

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Mucins are a family of macromolecular glycoproteins that are synthesized by the epithelial cells lining the respiratory tract. Mucins are involved in normal mucociliary clearance and contribute to the innate immune system [1, 2]. There are two types of mucins, one which is secreted and the other which is membrane-bound mucins. In the respiratory tract, MUC5AC and MUC5B are the main secreted mucins that account for approximately 90% of the mucin content of sputum; MUC5AC is produced by goblet cells, and MUC5B is secreted from the mucous cells in the submucosal glands [1, 3-6]. MUC1, MUC4, and MUC16 are the predominant membrane-bound mucins that are included in the remaining 10% of the mucin content of sputum. They are expressed on the apical surface of epithelial cells including the mammary glands, lacrimal glands, salivary glands, gastrointestinal tract, respiratory tract and genitourinary tract [4, 7, 8].
MUC16 is one of 10 membrane-bound mucins: MUC1, MUC3A, MUC3B, MUC4, MUC12, MUC13, MUC15, MUC16, MUC17, and MUC20 [7, 9, 10]. Its gene is located on chromosome 19p13.2. It is a large protein, with a core protein of 22,152 amino acids, a molecular mass of 2.5 MDa, and a potential glycosylated mass of approximately 20 MDa. MUC16 is the transmembrane counterpart to the well-known serum marker of ovarian cancer [5, 11]. However, it is also expressed on the apical surface of noncancerous native epithelia including the ocular surface and female reproductive tract [7, 12-14]. In 2007, MUC16 was first reported to be produced by normal human airway epithelial cells [8]. Although the biological role of MUC16, on the apical surface of noncancerous native epithelia, is not fully known, several studies have shown that MUC16 is associated with the maintenance of hydration and lubrication, the formation of a disadhesive barrier and the regulation of the innate mucosal defense of epithelium [7, 13-17].
Phorbol 12-myristate 13-acetate (PMA) is widely used as a protein kinase C (PKC) activator; PKC is involved in the secretion of mucins [18, 19]. PMA has been reported to up-regulate MUC2, MUC5AC and MUC5B via mitogen-activated protein kinase (MAPK) signaling pathway as well as cytokines and bacterial lipopolysaccharides [18, 20-22]. While a study has reported that PMA increased MUC16 expression in human nasal mucosa [4], the effect and signaling pathway of PMA on MUC16, in human airway epithelial cells, has not been reported in the English-language literature.
Therefore, the goal of this study was to determine the role of PMA on the regulation of MUC16 expression through PKC-MAPK signaling pathway in human airway epithelial cells.
MATERIALS AND METHODS
Cell culture and treatment
Inferior turbinate mucosa obtained from patients undergoing augmentation rhinoplasty was used as reference tissue for MUC16 expression after the approval of the Institutional Review Board for human studies at the Yeungnam University Medical Center. For the primary culture of the normal nasal epithelial cells, the inferior turbinate mucosa was washed with phosphate-buffered saline (PBS) and immersed in dispase (Boehringer Mannheim, Mannheim, Germany) for 90 minutes. After the tissue was scraped off the surface of the inferior turbinate mucosa with a scalpel, it was added to 1% PBS and filtered through a mesh. The cells were seeded in a 24-well plate at 2.5×105 cells/well and were then incubated with EpiLife Medium (Cascade Biologics, Portland, OR, USA) and Human Keratinocyte Growth Supplement (5/500 mL of medium, Cascade Biologics). When the cultures were confluent, the primary cultures of normal nasal epithelial cells were exposed to the indicated concentrations of PMA (Sigma, St. Louis, MO, USA).
The mucin-producing human NCI-H292 airway epithelial cells (American Type Culture Collection, Manassas, VA, USA) were cultured in RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 2 mM L-glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin and 10% fetal bovine serum (FBS). The cells were grown at 37℃ in 5% CO2 fully humidified air and were subcultured twice weekly. When confluent, the cells were incubated in RPMI 1640 medium containing 0.5% FBS for 24 hours. The cells were then rinsed with serum-free RPMI 1640 medium and exposed to the indicated concentrations of exposed to the indicated concentrations of PMA.
