

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
In the mammalian cochlea, there are two types of hair cells that subserve distinct functions and receive characteristic patterns of innervations. Inner hair cells (IHCs) receive nearly all afferent innervations and are primary acoustic transducers. The three IHC potassium currents are distinguishable by their pharmacology and their activation kinetics [1-3]. The fast activating current, IK,f, is blocked by tetraethylammonium (TEA) but is resistant to 4-aminopyridine (4-AP). IK,s is activated more slowly on depolarization and is blocked by 4-AP but not by TEA. Another potassium current, IK,n, is already activated at the resting potential of the cell and thus determines the membrane potential and membrane constant. To date, most presently available biophysical studies of IHC potassium currents have been carried out at room temperature. This limits meaningful comparisons between in vivo findings and in vitro experiments such as the whole-cell configuration.
Neurosecretion has been shown to be highly temperature sensitive in various preparations. The kinetics of endocytotic membrane retrieval following transmitter release was described as being highly temperature sensitive [4,5]. Similarly in mice IHCs, temperature enhances IHC presynaptic function [6]. The number of readily releasable vesicles available at the active zone is higher at physiological temperatures (35℃-37℃) with no significant change in the rate of sustained release, suggesting that experimental results acquired at room temperature should be scaled up if they are to be related to in vivo findings. Until recently, most patch-clamp recordings in IHCs were performed at room temperature. There are several reports in which experiments were performed in the physiological temperature [1,3], but they did not precisely mentioned about the kinetics of potassium currents. Potassium currents are known to participate in repolarization and discharge behaviors of action potentials in neurons [7,8], so changes in IHC potassium currents at physiological temperatures may cause enhanced IHC presynaptic function.
In the present study, IHCs were isolated from a mature guinea-pig cochlea and potassium currents were recorded at room temperature (around 25℃) and physiological temperatures (35℃-37℃). The effect of temperature on the amplitude of currents and kinetics, i.e., activation and inactivation, were investigated.
Go to : 
MATERIALS AND METHODS
Preparation of isolated IHCs
Adult albino guinea-pigs (200-350 g) with a normal Preyer reflex were killed by rapid cervical dislocation, both bullae were removed, and the cochlea exposed. The cochlea, fused to the bulla, was placed in a Ca2+-free external solution (mM: 142 NaCl, 4 KCl, 3 MgCl2, 2 NaH2PO4, 8 Na2HPO4, adjusted to pH 7.4 with NaOH). The otic capsule was opened, allowing removal of the organ of Corti attached to the modiolus. The organ of Corti was treated with trypsin (0.5 mg/mL, T-4665, Sigma-Aldrich, St. Louis, MO, USA) for 12 minutes, and gentle mechanical trituration was carried out. Trypsin was rinsed from the specimen by superfusing with a standard external solution (mM: 142 NaCl, 4 KCl, 2 MgCl2, 1 CaCl2, 2 NaH2PO4, 8 Na2HPO4, adjusted to pH 7.4 with NaOH) for at least 10 minutes before starting any experiments. The most important landmarks for identifying IHCs are a tight neck and angle between the cuticular plate and the axis of the cell as described previously [9]. In the present study, IHCs were isolated from all turns of cochlea. It is very difficult to distinguish between basal turn IHCs and apical turn IHCs by shape.
