

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Aminoglycoside antibiotics have been known to be very effective in treating Gram-negative bacteria [1-3]. However, although such antibiotics are effective and are widely available, they have long been known to exhibit nephrotoxicity and ototoxicity [1-3]. The former is known to be often reversible, while the latter is irreversible. The ototoxic effects of these drugs include sensorineural hearing loss, which is permanent because the hair cells in the cochlea do not regenerate [3]. The incidence of ototoxicity from aminoglycosides has been reported to range from 2% to 25%, which is likely underestimated because of the ambiguity of the symptoms [2].
The generation of reactive oxygen species (ROS) is one of the earliest events in the process of aminoglycoside ototoxicity. Production of ROS is thought to play a key role in the initiation of hair cell death via oxidative stress [1,4]. Histopathologic studies have shown that the outer hair cells are more sensitive to ototoxic injury than the inner hair cells [2,3]. Such histological findings resemble those of apoptotic cell death, which is primarily regulated by the activation of caspases through either internal or external pathways [3,5,6]. Despite the protective effects against oxidative stress of many agents that have been studied [3,5,7], none has been approved for use in clinical fields.
Trimetazidine (TMZ), 1-2,3,4 tri-metoxibencil piperazine dihydrochloride, is a clinically available drug used for ischemic heart disease because it prevents cell death secondary to transient myocardial ischemia that occurs during reperfusion [8,9]. Numerous experimental studies have reported that TMZ prevents an excessive release of free radicals, which are particularly toxic to phospholipid membranes and are responsible for the decrease in intracellular adenosin triphosphate (ATP) concentration and the extracellular leakage of potassium and protects mitochondrial function [8,10,11]. Zebrafish have hair cells on the surfaces of their bodies that function as part of the lateral line sensory system [12,13]. The hair cells of neuromasts in the zebrafish lateral line are a useful system for studying hair cell loss after exposure to ototoxic agents [14,15]. Thus, the purpose of the current study was to investigate the effects of TMZ on neomycin-induced ototoxicity in transgenic zebrafish.
Go to : 
MATERIALS AND METHODS
Fish preparation and maintenance
Transgenic zebrafish embryos were produced using paired matings of adult fish maintained at 28.5℃ in a zebrafish facility at Korea University Ansan Hospital. Embryos were maintained at a density of about 50 per 100 mm2 Petri dish in embryo media (1 mM MgSO4, 120 µM KH2PO4, 74 µM Na2HPO4, 1 mM CaCl2, 500 µM KCl, 15 µM NaCl, and 500 µM NaHCO3 in dH2O).
Examination of neuromasts in zebrafish
Neomycin solutions were prepared by adding the neomycin powder to the embryo medium. Neomycin has been shown to decrease the viability of neuromasts in a dose-dependent manner. Additionally, a preliminary study showed that neuromasts treated with 125 µM neomycin for 1 hour show a viability rate of 50% (data is not shown). Thus, a dose of 125 µM neomycin was chosen as an adequate experimental condition for our following studies. For the present experiment, 10 fish were tested at each of the following concentrations of TMZ and repeated three times. The larvae of 5-dpf transgenic zebrafish were simultaneously exposed to 125 µM neomycin and one of the following concentrations of TMZ for 1 hour: 10 µM, 100 µM, 500 µM, 1,000 µM, 1,500 µM, or 2,000 µM. After the transgenic zebrafish larvae were exposed to neomycin and TMZ, they were rinsed three times in embryo media and anesthetized with Tricane (3-aminobenzoic acid, 0.4 g/ethyl ester, 100 mL - pH 7, adjustments by Tris buffer) for 5 minutes.
The zebrafish were mounted with methylcellulose on a depression slide for evaluation under a fluorescence microscope. Hair cells within the neuromasts of the supraorbital (SO1 and SO2), otic (O1), and occipital (OC1) lateral lines were analyzed [12,13,16,17]. The average numbers of hair cells of the SO1, SO2, O1, and OC1 neuromasts were counted for each zebrafish under all experimental and control conditions (n=10) using a fluorescence microscope and confocal microscope (LSM5 PASCAL; Carl Zeiss, Oberkochen, Germany). The fluorescent dye, 2-[4-(dimethylamino) styryl]-N-ethylpyridinium iodide (DASPEI; Invitrogen, Carlsbad, CA, USA), was used as to stain mitochondria within hair cells. Larvae were incubated in the embryo medium containing 0.005% DASPEI for 15 minutes and were analyzed according to the same method as described above.
