

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Postoperative pain is directly associated with surgical incision and tissue damage inflicted during surgery. Inflammatory pain, caused by tissue injury, develops in patients with arthritis. Insults to tissue directly activate peripheral nociceptors to cause pain. Such pain is well-controlled by opioids or nonsteroidal anti-inflammatory drugs in most cases; but both drugs cause serious side-effects including respiratory depression, gastrointestinal discomfort, renal dysfunction, and hematologic abnormality. Furthermore, and unfortunately, tissue injury may unexpectedly enhance the activation of peripheral nociceptors, causing an activity-dependent increase in central neuronal excitability, in turn inducing both peripheral and central sensitization [1,2]. Such changes contribute to post-injury hypersensitivity, which manifests as an increase in responsiveness to painful stimuli (hyperalgesia), and a decrease in the pain threshold rendering (usually) non-painful stimuli painful (allodynia). Such events are not easy to manage using common analgesics, and compromise quality-of-life. Therefore, active pain therapy should commence early to avoid such sensitization. However, when pain develops, endogenous protective systems become activated to counteract the pain. Both γ-aminobutyric acid (GABA) and opioid receptors are well-known endogenous systems modulating nociception in the spinal cord [3].
Recently, much effort has been devoted toward development of materials meeting unmet therapeutic needs. In this context, traditional medicines have attracted a great deal of attention.
Curcumin is the principal biologically active component of turmeric, which is widely used as a food-coloring agent. Interestingly, curcumin has long been employed as an herbal medicine in Asian countries, including Korea. It is known that curcumin exhibits a variety of medical effects including anti-inflammatory action [4]. Recently, it has been shown that curcumin attenuates nociceptive behavior at the spinal level [5,6]. However, the spinal mechanisms of action of curcumin have not been fully elucidated.
Thus, the purposes of the present study were to explore spinal GABA and opioid receptor expression patterns in rats in which postoperative and inflammatory pain had been induced; to examine the effects of intrathecal curcumin on both types of pain; and, further, to assess the effects of curcumin on GABA and opioid receptor levels in the spine.
Go to : 
MATERIALS AND METHODS
Animal preparation
All experimental protocols were reviewed and approved by the Institutional Animal Care and Use Committee of Chonnam National University (no. CNU IACUC-H-2010-8). Adult male Sprague-Dawley rats (250–300 g) were used in the present study. Animals were housed four to a cage and kept in a vivarium maintained at 22°C with a 12 hours : 12 hours alternating light/dark cycle. All rats received food and water ad libitum. All test drugs were intrathecally administered. Thus, rats were implanted with intrathecal catheters under sevoflurane anesthesia, as described previously [7]. An 8.5-cm-long polyethylene-10 catheter was advanced caudally, through an incision in the cisternal membrane, to the thoracolumbar level of the spinal cord. The exterior portion of the catheter was secured at the skull via subcutaneous tunneling, and was closed with a 30-gauge wire. The skin was sutured with 3-0 silk. After catheter implantation, rats were housed in individual cages. Only animals with no evidence of neurological deficits developing after catheter implantation were included in the present study; such rats were housed individually. The behavioral study was performed 5 days after intrathecal catheterization.
Drugs
The following drugs were used in the present study: curcumin [1,7-Bis (4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione, Tocris Cookson, UK]; bicuculline [R-(R*,S*)]-6-(5,6,7,8-Tetrahydro-6-methyl-1,3-dioxolo[4,5-g]isoquinolin-5-yl) furo[3,4-e]-1,3-benzodioxol-8 (6H)-one, Tocris Cookson, UK]; saclofen [RS)-3-Amino-2-(4-chlorophenyl) propylsulfonic acid; Tocris Cookson, UK]; CTOP [D-Phe-Cys-Tyr-D-Trp-Orn-Yhr-NH2, Tocris Cookson, UK]; naltrindole [17-(Cyclopropylmethyl)-6,7-dehydro-4,5-epoxy-3,14-dihydroxy-6,7-2’,3’-indolomorphian hydrochloride, Tocris Cookson, UK]; and GNTI [5’-Guanidinyl-17-(cyclopropylmethyl)-6,7-dehydro-4,5-epoxy-3,14-dihydroxy-6,7-2’,3’-indolomorphian dihydrochloride, Tocris Cookson, UK]. Curcumin was dissolved in 10% dimethylsulfoxide (DMSO), whereas all other drugs were dissolved in distilled water. Intrathecal administration of all agents was performed using a hand-driven, gear-operated syringe pump. All drugs were delivered in volumes of 10 μl followed by an additional 10 μl of normal saline to flush the catheter [8].
Nociceptive test
Postoperative pain model
Postoperative pain was induced during sevoflurane anesthesia, as previously described [9]. The plantar surface of the left hindpaw was rendered sterile. A 1-cm longitudinal incision was made through the skin and fascia of the plantar aspect of the foot, commencing 0.5 cm from the proximal edge of the heel and extending toward the toes. The plantaris muscle was elevated and incised longitudinally, leaving the muscle origin and insertional points intact. After hemostasis was achieved by application of gentle pressure, two 4-0 silk sutures were placed in a mattress fashion along the wound. Antibiotic ointment was next applied to the incision site and rats were allowed to recover in their cages. Wounds were checked prior to behavioral testing.