In order to investigate the signaling pathway of PMA-induced MUC16 expression, the cultured cell were pretreated with specific inhibitors, Calphostin C (Sigma), Rottlerin (Sigma), U0126 (Calbiochem, San Diego, CA, USA), and/or SB2035 (BOMOL Research Laboratories, Plymouth Meeting, PA, USA), for 1 hour before exposure to the indicated concentrations of PMA. For the controls, the cultured cells were incubated with medium alone for the same amount of time.
Reverse transcription-polymerase chain reaction (RT-PCR) analysis of MUC16 mRNA expression
Total RNA was isolated from the cultured cells according to the manufacturer's instructions (Applied Biosystems, Branchburg, NJ, USA). Each sample was reverse transcribed into cDNA using GeneAmp RNA PCR Core Kit (Applied Biosystems). Briefly, The PCR conditions used to amplify the MUC16 were 30 cycles of denaturation (at 95℃/minute), annealing (at 60℃/minute), and extension (at 72℃/minute) in the presence of 2.5 mg MgCl2, followed by a final 20 minutes extension step at 72℃. The PCR of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was performed on each individual sample as a positive internal control. The primer sequences used in the PCR are shown in Table 1. The relative intensity of individual bands on the gel image was determined as the ratio of intensity of MUC16 to the intensity of GAPDH.
Real-time PCR analysis of MUC16 mRNA expression
Real-time PCR was performed using LC Fast Start DNA Master SYBR Green kit (Roche Applied Science, Mannheim, Germany) with 0.5 µL of cDNA, corresponding to 25 ng of total RNA in a 10 µL final volume, 2.5 mM MgCl2 and 0.5 µM of each primer (final concentration). Quantitative PCR was performed using LightCycler (Roche Applied Science) for 45 cycles at 95℃ for 10 seconds, specific annealing temperature for 5 seconds and 72℃ for 10 seconds. Data were normalized to GAPDH. Amplification specificity was evaluated using melting curve, following the manufacturer's instructions (Roche Applied Science).
Enzyme-linked immunosorbent assay (ELISA) analysis of MUC16 protein
MUC16 protein level was determined by ELISA. Each sample of the supernatant or cell lysates from the NCI-H292 airway epithelial cells was incubated at 40℃ in a 96-well plate. The plates were blocked with 2% bovine serum albumin, and incubated with primary antibody of MUC16 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) diluted with PBS containing 0.05% Tween 20 for 1 hour. The wells were treated with horseradish peroxidase (HRP)-conjugated secondary antibody of MUC16 (Santa Cruz Biotechnology). After 4 hours, color was developed using 3,3',5,5'-tetramethylbenzidine peroxidase solution and stopped with 2N-H2SO4. Optical density measurements were obtained using ELISA reader (EL800; BIO-TEK Instruments, Winooski, VT, USA) at 450 nm.
Western blot analysis of extracellular signal-regulated kinase 1/2 (ERK1/2) and p38 MAPK phosphorylation
The NCI-H292 airway epithelial cells were seeded in a 6-well plate and treated with PMA, and/or Calphostin C and Rottlerin for the indicated times. The cells were exposed to trypsin, and formed into pellets at 700 g at 4℃. The pellets were resuspended in lysis buffer. The preparation was then clarified by centrifugation, and the supernatant was saved as a whole-cell lysate. The proteins (50 µg) were separated using 10% reducing sodium dodecyl sulfate-polyacrylamide gel electrophoresis and electroblotted onto a nitrocellulose membrane. The membrane was then blocked with 5% non-fat dry milk, followed by incubation with the indicated primary antibody of ERK1/2 (Santa Cruz Biotechnology) or p38 MAPK (Santa Cruz Biotechnology) for 4 hours. Subsequently, the membrane was washed and incubated for 1 hour with secondary antibody of ERK1/2 or p38 MAPK conjugated to HRP, and developed using enhanced chemiluminescence reagent kit (PerkinElmer Life Sciences, Boston, MA, USA) and exposed to X-ray film for 10 seconds.