Terminal deoxynucleotidyl transferase (TdT)-mediated dUTP-biotin nick end labeling (TUNEL) assay
Apoptosis in the neuromasts was evaluated using the TUNEL method with an in situ cell detection kit (Roche Molecular Biochemicals, Mannheim, Germany), as per the manufacturer's protocol. The transgenic zebrafish larvae were exposed to medium containing 125 µM neomycin and 1,000 µM TMZ for 1 hour. Thereafter, the larvae were washed with phosphate buffered saline (PBS) and fixed in 4% paraformaldehyde. The larvae were then incubated with 50 µL TUNEL reaction mixture (TdT and fluorescein-dUTP) at 37℃ for 1 hour in a humid atmosphere. The zebrafish were then evaluated under a fluorescence microscope (AxioCam MRc5; Carl Zeiss) [16].
Scanning electron microscopy
For scanning electron microscopy (SEM), 5-dpf zebrafish larvae exposed to 125 µM neomycin and 1,000 µM TMZ for 1 hour were prefixed by immersion in 2% glutaraldehyde in 0.1 M phosphate buffer and postfixed for 2 hours in 1% osmic acid dissolved in PBS. Samples were treated in a graded series of ethanol and t-butyl alcohol, dried in a freeze dryer (Hitachi, ES-2030; Hitachi, Tokyo, Japan), platinum-coated using an ion coater (Eiko, IB-5; Eiko, Katsuda, Japan), and observed under an field emission-SEM (Hitachi, S-4700) [18].
Statistical analysis
All values are shown as mean±SD. For statistical comparisons, one-way analysis of variance was used for multiple comparisons. Results were considered statistically significant at P<0.05.
Go to :
RESULTS
The effect of TMZ on hair cells in transgenic zebrafish treated with neomycin
Four neuromasts, SO1, SO2, O1, and OC1, were evaluated. Treatment of the zebrafish with 125 µM neomycin for 1 hour significantly decreased the number of hair cells in the neuromasts. Hair cells from the four neuromasts were counted to investigate the changes after treatment with neomycin and TMZ.
Confocal microscopy was used to calculate hair cell survival as a percentage of the hair cells present in the control group not exposed to neomycin. While 125 µM neomycin significantly decreased the number of neuromast hair cells, TMZ protected against this neomycin-induced hair cell loss (control, 13.1±0.9 cells; 125 µM neomycin only, 4.2±0.5 cells; TMZ 10 µM, 4.2±0.7 cells; TMZ 100 µM, 4.1±0.6 cells; TMZ 500 µM, 9.4±1.1 cells; TMZ 1,000 µM, 11.2±0.4 cells; TMZ 1,500 µM, 10.0±0.7 cells; TMZ 2,000 µM, 8.7±1.7 cells; n=10; P<0.05) (Figs. 1, 2).
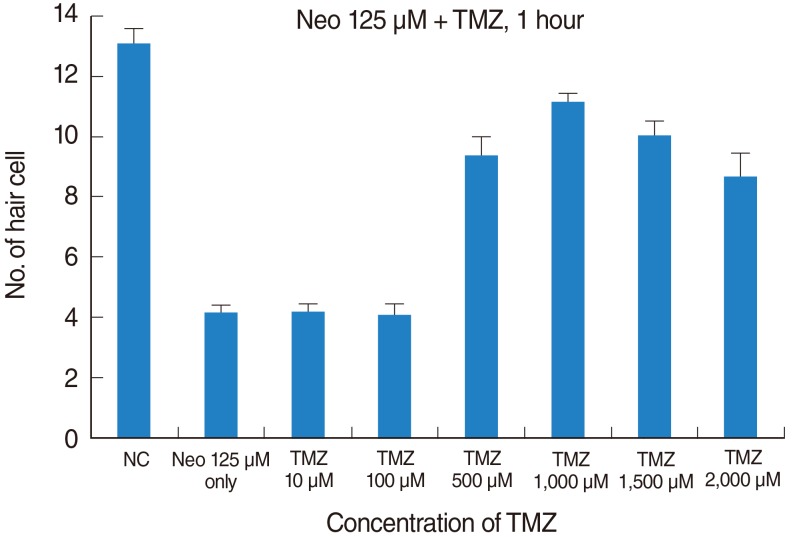 | Fig. 1Quantitative assay of neuromasts in zebrafish. Hair cells from four neuromasts (supraorbital 1, supraorbital 2, otic 1, and occipital 1 were counted. Treatment of zebrafish with 125 µM Neo for 1 hour significantly decreased the number of hair cells in the neuromasts. TMZ protected against neomycin-induced hair cell loss in neuromasts (NC, 13.1±0.9 cells; 125 µM Neo only 4.2±0.5 cells; TMZ 10 µM, 4.2±0.7 cells; TMZ 100 µM, 4.1±0.6 cells; TMZ 500 µM, 9.4±1.1 cells; TMZ 1,000 µM, 11.2±0.4 cells; TMZ 1,500 µM, 10.0±0.7 cells; TMZ 2,000 µM, 8.7±1.7 cells; n=10; P<0.05). NC, negative control; Neo, neomycin; TMZ, trimetazidine.