Inflammatory pain model
Inflammatory pain was induced by intraplantar injection of carrageenan [10]. Animals were injected subcutaneously with 100 μl amounts of 2% (w/v) carrageenan (degraded λ-carrageenan, Sigma Aldrich, USA) into the center of the plantar surface of the left hind paw, using a 30-G needle. Carrageenan was dissolved in saline to form the 2% (w/v) solution and stored at room temperature for 24 hours before use. Carrageenan was injected under sevoflurane anesthesia.
Behavioral pain testing
The extent of pain was evaluated by measurement of the paw withdrawal threshold to mechanical stimulation delivered using von Frey filaments. Withdrawal thresholds were measured prior to manipulation, on rats in cages with wire mesh floors, and these measurements were the baseline values. Next, operations were performed or carrageenan injected, and withdrawal thresholds measured for 4 hours. Paw withdrawal thresholds in response to mechanical stimulation were measured using the up-and-down method [11]. Employing calibrated von Frey filaments (Stoelting, USA) inserted from underneath each cage, through openings in the wire mesh floor, to touch the hindpaw. A series of eight von Frey filaments (0.4, 0.7, 1.2, 2.0, 3.6, 5.5, 8.5, and 15 g) were applied vertically to the plantar surface of the hindpaw for 5 seconds, with sufficient force to bend the filament. Brisk withdrawal or paw flinching was considered to be a positive response. In the absence of a response to the 15 g filament, a 15 g cut-off value was assigned. All tests were performed in duplicate, with approximately 3 minutes between each test, and average values were used. Only rats exhibiting marked allodynia (a withdrawal threshold < 5 g) after paw incision were studied. At the end of the experiment, all rats were euthanized using volatile anesthetics.
Experimental protocol
On the days of behavioral experiments, rats were placed in wire cages for 30 minutes to allow acclimation, and were next allocated to receive an experimental drug. The same volume of vehicle (distilled water or DMSO) was given to control animals. All rats were used only once (i.e., no rat received two test agents). All experimental tests were conducted by investigators blinded to treatment.
Effects of curcumin on postoperative and inflammatory pain
Withdrawal thresholds were determined before and 2 hours after incision, and before and 4 hours after carrageenan injection, and after intrathecal administration of curcumin (10, 30, or 100 μg in rats subjected to incisional pain; and 30, 100, or 300 μg in animals subjected to inflammatory pain). Withdrawal thresholds measured 2 hours after incision or 4 hours after carrageenan injection were considered to reflect the post-incision/post-injection control thresholds. Test drugs were intrathecally delivered immediately after measurement of these thresholds. Each ED50 value (the dose producing a 50% reduction in the control response) of curcumin was calculated separately.
GABA and opioid receptor expression upon induction of postoperative and inflammatory pain
The levels of mRNAs encoding GABA (A and B) and opioid receptors (μ, δ, and κ) were measured in the left dorsal quadrant of the spinal cord in naïve, incised carrageenan-injected and curcumin-treated rats (n = 16) via reverse transcriptase polymerase chain reaction (RT-PCR). Curcumin was administered intrathecally 2 or 4 hours after incision or carrageenan injection, respectively. At 2 hours after incision, 4 hours after carrageenan injection, and 60 minutes after curcumin administration, rats were euthanized by decapitation and the spinal cords quickly removed and stored at −80°C. The regions of spinal cords from L4 to L6 were dissected out and total RNA extracted using an RNAiso Plus kit (Takara Bio, Japan) according to the manufacturer’s instructions; precipitated with isopropanol; washed with 75% (v/v) EtOH; air-dried; and dissolved in 20-μl amounts of diethyl pyrocarbonate-treated water. RNA yields were determined by measuring absorbances at 260 and 280 nm. RT-PCR was performed with the aid of a PrimeScript™ RT-PCR kit (Takara Bio, Japan) according to the manufacturer’s protocol. Total RNA (1 μg amounts) was reverse-transcribed for 60 minutes at 42°C. PCR amplification was performed with the aid of an HS Prime Taq Premix kit (Genet Bio, Korea), and featured 30 cycles of denaturation (95°C for 30 seconds), annealing (57°C for 30 seconds), and extension (72°C for 30 seconds). Primer sequences amplifying the GABA receptors (A and B), the opioid receptors (μ, δ, and κ), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), were as follows: GABA-A, forward 5’-AAG AAG CTA TGG ACA GCC CT-3’ and reverse 5’-CCT CTG TGA TAC GCA GGA GTT-3’ (380 bp); GABA-B, forward 5’-ATC AGG TAC CGT GGC TTG AC-3’ and reverse 5’-TCG TAG TCC GGC AGG ATG TC-3’ (509 bp); μ opioid, forward 5’-CGA CAG CCT GTG CCC TCA GAC-3’ and reverse 5’-TGG ATT CAG GCA GCT GTT CG-3’ (841 bp); δ opioid, forward 5’-GGA AAC GTG GCC GTT CGG AG-3’ and reverse 5’-CCA GCA CAC CAC GAA GGC TC-3’ (490 bp); κ opioid, forward 5’-GCG ATA GTC CTT GGA GGC AC-3’ and reverse 5’-AGT CCC TAA AAC ACC GCT TGA A-3’ (454 bp); and GAPDH, forward 5’-GAC TCT ACC CAC GGC AAG TT-3’ and reverse 5’-CAT CCA CAG TCT TCT GAG TGG CA-3’ (421 bp). PCR products were separated on 1.5% (w/v) agarose gels and visualized using the SYBR Safe DNA gel stain. Band intensities were measured densitometrically, and the relative values of GABA receptor (A and B) and opioid receptor (μ, δ, and κ) intensities to that of the GAPDH band were calculated for each sample.
