

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Endoplasmic reticulum (ER) stress response is an important phenomenon observed in many inflammatory diseases, and results in the activation of unfolded protein response [1,2]. Moreover, ER stress response has been implicated in the pathogeneses of various inflammatory airway diseases, including viral and bacterial infections, cystic fibrosis, asthma, and chronic obstructive pulmonary disease [2-4]. Eukaryotic cells exhibit three major ER stress pathways, that is the double-stranded RNA-activated protein kinase-like ER kinase (PERK), inositol requiring enzyme 1 (IRE1) involved in X-box binding protein 1 (XBP-1), and the activating transcription factor 6 (ATF6) pathways. Furthermore, these three major pathways function differently in ways determined by the pathogeneses of airway diseases [3-7].
The surfaces of the human nasal mucosa are continuously exposed to particulates and pathogens. Human nasal airway epithelial cells secrete mucus which serves as the first line of defense against inhaled pathogens as protective mucus barriers trap and allow the elimination of pathogens via mucociliary clearance [8-10]. However, excessive mucus is major symptom, which is associated with goblet cell metaplasia and hypertrophy of nasal airway epithelium in chronic rhinosinusitis (CRS). Mucins are high molecular weight glycoproteins, one of major component responsible for mucus secretion, impart different physical and biological properties to mucus of the airway [11,12]. Among the airway mucins, the levels of MUC5B and MUC5AC mRNA in CRS were significantly increased compared with those in normal sinus mucosa. This change of nasal mucin gene expression by sinus epithelium as a consequence of sinus disease will almost certainly affect the composition and biophysical properties of sinus mucus [8]. Nevertheless, the mechanisms underlying overproduction of MUC5B and MUC5AC glycoprotein in CRS are not fully understood.
Accordingly, our aims were to determine whether ER stress affects regulation of the expressions of MUC5AC and MUC5B and to investigate the signaling pathways responsible for the ER stress-induced expressions of MUC5AC and MUC5B in human nasal airway epithelial cells.
Go to : 
MATERIALS AND METHODS
Materials
Tunicamycin (an ER stress inducer) and 4-phenylbutyric acid (4-PBA, an ER stress inhibitor) were obtained from Sigma-Aldrich (St. Louis, MO, USA). Primary human nasal epithelial cells (HNEpCs) and airway epithelial cell growth medium were purchased from PromoCell (Heidelberg, Germany). RPMI 1640 medium and Trizol were from Invitrogen (Carlsbad, CA, USA). EZ-Cytox Cell Viability Assay kits were purchased from Daeil Laboratories (Seoul, Korea). Reverse transcriptase-polymerase chain reaction (RT-PCR) kits were from Applied Biosystems (Foster City, CA, USA). Real-time PCR kits were obtained from Roche Applied Science (Mannheim, Germany). Primary and secondary antibodies for MUC5AC and MUC5B (used immunoassay) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). For Western blot analysis, primary and secondary antibodies of XBP-1, CCAAT-enhancer-binding protein homologous protein (CHOP), ATF6 and β-actin were purchased from Abcam (Cambridge, MA, USA). Fetal bovine serum (FBS) was purchased from Hyclone Laboratories (Logan, UT, USA). OPTI-MEM I Reduced Serum Medium, Lipofectamine 2000, predesigned small interfering RNAs (siRNAs) targeting XBP-1, CHOP, and ATF6, and control siRNA were purchased from Thermo Fisher Scientific (Lafayette, CO, USA). This study was approved by the Institutional Review Board of Yeungnam University Medical Center (IRB No. YUMC 2015-06-058).
Cell culture and treatment
HNEpCs were cultured in Airway Epithelial Cell Growth Medium at 37˚C in 5% CO2 fully humidified air and subcultured according to the recommendations of PromoCell. Passage-2 HNEpCs (1×105 cell/culture) were seeded in 0.5 mL of culture medium. When the cultures were confluent, the cells were exposed to the indicated concentrations of tunicamycin or 4-PBA. The proliferations and viabilities of HNEpCs were quantified using an EZ-Cytox Cell Viability Assay kit, and changes in cell morphology were documented using a microscope.