Cell transfection with small interfering RNA (siRNA) for p38 MAPK
Predesigned siRNA targeting p38 MAPK and negative control siRNA for p38 MAPK were purchased from Invitrogen Co. Sequences of p38 MAPK siRNA are as follows: sense, AUG AAU GAU GGA CUG AAA UGG UCU G and antisense, CAG ACC AUU UCA GUC CAU CAU UCA U. The transfection rate of p38 MAPK siRNA was verified to be over 90% in human NCI-H292 airway epithelial cells. Transfection was performed according to the manufacture's protocol (Invitrogen). The same procedure was performed with negative control siRNA.
Statistical analysis
Statistical analysis was performed using SPSS ver. 10.0 (SPSS Inc., Chicago, IL, USA). The mean for each of the obtained quantitative values was calculated. Comparisons were made using Student's t-test. For all tests, a P-value of less than 0.05 was considered statistically significant.
RESULTS
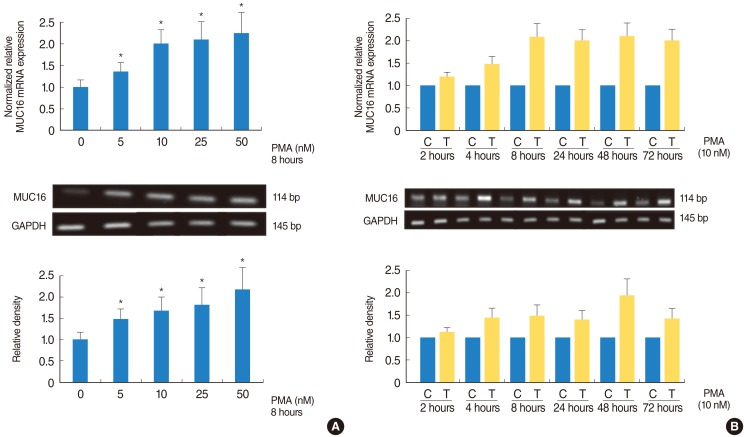
PMA induced MUC16 expression in the NCI-H292 airway epithelial cells