Roles played by GABA or opioid receptor subtypes in the effects of curcumin on postoperative and inflammatory pain
To explore the involvement of GABA or opioid receptor subtypes in mediating the effects of curcumin, rats were pretreated with GABA or opioid receptor antagonists. In the PCR study, the levels of mRNAs encoding GABA receptors increased when incisional pain but not inflammatory pain was inflicted, whereas the opposite effects were noted when opioid receptor mRNA expression levels were calculated. Thus, the involvement of GABA receptors in mediating the effects of curcumin was investigated in the postoperative pain model only, whereas that of the opioid receptors was explored in the inflammatory pain model only. All antagonists were given intrathecally 10 minutes before administration of intrathecal curcumin (100 or 300 μg) and behavioral tests were performed after curcumin delivery. The maximal doses of GABA or opioid receptor antagonists that did not affect control responses, or cause behavioral abnormalities, were determined in pilot experiments or calculated by reference to earlier data [12]. The GABA receptor antagonists used were as follows: a GABA-A receptor antagonist (bicuculline, 0.3 μg); and a GABA-B receptor antagonist (saclofen, 100 μg). The opioid receptor antagonists used were as follows: a μ opioid receptor antagonist (CTOP, 15 μg, n = 11); a δ opioid receptor antagonist (naltrindole, 10 μg, n = 11); and a κ opioid receptor antagonist (GNTI, 50 μg, n = 10). The antagonists were given 2 or 4 hours after incision and carrageenan injection, respectively (as described in Results).
General behavior
To examine behavioral changes induced by curcumin, high drug doses (300 μg) were given intrathecally to five additional rats. Motor function was assessed by evaluation of the placing-stepping and righting reflexes. The first reflex was evoked by drawing the dorsum of each hind paw across the edge of a table. Healthy rats generally try to place the paw ahead, into a position facilitating walking. The second reflex was evaluated by placing each rat horizontally with its back on the table. Healthy rats exhibit immediate and co-ordinated bodily twisting to assume the upright position. Pinna and corneal reflexes were evaluated using a paper string [13]. Each reflex was evoked by stimulation of the ear canal or the cornea with a string. Healthy rats spontaneously shake their heads or blink, respectively. Behavioral normality was scored as present or absent.
Statistical analysis
Data are expressed as means ± standard errors of the means. Time response data are presented as withdrawal thresholds. Dose-response data are presented as percentages of the maximum possible effect (%MPE). To calculate ED50 values of curcumin given to alleviate incisional and inflammatory pain, the withdrawal threshold data derived via von Frey filament testing were converted to %MPEs, using the formula: %MPE = ([post-drug threshold − baseline post-incision threshold] / [pre-incision threshold − baseline post-incision threshold]) × 100 for postoperative pain; and %MPE = ([post-drug threshold − baseline post-injection threshold] / [pre-injection threshold − baseline post-injection threshold]) × 100 for inflammatory pain. Dose-response data were analyzed by one-way analysis of variance with Scheffe post hoc testing. ED50 values with 95% confidence intervals (CIs) were calculated via linear regression. The pharmacological actions of curcumin, and RT-PCR data, were compared using the t-test. P values < 0.05 were considered to reflect statistical significance.
Go to :
RESULTS
Incisional surgery and carrageenan injection decreased withdrawal thresholds, and such effects persisted for approximately 4 days in the ipsilateral paw, whereas the two nociceptive stimuli per se did not affect withdrawal thresholds of the contralateral normal hindpaw (Fig. 1). The decreases in withdrawal thresholds were greatest 2 or 4 hours (P < 0.001 vs. contralateral) after incision and carrageenan injection, respectively (Fig. 1).
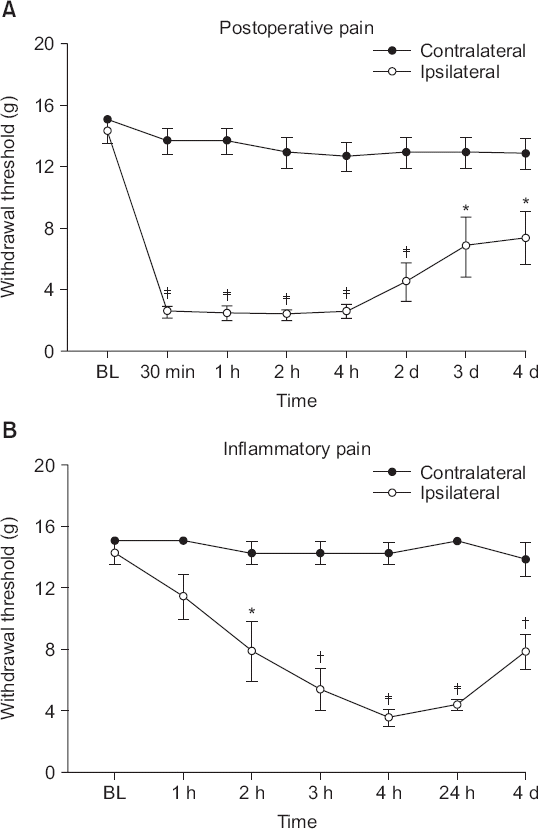 | Fig. 1Time course of hindpaw withdrawal responses to von Frey filament pressure after (A) incisional surgery or (B) carrageenan injection. Data are presented as withdrawal thresholds. Each line represents the mean ± SEM of data from five or six rats. BL: baseline withdrawal threshold measured prior to paw incision or carrageenan injection. Significant differences between data from the ipsilateral and contralateral sites are indicated, *P < 0.05, †P < 0.01, ‡P < 0.001.