|
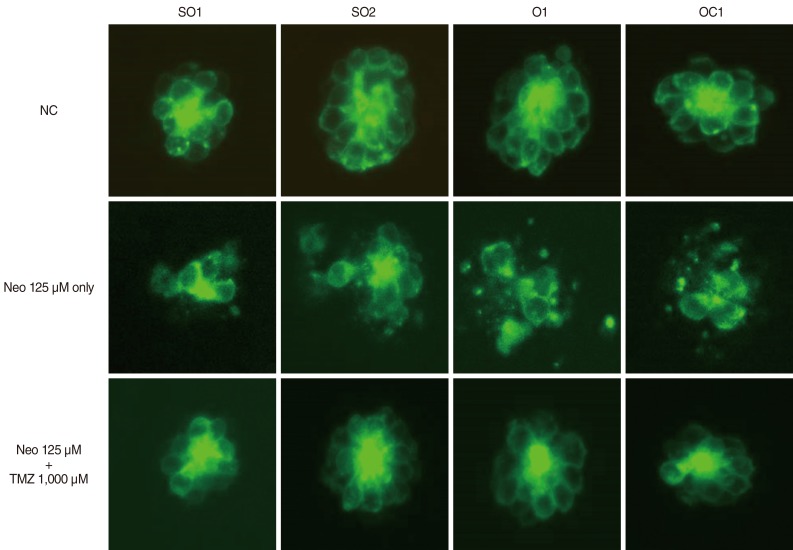 | Fig. 2Confocal microscopy (×40). The 5-dpf transgenic zebrafish were treated simultaneously with 125 µM Neo and 1,000 µM TMZ for 1 hour. Treatment with 125 µM Neo resulted in a significant decrease in the number of hair cells in the neuromasts, and TMZ protected against this neomycin-induced hair cell damage. TMZ, trimetazidine; SO1, supraorbital 1; SO2, supraorbital 2; O1, otic 1; OC1, occipital 1; NC, negative control; Neo, neomycin.
|
Because 1,000 µM TMZ was the most effective dose, this concentration was used in the DASPEI and TUNEL assays. DASPEI staining was performed to evaluate mitochondrial damage and revealed that mitochondria within hair cells were protected by TMZ treatment (Fig. 3). The TUNEL reaction was used to determine if neomycin induced the death of neuromasts by apoptosis and whether that cell death could be prevented by TMZ treatment. As shown in Fig. 4, neomycin increased TUNEL-positive cells, and TMZ treatment decreased TUNEL-positive cells. These results indicate that neomycin promotes apoptotic cell death, and that this apoptosis can be inhibited by TMZ treatment.