Recording procedures
Membrane currents were measured by conventional whole-cell voltage-clamp recordings using an EPC-10 (HEKA, Lambrecht, Germany). Data acquisition was controlled by the software PatchMaster (HEKA). Recording electrodes were pulled on a two-stage vertical puller (PP830, Narishige, Tokyo, Japan) using 1.5 mm outside diameter borosilicate glass (GC-1.5, Narishige) filled with an internal solution (mM: 144 KCl; 2 MgCl2; 1 NaH2PO4; 8 Na2HPO4; 2 ATP; 3 D-glucose; 0.5 EGTA; adjusted to pH 7.4 with potassium hydroxide [KOH]). Pipettes showed a resistance of 4-8 MΩ in the bath and were coated with ski wax (Tour-DIA, DIAWax, Otaru, Japan) to minimize capacitance. The cell's capacitance was 10.1±4.1 pF (mean±SD) and the series resistance was 14.0±4.9 MΩ (n=28). Cells were continuously perfused with external saline to avoid evaporation. An uncompensated series resistance causes the membrane voltage (VM) to deviate from the command voltage (VC). Voltage amplitudes were corrected for this error using the following formula: VM=VC - IMRS, where IM is the membrane current (before leak subtraction). Point-by-point current-voltage (I-V) curves were constructed and linear interpolation of the I-V plots allowed reconstruction of the corrected current at a fixed membrane potential [10].
To obtain TEA-sensitive potassium currents, 25 mM TEA solution (T-2265, Sigma-Aldrich; replacing 25 mM NaCl in the standard external solution) was applied under pressure (PMI-200, Dagon, Minneapolis, MN, USA) using pipettes with a tip diameter of 2-4 µm positioned around 50 µm from the IHCs. TEA-sensitive currents were reconstructed by subtracting the current recorded in TEA from that recorded in standard saline.
Temperature control
Temperature was adjusted by heating the microscope stage with a dual channel temperature controller (DTC-200, Dagon). Temperature was measured by a miniature thermistor close to the IHC. The influence of temperatures on potassium current properties was first investigated on two different populations of IHCs (at room and physiological temperature), however, to allow direct comparison between room and physiological temperature, the currents of successive changes in temperature (35℃-37℃ and 25℃-26℃) in individual cells were recorded. Since the cells were easily moved by convection of heated solution, fixed cells onto the chamber floor were selected. Most experiments were performed in higher to lower temperature to avoid moving the cells by conviction by heating.
The tesmperature dependence of potassium currents was described by its temperature coefficient (Q10), which was calculated from the van't Hoff equation: Q10=(k2 / k1)(10 / [T2-T1]), where k1 and k2 are the values of potassium currents measured at lower (T1) and higher (T2) temperatures, respectively.
Animal care
The experimental design was reviewed and approved (Accession No. A23-020-0) by the Animal Care and Use Committee, Kyushu University. All procedures were conducted in accordance with the Guidelines for Animal Care and Use Committee, Kyushu University.
Go to :
RESULTS
Potassium currents at physiological temperature
Currents in response to hyperpolarizing and depolarizing voltage steps from the holding potential of -60 mV were recorded in IHCs. Typical current records at room temperature and physiological temperature (36℃) were shown in Fig. 1. IHCs had outwardly rectifying currents (IK,f) in response to depolarizing voltage pulses, with only a slight inward current (IK,n) when hyperpolarized. Fig. 1B demonstrated the inward components by enlarging the scale. The amplitude of steady outward currents was not changed with temperature. The outward current at a membrane potential of 110 mV was 9.2±3.8 nA (n=20) at room temperature and 8.5±2.6 nA (n=11) at 36℃ (Fig. 2A). The amplitude of peak inward currents was not changed with temperature (Fig. 1B). The inward current at a membrane potential of -130 mV was 0.63±0.37 nA (n=22) at room temperature and 0.68±0.34 nA (n=11) at 36℃ (Fig. 2B). At room temperature, the currents immediately activated but successive rising phases were observed, however, at 36℃, the currents immediately reached the maximum size, suggesting very fast activation (Fig. 1A).