|
RT-PCR revealed that mRNAs encoding opioid receptors (μ, δ, and κ) and GABA receptors (A and B) were present in the naïve rat lumbar spinal cord (Fig. 2). After incisional surgery, the levels of mRNAs encoding both GABA-A (P < 0.05 vs. naïve) and GABA-B (P < 0.001 vs. naïve) receptors decreased, but the levels encoding opioid receptors remained unchanged, compared to naïve rats (Fig. 2A). After carrageenan injection, the levels of mRNAs encoding all three μ (P < 0.05 vs. naïve), δ (P < 0.01 vs. naïve), and κ (P < 0.05 vs. naïve) subtypes of spinal opioid receptors increased, whereas the levels encoding the GABA receptors levels remained unchanged, compared to naïve rats (P > 0.05 vs. naïve, Fig. 2B).
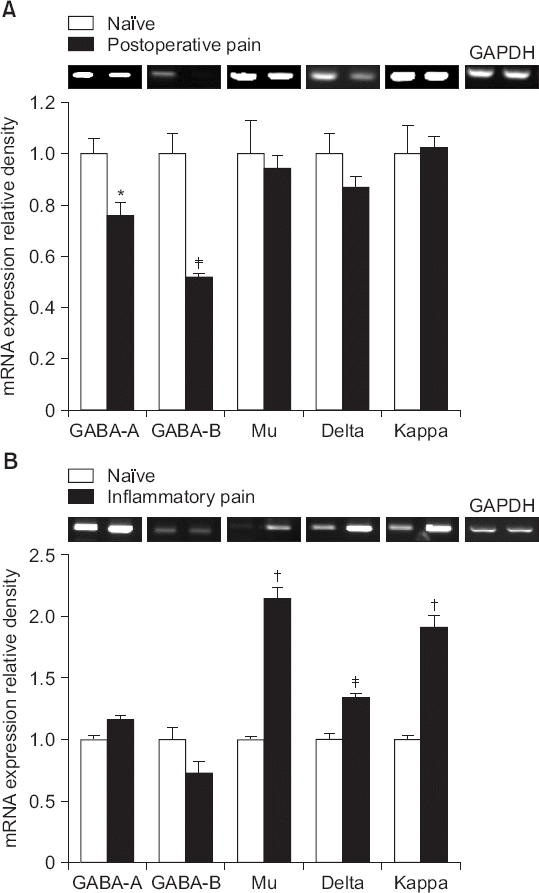 | Fig. 2Expression levels of mRNAs encoding γ-aminobutyric acid (GABA) and opioid receptors in the left dorsal horn of the spinal cord in naïve, incised, and carrageenan-injected rats. Data are presented as relative values of GABA and opioid receptors to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Each bar represents the mean ± SEM of data from four rats. Reverse transcriptase polymerase chain reaction detected mRNA bands corresponding to the GABA and opioid receptors in naïve animals. (A) Quantitative analysis indicated that incisional surgery reduced the levels of expression of mRNAs encoding GABA receptors, but not of those encoding opioid receptors. (B) Carrageenan injection increased expression of opioid receptor mRNAs, but not those of GABA receptor mRNAs. *P < 0.05, †P < 0.01, ‡P < 0.001 compared to naïve rats.
|
Intrathecal curcumin dose-dependently increased the withdrawal thresholds of incised (P < 0.01 at the 100 μg vs. vehicle) and carrageenan-injected (P < 0.05 at the 300 μg vs. vehicle) paws (Figs. 3 and 4). The ED50 values (with 95% CIs) of curcumin used to alleviate postoperative and inflammatory pain were 22.3 μg (12.2–40.8 μg) and 126.5 μg (59.6–268.6 μg), respectively.
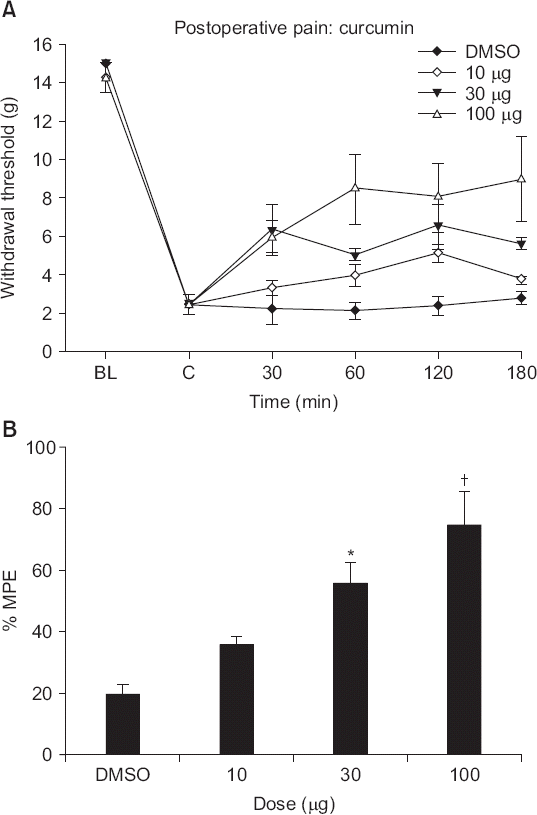 | Fig. 3(A) The time effect and (B) dose-response of intrathecal curcumin on the hindpaw withdrawal response to von Frey filament pressure applied after incisional surgery. Data are presented as withdrawal thresholds or percentages of maximal possible effects (%MPEs). Each line represents the mean ± SEM of data from five-to-seven rats. Control data were obtained 2 h after incision. Curcumin was administered immediately after measurement of control post-incisional thresholds. BL: baseline withdrawal threshold measured before paw incision. Curcumin produced dose-dependent increases in withdrawal thresholds of injured paws after incision. *P < 0.05, †P < 0.01 compared to vehicle (dimethylsulfoxide, DMSO) alone.