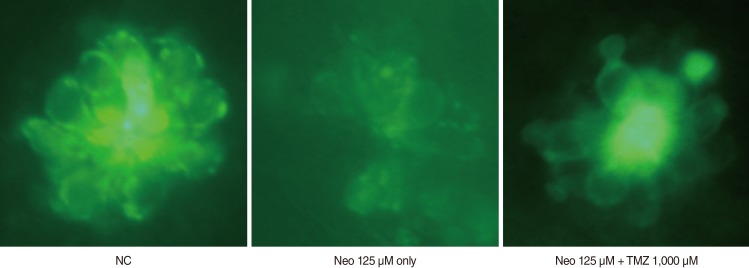 | Fig. 3Evaluation of mitochondrial damage with DASPEI (fluorescent microscopy, O1, ×40). The 5-dpf wild-type zebrafish were treated with 125 µM Neo and 1,000 µM TMZ for 1 hour. Treatment with 125 µM Neo resulted in a significant decrease in mitochondrial staining with DASPEI, and this mitochondrial damage was prevented by TMZ. NC, negative control; Neo, neomycin; TMZ, trimetazidine; O1, otic.
|
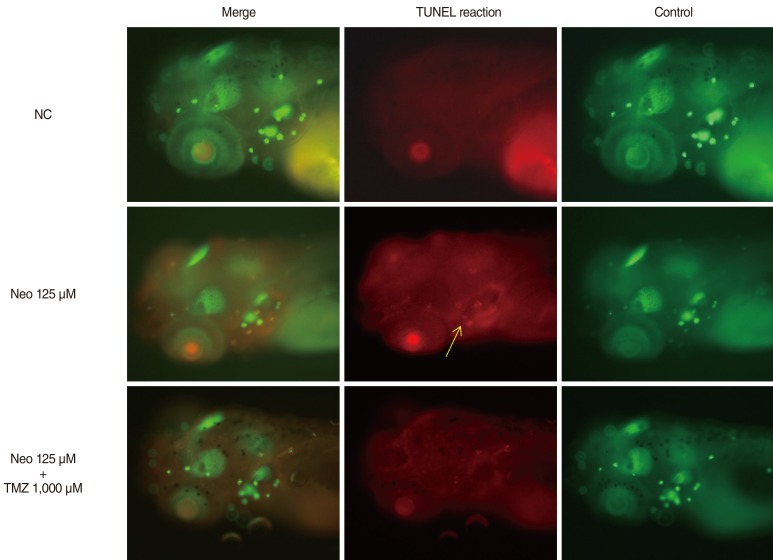 | Fig. 4Terminal deoxynucleotidyl transferase (TdT)-mediated dUTP-biotin nick end labeling (TUNEL) assay (fluorescent microscope, ×10). Neo-induced apoptotic cells were confirmed by the TUNEL assay. Apoptotic cells are marked as light red dots in red-colored fish after the TUNEL reaction under a fluorescent microscope (arrow indicates TUNEL-positive cells). No light-red dots were found in the NC group. A comparison of the color intensity between the group treated with 125 µM Neo and the group treated with 1,000 µM TMZ for 1 hour showed that TMZ significantly decreased the number of TUNEL-positive cells and protected against Neo-induced apoptotic cell death of the neuromast hair cells (green-colored fish of right column, control for TUNEL reaction; red-colored fish of middle column, TUNEL reaction; green- and red-colored fish of left column, combination of control and TUNEL reaction). NC, negative control; Neo, neomycin; TMZ, trimetazidine.
|
TMZ protects stereocilia and kinocillium in transgenic zebrafish treated with neomycin
Fig. 5 shows scanning electron micrographs of stereocilia and kinocilium from neomycin- and TMZ-treated zebrafish. Compared to control larvae, exposure to 125 µM neomycin for 1 hour resulted in severe morphological damage to hair cells, including loss or fusion of stereocilia and kinocilium. TMZ protected the larvae against neomycin-induced hair cell loss compared to treatment with neomycin alone.
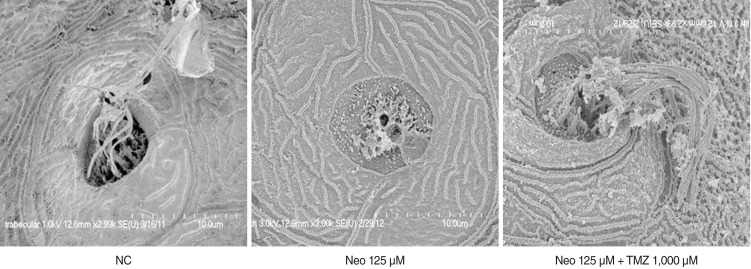 | Fig. 5Scanning electron microscopy (SEM; O1, ×3,000). Compared to control neuromasts, neuromasts in zebrafish treated with 125 µM Neo for 1 hour showed severe morphological damage to stereocilia and kinocilium under SEM. However, this damage was prevented by TMZ. NC, negative control; Neo, neomycin; TMZ, trimetazidine, O1, otic.