To investigate the effect of PMA on MUC16 expression in a dose- and time-dependent manner, real-time PCR, RT-PCR, and ELISA analysis were performed in the NCI-H292 airway epithelial cells. PMA significantly induced MUC16 expression in a dose-dependent manner (P<0.05) (Fig. 1A). However, PMA did not induce MUC16 expression in a time-dependent manner (Fig. 1B).
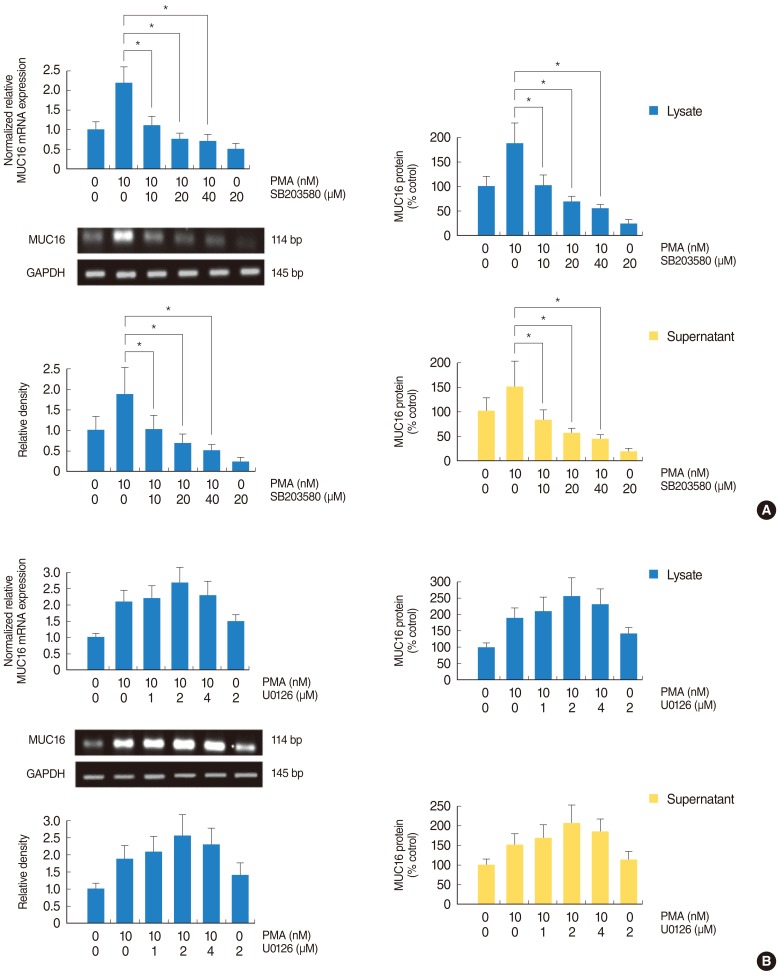
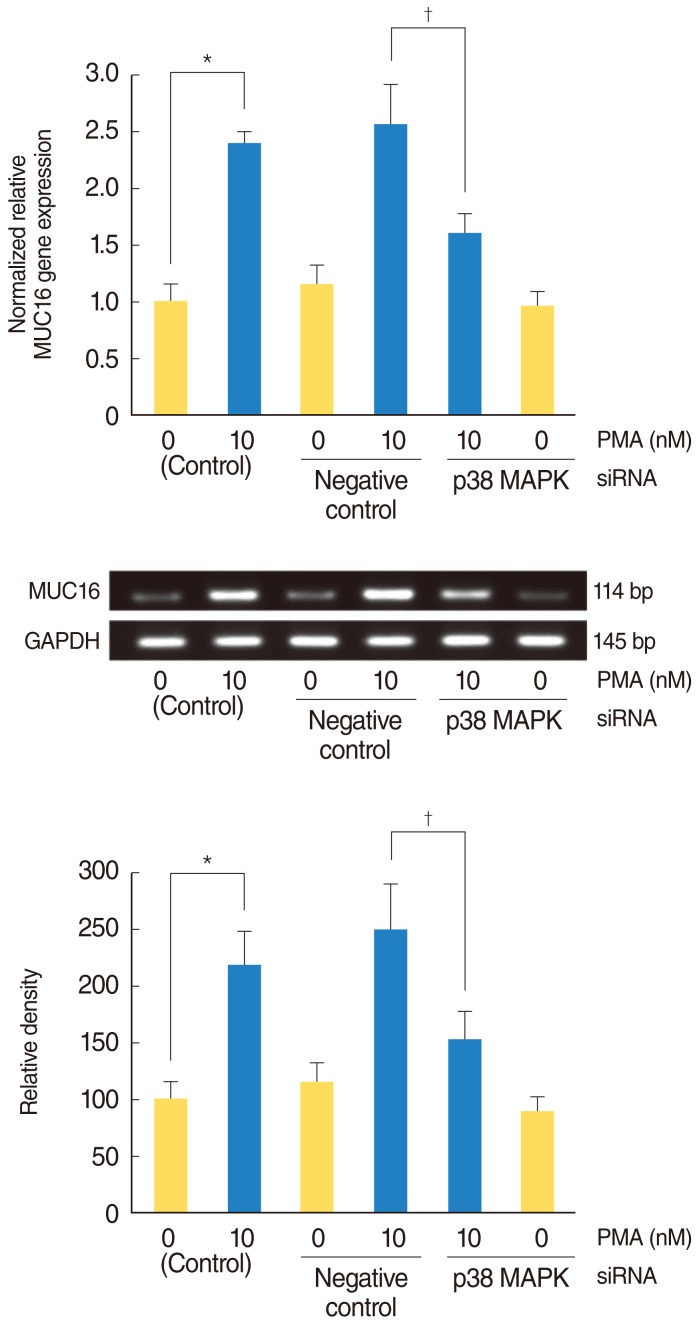
PMA activated phosphorylation of p38 MAPK associated with MUC16 expression in the NCI-H292 airway epithelial cells
To investigate the brief intracellular mechanisms of PMA-induced MUC16 expression in the NCI-H292 airway epithelial cells, Western blot analysis was performed to evaluate the phosphorylation of ERK1/2 and p38 MAPK. PMA activated the phosphorylation of p38 MAPK, but it did not activate the phosphorylation of ERK1/2 (Fig. 2). To confirm the phosphorylation of p38 MAPK, real-time PCR, RT-PCR, ELISA and cell transfection with siRNA analysis were performed to investigate the effect of MAPKs inhibitors on PMA-induced MUC16 expression. SB203580 inhibited the PMA-induced MUC16 expression (P<0.05) (Fig. 3A), while U0126 did not (Fig. 3B). In addition, the knockdown of p38 MAPK by p38 MAPK siRNA significantly blocked PMA-induced MUC16 mRNA expression (P<0.05) (Fig. 4).