|
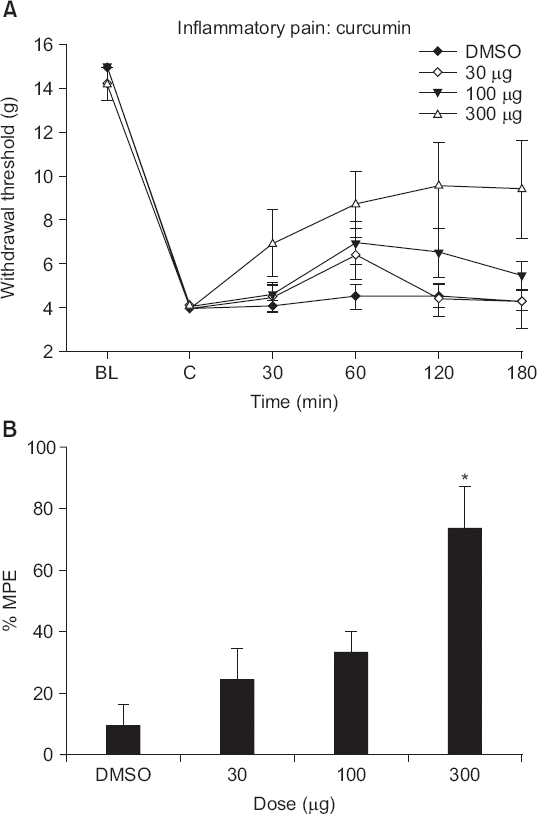 | Fig. 4(A) The time effect of and (B) dose-response to intrathecal curcumin, assessed by measuring hindpaw withdrawal responses to von Frey filament pressure applied after carrageenan injection. Data are presented as withdrawal thresholds or percentages of maximal possible effects (%MPEs). Each line represents the mean ± SEM of data from five-to-seven rats. Control data were obtained 4 h after carrageenan injection. Curcumin was administered immediately after measurement of control post-injectional thresholds. BL: baseline withdrawal threshold measured before carrageenan injection. Curcumin caused a dose-dependent increase in the withdrawal thresholds of injected paws after carrageenan injection. *P < 0.05 compared to vehicle (dimethylsulfoxide, DMSO) alone.
|
The antinociceptive effect of intrathecal curcumin on postoperative pain induced by incisional surgery was antagonized by intrathecal bicuculline (a GABA-A receptor antagonist, P < 0.01 vs. curcumin) and saclofen (a GABA-B receptor antagonist, P < 0.001 vs. curcumin) (Fig. 5A). Additionally, intrathecal curcumin administration increased the levels of mRNAs encoding both GABA-A (P < 0.01 vs. incised) and GABA-B (P < 0.001 vs. incised) receptors in the spines of incised rats (Fig. 5B). Intrathecal CTOP (a μ opioid receptor antagonist, P < 0.001 vs. curcumin), naltrindole (a δ opioid receptor antagonist, P < 0.001 vs. curcumin), and GNTI (a κ opioid receptor antagonist, P < 0.001 vs. curcumin) reversed the antinociceptive effect of curcumin on incisional pain (Fig. 6A), but intrathecal curcumin did not affect the levels of spinal cord opioid receptors (Fig. 6B).