|
Go to :
DISCUSSION
The current study evaluated the effect of TMZ to prevent neomycin-induced ototoxicity. Results indicated that concomitant administration of TMZ prevents neomycin-induced hair cell damage in zebrafish. It is known that aminoglycosides appear to generate free radicals within the inner ear, with subsequent permanent damage to sensory cells, resulting in permanent hearing loss [3]. The production of ROS is known to be the initiating step of aminoglycoside ototoxicity, which ultimately results in apoptosis [1-3,6]. ROS is thought to occur as a result of aminoglycoside affinity for iron. The drug binds with iron, and the resulting iron-aminoglycoside complex is known to promote the formation of free radicals [19]. Free radicals are known to cause damage to many cells in the body in situations such as inflammation, aging, and reperfusion injury [2,19]. Free radicals can attack a variety of cell components ranging from cell membranes to DNA, causing irreversible damage and cell death. Morphologic features of apoptosis are condensed chromatin, nuclear pyknosis, apoptotic body formation, and cellular dissolution [2]. Numerous studies have attempted to identify a mechanism that protects the inner ear from aminoglycoside ototoxicity. Unfortunately, the United States Food and Drug Administration (FDA) has not yet approved any drugs for the prevention of drug-induced ototoxicity [16]. TMZ is a clinically useful drug used for ischemic heart disease since 1970. It was also demonstrated that TMZ is affective in hepatic ischemia and neurotoxicity caused by cyclosporine [10]. TMZ has been known to decrease the intracellular accumulation of sodium and calcium and to inhibit platelet adhesion-aggregation, neutrophil infiltration, and the generation or activity of oxygen-derived free radicals [9,20,21]. Additionally, TMZ reduces the vasoconstriction mediated by the components released by free radicals. Some researchers have reported a protective effect of TMZ on ROS-induced renal injury during ischemia/reperfusion [9,22]. However, studies of TMZ on hair cell damage are rare.
The use of the zebrafish in a chemical screen was first described by identification of small molecules that modulate development of the cardiovascular system, central nervous system, neural crest, and ear [23,24]. Subsequent chemical screens have demonstrated a number of different compounds, including novel retinoids, drugs that affect heart rate in a zebrafish mode, chemicals that inhibit melanoma [24-27]. Recently, zebrafish have emerged as a model for studies of hearing and vestibular function [28]. Zebrafish have hair cells that are functionally and structurally similar to mammalian inner ear hair cells. These hair cells are organized into clusters of 5-20 cells, called neuromasts, and are located on the head and along the body (lateral line) [12,13]. The superficial location of hair cells permits easy visualization and manipulation of hair cells in living animals [1]. Hair cells of the lateral line allow for rapid in vivo assessment of hair cell death and ototoxic and potentially protective compounds [14-16,25,28]. However, it is important to know some critical differences between zebrafish hair cells and mammalian hair cells. Hair cells of the lateral line lacks stria vascularis in contrast to the inner ear, thus hair cell loss through strial mechanisms cannot be elucidated [12,24,29]. In addition, lateral line hair cells are morphologically and physiologically much more similar to vestibular than cochlear hair cells of the mammalian inner ear. Therefore, any discovery made in zebrafish must be confirmed in mammals prior to translational applications [12,24,29]. In the present study, transgenic zebrafish (Brn3C: EGFP) were used to determine the potential ototoxicity of drugs and to investigate the protective activity of TMZ with regard to neomycin-induced ototoxicity. These transgenic zebrafish have green neuromasts that are visible under a fluorescent microscope without staining [16,30]. Exposure to 125 µM neomycin for 1 hour resulted in apoptotic cell death, reduction in mitochondrial injury, and severe morphological damage to stereocilia and kinocilium. However, TMZ provided nearly complete protection against neomycin-induced hair cell damage. These results suggest that neomycin induces apoptosis, and that apoptotic cell death was prevented with TMZ treatment.
In conclusion, TMZ attenuated neomycin-induced hair cell damage by reducing the amount of free radicals. Further studies are currently ongoing to define the functional effects of TMZ in vivo.
Go to :
 XML Download
XML Download