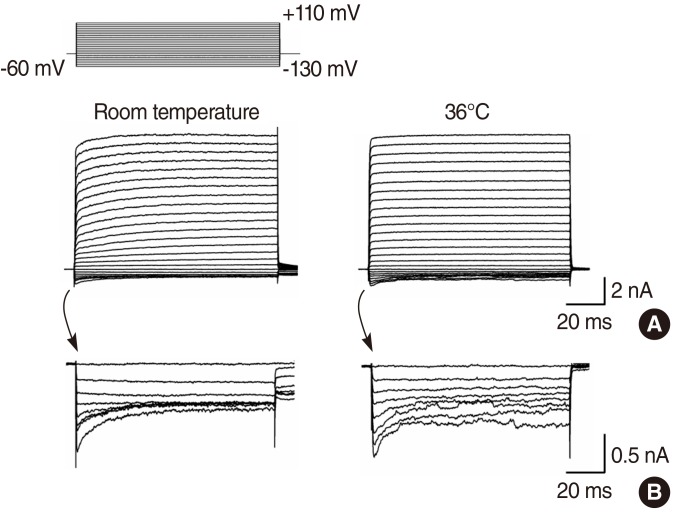 | Fig. 1Potassium currents at room temperature and 36℃. Currents in response to hyperpolarizing and depolarizing voltage steps from a holding potential of -60 mV (A, lower panel). Voltage protocol (A, upper panel). The inward component of the currents by enlarging the scale (B).
|
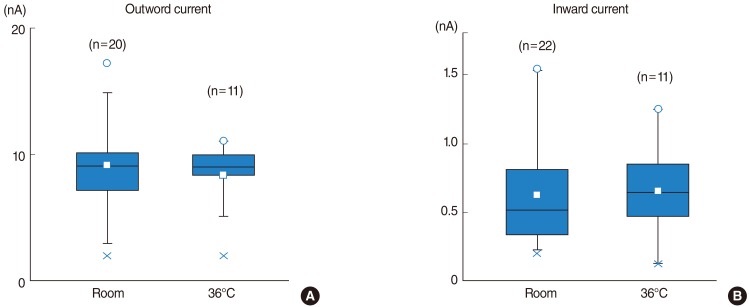 | Fig. 2Comparison between the amplitudes of potassium currents at room temperature and 36℃. (A) Amplitudes of outward currents at 110 mV. Open squares indicate the mean values at room temperature (9.2 nA ) and 36℃ (8.5 nA). (B) Amplitudes of inward currents at -130 mV. Mean values were 0.63 nA at room temperature and 0.68 nA at 36℃. Open circles were maximum values and crosses were minimum values.
|
Temperature-dependent activation kinetics
The initial activation phase of outward currents at a membrane potential of 60 mV at 35.8℃ and 25.8℃ were normalized by the maximum steady currents, and both traces were superimposed in Fig. 3. The rate of activation at 25.8℃ was slower than that recorded at physiological temperature (35.8℃). Activation kinetics is dependent on membrane potentials, becoming faster at more negative potentials. In Fig. 4, half-times (t1/2) for activation were measured at room temperature and 36℃, and plotted against various membrane potentials. This approximation is justified since there is a direct relationship between the time constant (τ) and t1/2; i.e., -t1/2=ln [1- (0.5)1/x] τ, where x=4 for activation from Hodgkin-Huxley (n4j) modeling of potassium currents [11]. The activation rate was faster at 36℃ than at room temperature. Statistical analysis (ANOVA) demonstrated significant differences between 36℃ and room temperature at membrane potentials of -10, 10, 20, 30, and 40 mV. The normalized value of half-times in activation at 36℃ relative to that at room temperature (averaged from -20 to 60 mV) was 1.94, suggesting that Q10 was 1.83 assuming a room temperature of 25℃.


Temperature-dependent inactivation kinetics
TEA-sensitive potassium currents in IHCs possess the kinetics of fast inactivation [12]. The inactivation phase could be fitted by a single exponential curve and the time constant of inactivation is dependent on membrane potentials, becoming slower at more negative potentials. Fig. 5 showed the inactivation time constants against various test potentials (from -20 to 60 mV) at room temperature and 36℃. The inactivation rate was much faster at 36℃ than at room temperature, showing significant differences at all membrane potentials. The normalized value of time constant at 36℃ relative to that at room temperature (averaged from -20 to 60 mV) was 3.58, suggesting that Q10 was 3.19. The degree of inactivation could be defined by calculating the ratio of the steady-state (Iss) to the peak (Ip) current (Iss/Ip). Fig. 6 showed Iss/Ip against various test potentials at room temperature and 36℃. The degree of inactivation was less voltage dependent and no differences are identified between 36℃ and room temperature.