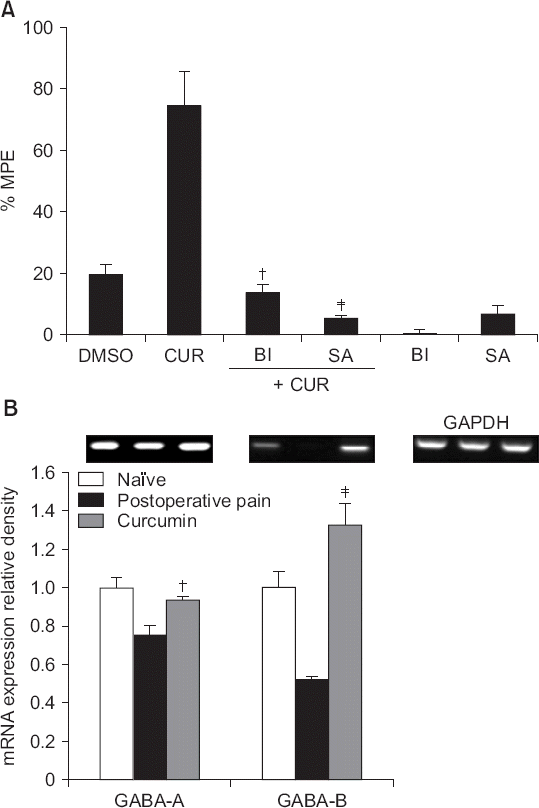 | Fig. 5(A) The effects of intrathecal bicuculline (a γ-aminobutyric acid [GABA]-A receptor antagonist, 0.3 μg), and saclofen (a GABA-B receptor antagonist, 100 μg), on the antinociceptive effects of intrathecal curcumin (100 μg) in rats experiencing postoperative pain; and (B) the effects of intrathecal curcumin (100 μg) on the expression levels of mRNAs encoding GABA receptors in the left dorsal horn of the spinal cord in incised rats. Data are presented as percentages of maximal possible effects (%MPEs) or the relative values of GABA receptor levels to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Each bar represents the mean ± SEM of data from four-to-six rats. DMSO: dimethylsulfoxide, CUR: curcumin, BI: Bicuculline, SA: saclofen. (A) Both bicuculline and saclofen reversed the effects of curcumin, †P < 0.01, ‡P < 0.001 compared with curcumin. (B) Intrathecal curcumin increased the expression levels of mRNAs encoding GABA receptors, †P < 0.01, ‡P < 0.001 compared with the incised group.
|
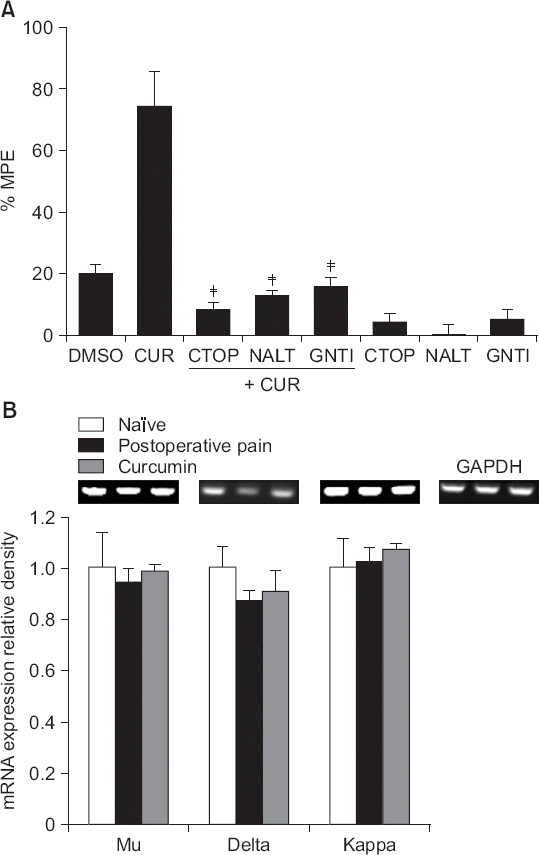 | Fig. 6(A) The effects of intrathecal CTOP (a μ opioid receptor antagonist, 15 μg), naltrindole (a δ opioid receptor antagonist, 10 g), and GNTI (a κ opioid receptor antagonist, 50 μg) on the antinociceptive effects of intrathecal curcumin (100 μg) on postoperative pain, and (B) the effects of intrathecal curcumin (100 μg) on the expression levels of mRNAs encoding opioid receptors in the left dorsal horn of the spinal cord in incised rats. Data are presented as percentages of maximal possible effects (%MPEs) or the relative expression levels of opioid receptors compared to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Each bar represents the mean ± SEM of data from four-to-six rats. DMSO: dimethylsulfoxide, CUR: curcumin, NALT: naltrindole. (A) CTOP, naltrindole and GNTI reversed the effects of curcumin, ‡P < 0.001 compared with curcumin. (B) Intrathecal curcumin only marginally affected the expression levels of mRNAs encoding opioid receptors in the incised group.
|
The antinociceptive effect of intrathecal curcumin on inflammatory pain induced by carrageenan injection was antagonized by intrathecal CTOP, naltrindole, and GNTI (Fig. 7A). Additionally, intrathecal curcumin decreased the levels of mRNAs encoding the three opioid receptors in the spines of carrageenan-injected rats (Fig. 7B). Intrathecal bicuculline and saclofen reversed the antinociceptive effect of intrathecal curcumin on inflammatory pain (Fig. 8A), but intrathecal curcumin did not affect the levels of mRNAs encoding spinal GABA receptors (P > 0.05 vs. incised, Fig. 8B).