RT-PCR of the mRNA expressions of MUC5AC, MUC5B, spliced XBP-1, CHOP, and ATF6
Total RNA was isolated from cultured cells using RT-PCR kits (Applied Biosystems). Samples were reverse transcribed into complementary deoxyribonucleic acid (cDNA) using the GeneAmp RNA PCR core kit (Applied Biosystems). The primer sequences and conditions used for MUC5AC, MUC5B, spliced XBP-1, CHOP, and ATF6 were as previously described protocols (Table 1) [6,13,14]. RNA integrities and the success of RT reactions were monitored by PCR amplification of transcripts using of the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene. PCR products were electrophoresed in 2% agarose gels, and visualized by staining with ethidium bromide and ultraviolet fluorescence. RT-PCR products were semiquantitatively analyzed using scanned gel images and by measuring the intensities of PCR products using commercially available imaging software (Scion, Frederick, MD, USA). Relative intensities were calculated by expressing the intensities of MUC5AC, MUC5B, spliced XBP-1, CHOP, and ATF6 as ratios of GAPDH intensities.
Table 1.
Primer sequences, lengths of amplified templates, and primer-specific conditions for RT-PCR
![]()
Real-time PCR for the mRNA expressions of MUC5AC, MUC5B, spliced XBP-1, CHOP, and ATF6
Real-time PCR was performed using the LightCycler FastStart DNA Master SYBR Green kit (Roche Applied Science) and 0.5 μL of cDNA (corresponding to 25 ng of total RNA) in a final volume of 10 μL of 2.5 mM MgCl2 and 0.5 μM of each primer (final concentration). Quantitative PCR was performed using a LightCycler using 45 cycles of 95˚C for 10 seconds, a specific annealing temperature for 5 seconds, and 72˚C for 10 seconds. Data were normalized versus GAPDH. Melting curves were used to evaluate amplification specificity, according to the manufacturer’s instructions (Roche Applied Science).
Enzyme-linked immunosorbent assay analysis of MUC5AC and MUC5B glycoproteins
The protein levels of MUC5AC and MUC5B were determined by enzyme-linked immunosorbent assay (ELISA). Samples of cell supernatants were prepared in phosphate-buffered saline (PBS) at several dilutions, incubated at 40˚C in a 96-well plate until dry, washed three times with PBS, blocked with 2% FBS for 1 hour at room temperature, rewashed three times with PBS, and incubated with primary antibody for MUC5AC or MUC5B (diluted at 1:200 with PBS containing 0.05% Tween 20) for 1 hour. Wells were then washed three times with PBS, and horseradish peroxidase-conjugated secondary antibody for MUC5AC or MUC5B was added to wells, and allowed to stand for 4 hours. Color was developed using 3,3´,5,5´-tetramethylbenzidine peroxidase solution and stopped with 2N-H2SO4. Optical densities were measured using an ELISA reader (EL800; BIO-TEK Instruments, Winooski, VT, USA) at 450 nm. Results are expressed as percentages of baseline controls.
Western blot analysis for XBP-1, CHOP, and ATF6
HNEpCs were seeded in each well of a 6-well plate, and treated with the indicated concentrations of tunicamycin. The cells harvested 200 μL radioimmunoprecipitation assay buffer with phosphatase inhibitor cocktail and incubated for 20 minutes at 4˚C. And then sample were centrifuged at 2,500×g for 10 minutes at 4˚C and the supernatant (whole-cell lysate) was collected. Proteins (20 μg) were separated using 10% reducing sodium dodecyl sulfate-polyacrylamide gel electrophoresis and electroblotted onto a polyvinylidene difluoride membrane. The membrane was blocked 5% bovine serum albumin in tris buffered saline with 0.1% Tween 20, followed by incubation with the indicated primary antibody of XBP-1 (ab37152), CHOP (ab11419) and ATF6 (ab37149) for 4 hours. Subsequently, the membrane was incubated for 1 hour with secondary antibody of XBP-1, CHOP and ATF6 conjugated to horseradish peroxidase, and developed using an enhanced chemiluminescence system kit. Signals corresponding to the protein of interest were developed using an enhanced West Pico Chemiluminescent Substrate (Thermo Fisher, Boston, MA, USA). Bands were detected after exposure of the membrane to an imaging system (Chemiluminescence image system, FUSION-FX7 820wL, Vilber, Eberhardzell, Germany).