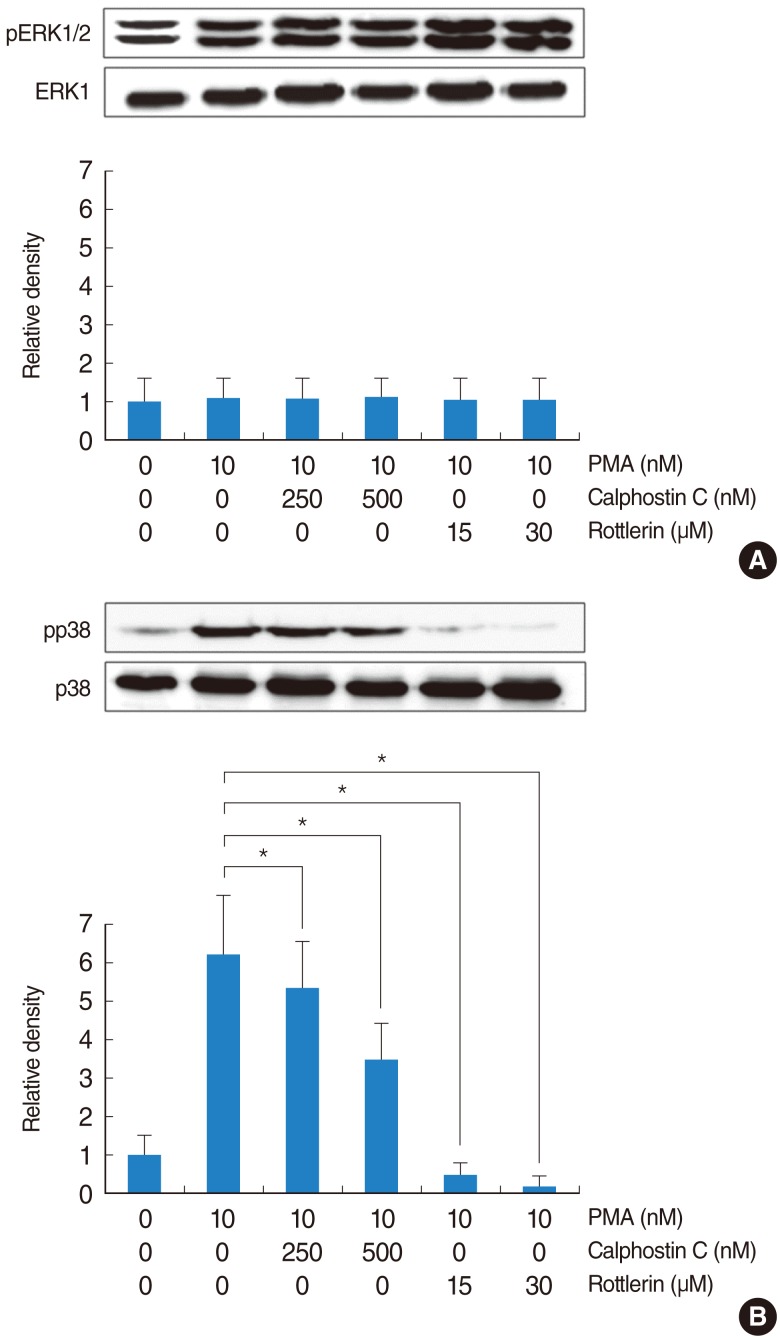
PKCδ mediated PMA-induced MUC16 expression in the NCI-H292 airway epithelial cells
Real-time PCR, RT-PCR, and ELISA analysis were performed to investigate the role of PKC on PMA-induced MUC16 expression in the NCI-H292 airway epithelial cells. Calphostin C and Rottlerin inhibited PMA-induced MUC16 expression (P<0.05) (Fig. 5). In addition, the involvement of PKC in the phosphorylation of MAPKs after treatment with PMA was investigated as well. The maximum phosphorylation levels of ERK1/2 and p38 MAPK were observed 25 minutes after initiation of treatment. While the phosphorylation of ERK1/2 was not inhibited by Calphostin C and Rottlerin after treatment with PMA (Fig. 6A), the phosphorylation of p38 MAPK was inhibited by Calphostin C and Rottlerin after treatment with PMA (Fig. 6B). In addition, the inhibiting effect of Rottlerin was significantly stronger than Calphostin C (P<0.05).