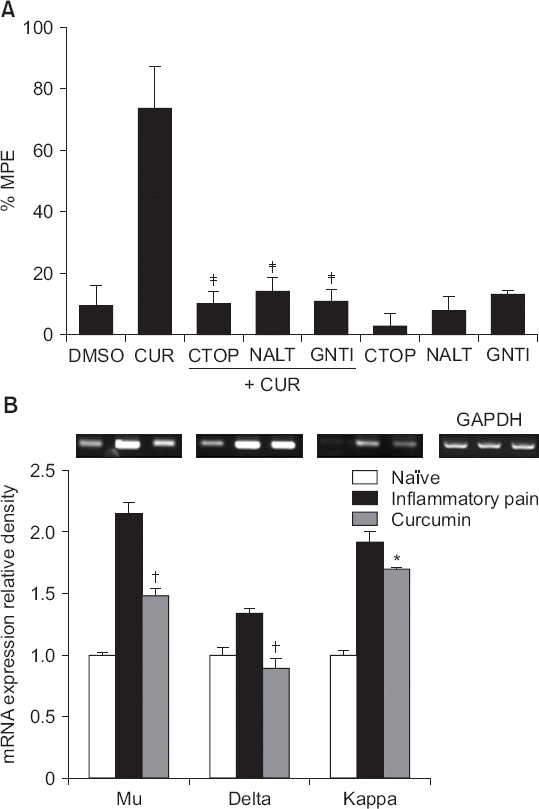 | Fig. 7(A) The effects of intrathecal CTOP (a μ opioid receptor antagonist, 15 μg), naltrindole (a δ opioid receptor antagonist, 10 μg) and GNTI (a κ opioid receptor antagonist, 50 μg) on the antinociceptive effects of intrathecal curcumin (300 μg) on inflammatory pain, and (B) the effects of intrathecal curcumin (300 μg) on the expression levels of mRNAs encoding opioid receptors in the left dorsal horn of the spinal cord in carrageenan-injected rats. Data are presented as percentages of maximal possible effects (%MPEs) or the relative expression levels of opioid receptors compared to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Each bar represents the mean ± SEM of data from four-to-six rats. DMSO: dimethylsulfoxide, CUR: curcumin, NALT: naltrindole. (A) All of CTOP, naltrindole, and GNTI reversed the effects of curcumin. ‡P < 0.001 compared with curcumin. (B) Intrathecal curcumin decreased the expression levels of mRNAs encoding all three opioid receptors, *P < 0.05, †P < 0.01, compared to the inflammatory group.
|
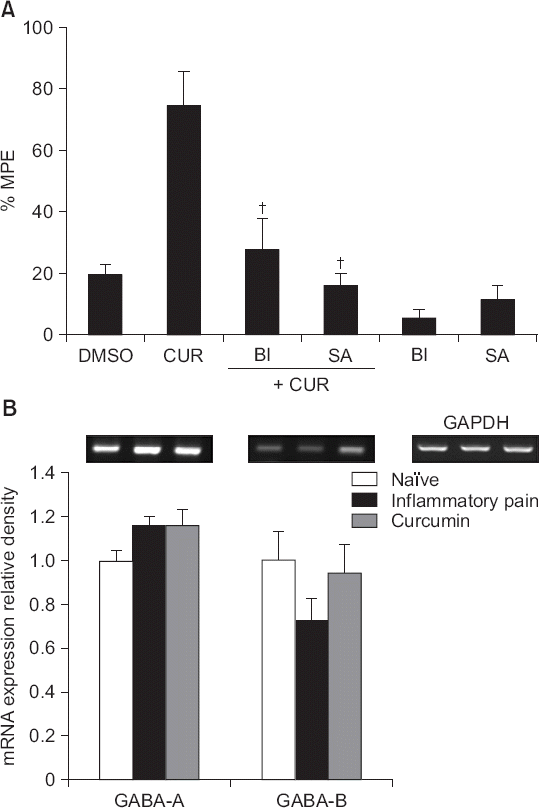 | Fig. 8(A) The effects of intrathecal bicuculline (a γ-aminobutyric acid [GABA]-A receptor antagonist, 0.3 μg) and saclofen (a GABA-B receptor antagonist, 100 μg) on the antinociceptive effects of intrathecal curcumin (300 μg) on inflammatory pain, and (B) the effects of intrathecal curcumin (300 μg) on the expression levels of mRNAs encoding GABA receptors in the left dorsal horn of the spinal cord in carrageenan-injected rats. Data are presented as percentages of maximal possible effects (%MPEs) or the relative expression levels of GABA receptors to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Each bar represents the mean ± SEM of data from four-to-six rats. DMSO: dimethylsulfoxide, CUR: curcumin, BI: Bicuculline, SA: saclofen. (A) Both bicuculline and saclofen reversed the effects of curcumin. †P < 0.01 compared with curcumin. (B) Intrathecal curcumin only marginally affected the expression levels of mRNAs encoding GABA receptors in the inflammatory group.
|
The placing-stepping reflex, the righting reflex, and the pinna and corneal reflexes were present after intrathecal administration of curcumin at the highest doses used.