Cell transfection with siRNAs for XBP-1, CHOP, or ATF6
The transfection rate of each siRNA was confirmed to be ≥90%. Transfection was performed according to the manufacturer’s instructions (Thermo Fisher Scientific); XBP-1 siRNA (s14913), CHOP (VHS40605), and ATF6 (s223543). In brief, HNEpCs were seeded in the wells of 6-well plates at 1×105 cells/well and incubated overnight in RPMI 1640 medium. When cells were 80%–90% confluent, the following day, they were washed with PBS and OPTI-MEM I Reduced Serum Medium was added. Separately, XBP-1 siRNA and Lipofectamine 2000 were incubated together in OPTI-MEM I Reduced Serum Medium to form a XBP-1 siRNA-Lipofectamine complex, and this complex-containing medium was then added to wells containing the cells, which were then incubated for 24 hours at 37˚C in a CO2 incubator. The XBP-1 siRNA-Lipofectamine complex-containing medium was then replaced with RPMI 1640 medium after 4 hours without loss of transfection activity. After 24 hours of transfection with XBP-1 siRNA, cells were exposed to tunicamycin (0.5 μg/mL) and then harvested for RT-PCR analysis. The transfection rate of XBP-1 siRNA was confirmed to be ≥90%. The same procedure was used to transfect cells with the siRNAs for CHOP or ATF6.
Statistical analysis
Statistical analysis was performed using the SPSS ver. 12.0 (SPSS Inc., Chicago, IL, USA). The mean and standard deviation were calculated for sets of measurements. Data comparison was done with the unpaired t-test or Kruskal-Wallis one-way analysis of variance followed by the Mann-Whitney test. For all tests, P-value <0.05 was considered statistically significant.
Go to :
RESULTS
ER stress increased the expressions of MUC5AC and MUC5B in HNEpCs
HNEpCs were incubated with tunicamycin (0, 0.25, 0.5, 1, or 2.5 μg/mL) for 8 hours and then RT-PCR and real-time PCR were performed to determine the expressions of MUC5AC and MUC5B. The results obtained showed that tunicamycin significantly induced the mRNA expression of MUC5AC and MUC5B (Fig. 1A, B). To determine changes in the mRNA expressions of MCU5AC and MUC5B after different exposure times, HNEpCs were treated with tunicamycin (0.5 μg/mL) for various times (0, 1, 2, 4, 8, or 24 hours). Real-time PCR showed that tunicamycin induced the expressions of MUC5AC and MUC5B at all times, but peak expression times differed; the mRNA expression level of MUC5AC peaked after 4 hours of exposure to tunicamycin, whereas the mRNA expression level of MUC5B peaked at 8 hours (Fig. 1C). ELISA also showed tunicamycin induced the productions of MUC5AC and MUC5B glycoproteins (Fig. 1D).
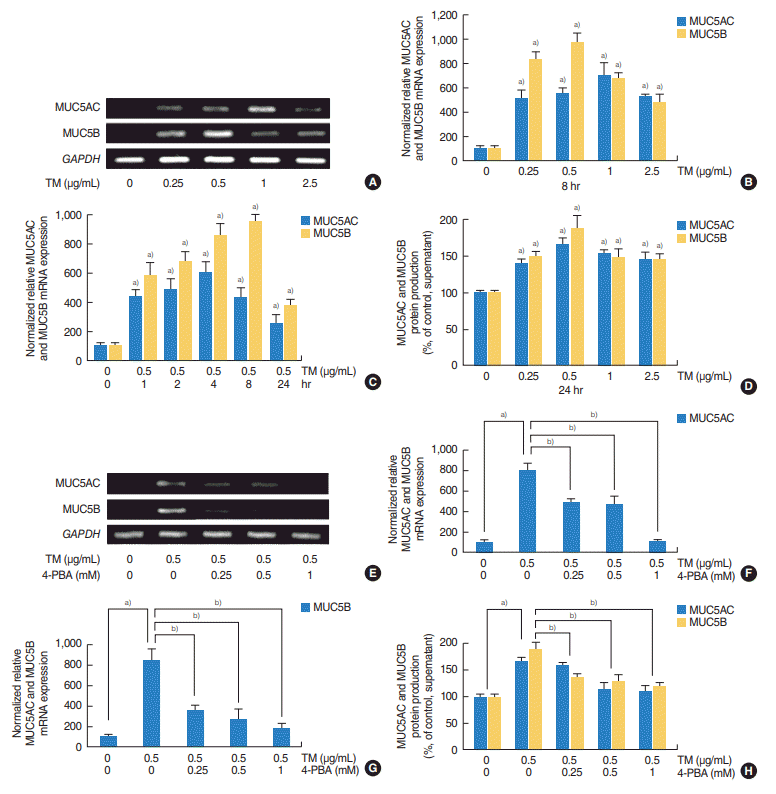 | Fig. 1.Effects of tunicamycin (TM; an endoplasmic reticulum stress inducer) on the expressions of MUC5AC and MUC5B in human nasal epithelial cells. (A, B) Reverse transcriptase-polymerase chain reaction (RT-PCR) and real-time PCR showed that TM significantly increased the mRNA expressions of MUC5AC and MUC5B at all concentrations examined. (C) Time-dependent changes, real-time PCR showed that TM induced the expressions of MUC5AC and MUC5B mRNA at all times. However, peak times differed; MUC5AC peaked after 4 hours of exposure to TM, whereas