronic liver disease.
To evaluate brain function alterations in a disease model, determination of appropriate time points for behavioral tests is important. In previous studies using BDL model, behavioral tests were performed at a different time point [6–11]. In studies using female Sabra mice (8 weeks of age, 25–30 g), eight-arm maze test (learning/memory) and open filed test (motor function) were performed 3 weeks after BDL [8,9]. Meanwhile, in studies using male NMRI mice (25–35 g), passive avoidance test (learning/memory) and novel object recognition test (learning/memory) were performed 24 days after BDL and 4 weeks after BDL, respectively [10,11]. Therefore, we had to determine the time point for behavioral tests, as no previous study had evaluated learning/memory function in C57BL/6 mice after BDL. In previous studies using male C57BL/6 mice (6–8 weeks of age), recruitment of activated monocytes into the brain were observed 10 days after BDL [6,7], which implies that inflammatory responses were already triggered in the brain 10 days after BDL. In the present study, impairment of learning/memory was identified by a passive avoidance test 14 days after BDL when liver fibrosis and elevation of ALT were also observed. A previous study reported that ALT and AST peaked after 10–14 days, total bilirubin reached a plateau after 7 days, and periportal fibrosis and perisinusoidal fibrosis manifested after 14 days of BDL [14], thus 14 days of BDL may be appropriate to investigate secondary organ dysfunction in liver disease.
Mice aged 12–14 weeks were used in the present study, in contrast to previous studies that used 6–8-week-old C57BL/6 mice [6,7]. Many investigators choose 8–12-week-old mice usually because of cost; however, many developmental processes occur during this timeframe that can have a large impact on experimental variables [22]. Three- to six-month-old C57BL/6 mice (equivalent to 20–30 human years) are considered mature adult mice [22]. Therefore, we argue that 12- to 14-week-old male C57BL/6 mice are good candidates for research on HE.
In the present study, general locomotor activity was assessed by an open field test (total distance moved) and anxiety was assessed by an open field test (time in center) and elevated plus maze test (time in open arms) [23,24]. General locomotor activity was not impaired and anxiety was not increased 14 days after BDL in C57BL/6 mice. To investigate hippocampus-dependent learning/memory, we used a novel object recognition test and passive avoidance test [23–25]. No difference between BDL and sham-operated mice was observed in the novel object recognition test; however, BDL mice showed significant learning/memory impairment in a passive avoidance test compared to sham-operated mice. The reason for memory impairment in the passive avoidance test but not novel object recognition test may be due to the different stress intensities of the two tests. Novel object recognition test is relatively less stressful, however the passive avoidance test is fairly stressful because the mouse receives an aversive stimulus (electric foot shock) when entering a comfortable environment (dark compartment) [23–25]. In fact, the passive avoidance test has been used in various models of brain disease to evaluate learning/memory impairment because strong memory retention can be gained after a single training trial in most mouse strains [24]. Therefore, the passive avoidance test may be the best option to detect minimal HE in C57BL/6 mice.
Although the correlation between blood ammonia level and severity of HE is not always consistent, ammonia is a crucial factor in the pathogenesis of HE [26]. This is supported by the fact that neurological/psychiatric symptoms and electroencephalographic abnormalities improve when blood ammonia levels are lowered in HE patients [27,28]. Moreover, the severity of HE has been found to correlate well with blood and brain ammonia levels in animal models of HE [29,30], and feeding rodents a high-ammonia diet is one of the methods used to induce HE in rodents [3]. Therefore, a high level of blood ammonia, along with an abnormal behavioral test, can serve as supportive evidence of HE in mice. In our study, blood ammonia levels were approximately double in BDL mice than sham-operated mice, which supports our argument that a 14-day BDL model in C57BL/6 mouse is a clinically relevant model for HE. However, further studies investigating the effects of hyperammonemia on the alteration of signal transduction pathway, gene expression, and astrocyte function are needed to confirm the development of HE.
In conclusion, a 14-day BDL model using 12- to 14-week-old male C57BL/6 mice is an appropriate HE model of chronic liver disease as evidenced by liver fibrosis with impaired liver function, hyperammonemia, and learning/memory impairment. Passive avoidance can be considered as the primary behavioral test of HE in this model.
Go to :
 XML Download
XML Download