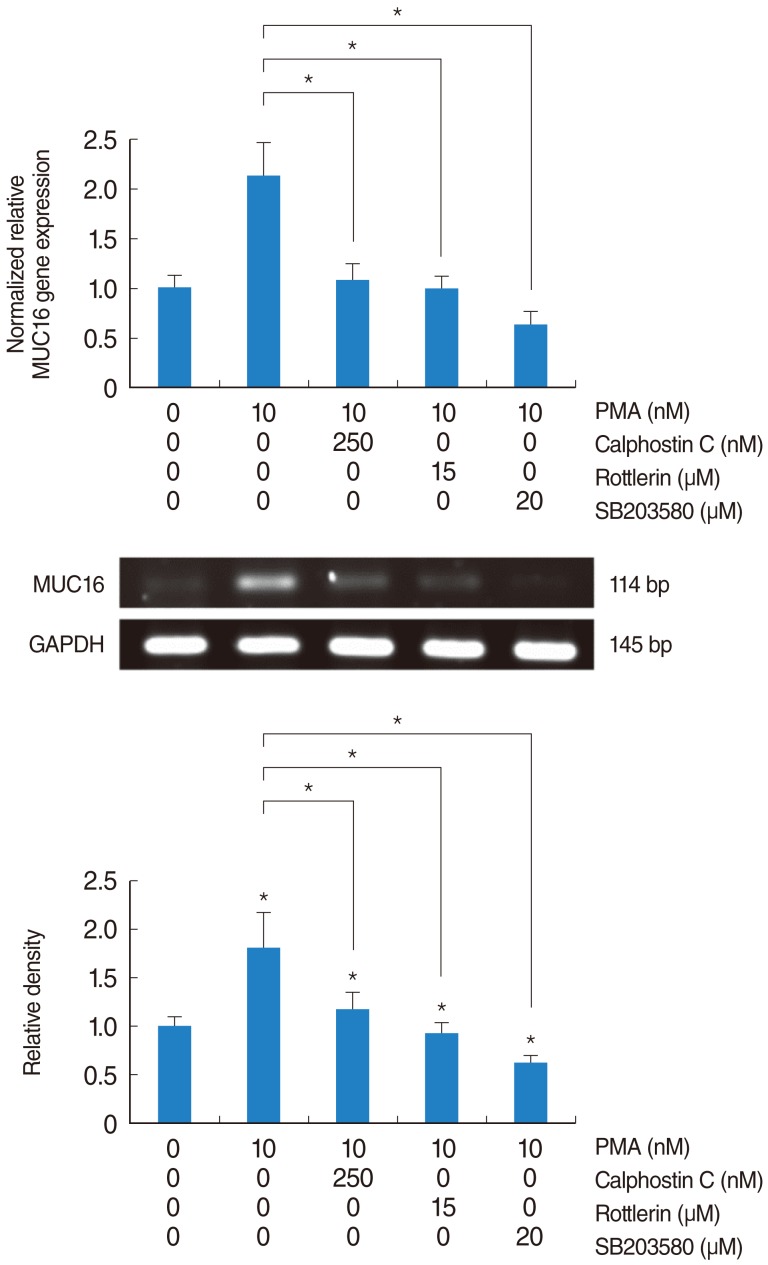
PKCδ and p38 MAPK associated with PMA-induced MUC16 expression in the primary cultures of normal nasal epithelial cells
Real-time PCR and RT-PCR analysis were performed to investigate PKCδ and p38 MAPK in associated with PMA-induced MUC16 expression in the primary cultures of normal nasal epithelial cells. Calphostin C, Rottlerin and SB203580 inhibited PMA-induced MUC16 expression (P<0.05) (Fig. 7). In addition, the inhibiting effect of Rottlerin was significantly stronger than Calphostin C (P<0.05).