Go to :
DISCUSSION
We showed, in the present study, that incisional surgery and carrageenan injection decreased withdrawal thresholds in the ipsilateral, but not contralateral, paws. These findings are consistent with those of previous reports [9,10], validating the postoperative and inflammatory pain models used in the present study. In addition, RT-PCR revealed that both GABA (A and B) and opioid receptors (μ, δ, and κ) were expressed in the spinal cords of naïve rats, in line with previous findings [14,15]. Both the GABA and opioid systems are major contributors to nociceptive processing in the spinal cord [4], and we thus focused on these two systems. Several lines of evidence indicate that GABA and opioid receptor agonists exert antinociceptive actions at the spinal level. Intrathecal GABA receptor agonists decreased complete Freund’s adjuvant-induced mechanical hyperalgesia [12], mechanical allodynia in neuropathic pain [16], and incisional pain [17]. Intrathecal opioid receptor agonists also triggered antinociceptive activity in animals subjected to incisional and formalin-induced [13,18]. In the present study, the levels of mRNAs encoding spinal GABA and opioid receptors decreased or increased (respectively) after incisional surgery and carrageenan injection. However, neither incisional surgery nor carrageenan injection materially affected the levels of spinal opioid or GABA receptors. Previous molecular biological studies of incisional and inflammatory pain have yielded a variety of results. The expression levels of spinal cord GABA-A, and opioid μ and δ receptors, were not affected by incision [17,19]. GABA-B receptor binding was reduced after injection of complete Freund’s adjuvant, whereas that of the GABA-A receptor was unchanged [20]. No changes in the levels of mRNAs encoding the μ, δ, or κ opioid receptors were observed in the spinal cord after injection of formalin [21]. The expression levels of the μ, δ, and κ opioid receptors increased after injection of complete Freund’s adjuvant [22,23]. Such findings suggest that the GABA and opioid receptor systems may be differentially activated depending on the form of nociception required at the spinal level. Our present data allow us to advance certain suggestions. Reduction of GABA receptor expression in rats experiencing postoperative pain suggests that acute pain per se, inflicted (for example) by incisional surgery, may inhibit GABA receptor expression in the spinal cord. Thus, high doses of spinal GABA receptor agonists may be necessary to treat postoperative pain. On the other hand, the increased levels of opioid receptors evident after carrageenan injection suggests that inflammatory pain per se may stimulate spinal cord opioid receptor expression in an effort to reduce pain, and it may thus be possible to relieve pain using small doses of opioids. Interestingly, the spinal cord levels of neither the opioid nor the GABA receptors were altered upon infliction of postoperative or inflammatory pain, suggesting that receptor levels may not be directly affected by pain.
In the present study, intrathecal curcumin exhibited antinociceptive effects on postoperative and inflammatory pain induced by incision and carrageenan injection, respectively. Previous studies have revealed the antinociceptive actions of curcumin. Intrathecal curcumin attenuated formalin-induced nociception, spinal nerve ligation-induced neuropathic pain and complete Freund’s adjuvant-induced pain hypersensitivity, which was mediated through the inhibition of glial activation [5,6,24]. In addition, oral curcumin reduced the formalin-induced flinching response, thermal hyperalgesia and neuropathic pain [25–27]. Intravenous curcumin attenuated visceral nociception [28]. Intraperitoneal curcumin and a synthetic curcuminoid analog reduced both formalin- and thermally induced nociception and neuropathic pain [29,30]. An interesting finding of the current study was that curcumin action varied depending on the type of nociception analyzed. The ED50 for inflammatory pain was five-fold higher than that for postoperative pain, suggesting that curcumin may be considerably more effective when used to treat postoperative rather than inflammatory pain.
In the current study, the antinociceptive effects of intrathecal curcumin on postoperative and inflammatory pain were antagonized by intrathecal GABA (A and B) and opioid (μ, δ, and κ) receptor antagonists. However, the RT-PCR data differed from those of the behavioral study. Intrathecal curcumin elevated the levels of GABA receptors, but not opioid receptor levels, when postoperative pain was induced. In contrast, intrathecal curcumin decreased opioid receptor levels, but not the levels of GABA receptors, when inflammatory pain was inflicted. In addition, we found that incisional surgery affected GABA receptor levels only, and carrageenan injection changed opioid receptor levels only. Together, these findings suggest that postoperative pain may be modulated by spinal cord GABA rather than opioid receptors. Additionally, curcumin may directly affect the synthesis of mRNAs encoding the spinal GABA receptors that mediate antinociception of postoperative pain. Opioid receptors may play indirect roles only in this context, when it is considered that no changes in the expression levels of opioid receptor mRNAs were noted despite the fact that curcumin activity was antagonized by opioid receptor blockers. On the other hand, the roles played by GABA and opioid receptors upon infliction of inflammatory pain may be the opposite of those under conditions of postoperative pain. This is the first work to clarify the pharmacologic mechanisms of curcumin at the spinal level. In general, it seems likely that the effects of curcumin for postoperative and inflammatory pain are mediated by both GABA and opioid receptors of the spinal cord. However, these receptors may be differentially involved depending on the type of pain inflicted. Because not all observed phenomena could be explained in detail, further research is needed.
The present study had several limitations. First, RT-PCR was performed at only one time point. The expression levels of spinal GABA and opioid receptors may vary over time. However, the selected time point was that at which the withdrawal thresholds were maximally decreased after incision and carrageenan injection. Second, we did not determine how curcumin acted on spinal GABA and opioid receptors. Moreover, the mechanism by which curcumin may exhibit antinociceptive activities without altering receptor expression levels was not investigated. Thus, further studies are required.
Together, the data suggest that postoperative pain caused by incisional surgery decreases spinal cord GABA receptor expression, whereas inflammatory pain caused by carrageenan injection increases such receptor expression. Intrathecal curcumin attenuates both postoperative and inflammatory pain. Additionally, spinal cord GABA and opioid receptors are directly involved in the antinociceptive actions of curcumin on postoperative and inflammatory pain, respectively. However, spinal cord opioid receptors may also play indirect roles in mediating curcumin antinociception of postoperative pain, and spinal cord GABA receptors may be indirectly responsible for curcumin antinociception of inflammatory pain. In general, curcumin may be a useful therapeutic option for management of postoperative and inflammatory pain at the spinal level.
Go to :
 XML Download
XML Download