Go to :
DISCUSSION
The amplitude of potassium currents in IHCs showed no temperature dependence either in outward-going IK,f, or inward-going IK,n, however, activation rates became faster at 36℃ than at room temperature (Fig. 4). Inactivation, that was characteristically possessed in TEA-sensitive IK,f, was much faster at 36℃ than at room temperature (Fig. 5). Q10 of activation and inactivation was 1.83 and 3.19, respectively.
A temperature dependency of potassium currents has been described in several cells. In guinea pig ventricular cells, the conductance of inward rectifier potassium currents decreased by lowering the temperature with a Q10 of 1.28 (whole cell recordings), or 1.41 (single channel recordings) between 20℃ and 30℃ [13]. The channel open probability was scarcely affected by temperature, however, the time constant of open-time histogram progressively increased as the temperature was lowered and the configuration of closed-time distribution was markedly modified by changing the temperature. These findings strongly suggested the kinetics of potassium channels were dependent on temperature. In rat skeletal muscle (omohyoid muscle), potassium currents were measured from 1℃ to 37℃ using the three-microelectrode voltage-clamp technique [14]. The effect of temperature on the activation time constant (τn) was much greater in the cold than in the warm: τn had a Q10 of nearly 6 at temperatures below 10℃, but a Q10 of only ~2 over the range of 30℃-38℃. Voltage-gated potassium channels in human lymphocytes, which are similar to IK,f in IHCs for the sensitivity of TEA, were studied at temperatures from 5 to 42℃ [15]. Heating the cell increased the current amplitude at any given membrane potentials between -30 and 30 mV. As for conductance, the channel activation rate was increased by warming the cell and the inactivation rate also progressively increased with increasing temperature. However, in contrast with conductance and activation rates, inactivation rates were most sensitive to temperature changes above room temperature (there was no inactivation below 20℃).
In IHCs, a temperature enhancement of presynaptic function was reported [6]. Readily releasable pool exocytosis (within 20 ms) was increased at physiological temperature with a Q10 of 2.1. In spite of the temperature enhancement of fast exocytosis, sustained exocytosis (between 20 and 100 ms) was not significantly different between room and body temperature. Voltage-gated L-type Ca2+ channels trigger the release of synaptic vesicles at IHC active zones [16]. An increase in temperature increased the L-type Ca2+ current amplitude and accelerated activation kinetics [6]. Q10 of amplitude is 1.36 and 1.90 in adult IHCs and immature IHCs, respectively [17]. They assumed that since open channel probability of Ca2+ channels become less temperature dependent with maturation, Q10 is smaller in adult IHCs than in immature IHCs. Lowering the temperature from ~37℃ to room temperature slowed the activation time constant of Ca2+ currents by about half [6], suggesting that Q10 is around 1.8.
Outward potassium currents in IHCs, such as IK,f, are expressed during development of auditory responsiveness [18], and allow the cells to perform high frequency transduction by greatly shortening the membrane time constant [1]. Accelerating potassium current activation by temperature can lead to the faster response of IHCs' exocytosis mentioned above. Fast potassium currents were about 5 times slower at room temperature than at body temperature and showed a clear delay at onset [1]. Q10 for the kinetics of the fast current was estimated as 3.0 on the basis of tail current relaxations. In the present study, Q10 was larger for inactivation (3.19) than for activation (1.83). Inactivation of potassium currents are considered to be conferred by many treatments and cytosolic domains, such as intracellular Ca2+ [19], cyclic adenosine monophosphate [20], papain [21], trypsin [22], and 'ball' peptide [21]. Intracellular agents are predominantly entropy driven, so temperature dependency for the inactivation process may be greater than for the activation process.
Go to :
 XML Download
XML Download