DISCUSSION
MUC16, as a multifunctional molecule, has the classical mucin-associated functions of maintenance of hydration and lubrication of the epithelial surface, and the formation of a disadhesive barrier. And MUC16 is also considered to be a signaling molecule which influences the transcription of genes involved in cell proliferation and differentiation [14]. MUC16 was reported to be produced by human tracheal surface epithelium and bronchial epithelial cells after treatment with bacterial exoproducts [8, 15]. These previous studies suggest that MUC16 may play an important role in the defense against bacteria, and may be induced by a variety of inflammatory mediators such as bacterial lipopolysaccharide, interleukin-1β (IL-1β) and PMA in the inflammatory airway diseases. Of particular interest, a recent study has reported that MUC16 differed from MUC4 in the regulation of expression in human nasal polyp epithelial cells; unlike MUC4, PMA increased MUC16 expression, while IL-1β had no effect [4].
Among the inflammatory mediators, PMA can substitute for diacylglycerol, and is known to activate PKC, which in turn initiates signaling cascades that lead to a variety of cell events [18-20]. In the induction of mucins, several studies have also reported that PMA stimulates the expression of MUC5AC and MUC5B in the airway and intestinal epithelial cells [18, 21, 22]. However, unlike MUC5AC and MUC5B, there are few or no reports on the effect of PMA on MUC16 expression [4]. In this study, PMA was found to induce MUC16 expression in a dose-dependent manner. This result suggests that MUC16 is produced in human airway epithelial cells and PMA up-regulates MUC16 as well as MUC5AC and MUC5B in human airway epithelial cells.
Several studies have reported on the signaling pathways of PMA-induced MUC5AC and MUC5B expression. PMA induced MUC5AC expression via the epidermal growth factor receptor (EGFR), Ras, Raf, ERK, and Sp1 signaling pathway in the airway epithelial cells [18, 21, 22]. However, the signaling pathway of PMA-induced MUC5B was PKCδ-, Ras-, MAPK kinase kinase1-, and JNK/p38 MAPK-dependent pathway but EGFR/ERK-dependent pathway in the airway epithelial cells [21, 22]. Among the factors involved in the signaling pathways of PMA-induced MUC5AC and MUC5B expression in the airway epithelial cells, PKCδ as a novel PKC isoform, was activated by phosphatidylserine, diacylglycerol or PMA. PKCδ is involved in regulating the expression of inducible nitric oxide synthase, granulocyte colony stimulating factor, and intercellular adhesion molecule-1 [19]. PKCδ is also reported to regulate mucin production and secretion; the PKCδ signaling pathway is considered as a pathway by which PMA induces MUC5B expression [1, 19, 22, 23]. The p38 MAPK is responsive to stress stimuli, and is involved in cell differentiation and apoptosis. In addition, p38 MAPK is also involved in regulating the expression of the mucin gene, and its signaling pathway is considered a kind of pathway by which PMA induces MUC5B expression [1, 21, 22]. However, the signaling pathway associated with PMA-induced MUC16 expression has not yet been reported. Therefore, in this study, PMA-induced MUC16 expression through the PKC-MAPK signaling pathway was investigated; this investigation focused on PKCδ and p38 MAPK in the signaling pathway. PMA was found to activate phosphorylation of p38 MAPK. Rottlerin (PKCδ inhibitor) inhibited PMA-induced MUC16 expression, and also inhibited the phosphorylation of activated p38 MAPK by PMA. SB203580 (p38 MAPK inhibitor) inhibited PMA-induced MUC16 expression. In addition, a knockdown of p38 MAPK by p38 MAPK siRNA significantly blocked PMA-induced MUC16 mRNA expression. Although the effect of PMA on EGFR, Raf, Ras, and JNK signaling pathways was not investigated in the present study, the results of this study suggest that the signaling pathway associated with PMA-induced MUC16 are PKCδ and p38 MAPK in the airway epithelial cells. However, the precise signaling pathways of PMA-induced MUC16 expression in the airway epithelial cells require further confirmation.
In conclusion, the results of this study show for the first time that PMA-induced MUC16 expression appears to be regulated by activation of PKCδ and p38 MAPK signaling pathway in human airway epithelial cells. These findings provide important information about the biological role of MUC16 and the intracellular mechanisms of MUC16 expression in airway epithelial cells.



 XML Download
XML Download