MUC5B peaked after 8 hours of exposure. (D) Enzyme-linked immunosorbent assay (ELISA) showed TM induced the productions of MUC5AC and MUC5B glycoproteins. Effects of 4-phenylbutyric acid (4-PBA; an endoplasmic reticulum stress inhibitor) on the TM-induced expressions of MUC5AC and MUC5B in human nasal epithelial cells. (E-G) RT-PCR and real-time PCR showed that 4-PBA significantly attenuated the TM-induced expressions of MUC5AC and MUC5B mRNA. (H) ELISA also showed that 4-PBA significantly attenuated TM-induced the productions of MUC5AC and MUC5B glycoprotein. The images shown are representative of three separate experiments performed in triplicate. Bars represent the averages±standard deviation of three independent experiments performed in triplicate. GAPDH, glyceraldehyde-3-phosphate dehydrogenase. a)P<0.05 vs. baseline. b)P<0.05 vs. TM (0.5 µg/mL) alone.
|
4-PBA reduced tunicamycin-induced MUC5AC and MUC5B expressions in HNEpCs
To confirm the effect of tunicamycin (an ER stress inducer) on MUC5AC and MUC5B expressions. HNEpCs pre-exposed to tunicamycin (0.5 μg/mL) were treated with different doses of 4-PBA (0, 0.25, 0.5, 1 mM); an ER stress inhibitor. RT-PCR and real-time PCR showed that 4-PBA significantly attenuated the tunicamycin-induced expressions of MUC5AC and MUC5B mRNA (Fig. 1E-G), ELISA also showed that 4-PBA significantly attenuated the tunicamycin-induced productions of MUC5AC and MUC5B glycoproteins (Fig. 1H).
The three major signaling mechanisms were involved in the ER stress-induced expressions of MUC5AC and MUC5B in HNEpCs
To investigate the roles of three major signaling pathways, that is, the IRE1/XBP-1, PERK/ATF4/CHOP, and the ATF6 pathways, on the expressions of MUC5AC and MUC5B in HNEpCs, cells were stimulated with different doses of tunicamycin with/without 4-PMA for 1 hour, and the mRNA expressions of spliced XBP-1, CHOP, and ATF6 were then checked. RT-PCR and real-time PCR showed that their mRNA expressions were significantly increased at all tunicamycin doses (0.25, 0.5, 1, 2.5 μg/mL) (Fig. 2A, B) and attenuated by 4-PBA. (Fig. 2C, D). Western blot was also performed to check the levels of XBP-1, CHOP, and ATF6. The results showed that tunicamycin induced XBP-1, CHOP and ATF6 and attenuated by 4-PBA (Fig. 2E-G). In addition, transfection with siRNAs for XBP-1, CHOP, and ATF6 were performed to evaluate individual pathway involvements. RT-PCR, real-time PCR and ELISA were used to assess the contributions of the three major mechanisms to the ER stress-mediated expressions of MUC5AC and MUC5B. RT-PCR, real-time PCR and ELISA showed that siRNA knockdowns of XBP-1, CHOP, or ATF6 significantly blocked ER stress-induced MUC5AC mRNA expression and glycoprotein production (Fig. 3A-I) although inhibition by XBP-1 siRNA was greatest (Fig. 3A-C), and inhibition by CHOP siRNA was weakest (Fig. 3D-F). In the study on MUC5B expression, all three siRNAs significantly blocked ER stress-induced MUC5B mRNA expression and glycoprotein production (Fig. 4A-I). However, the inhibitory pattern differed from that of MUC5AC. In this case, inhibition by CHOP siRNA was the greatest (Fig. 4D-F), and inhibitions by XBP-1 was weakest (Fig. 4A-C).
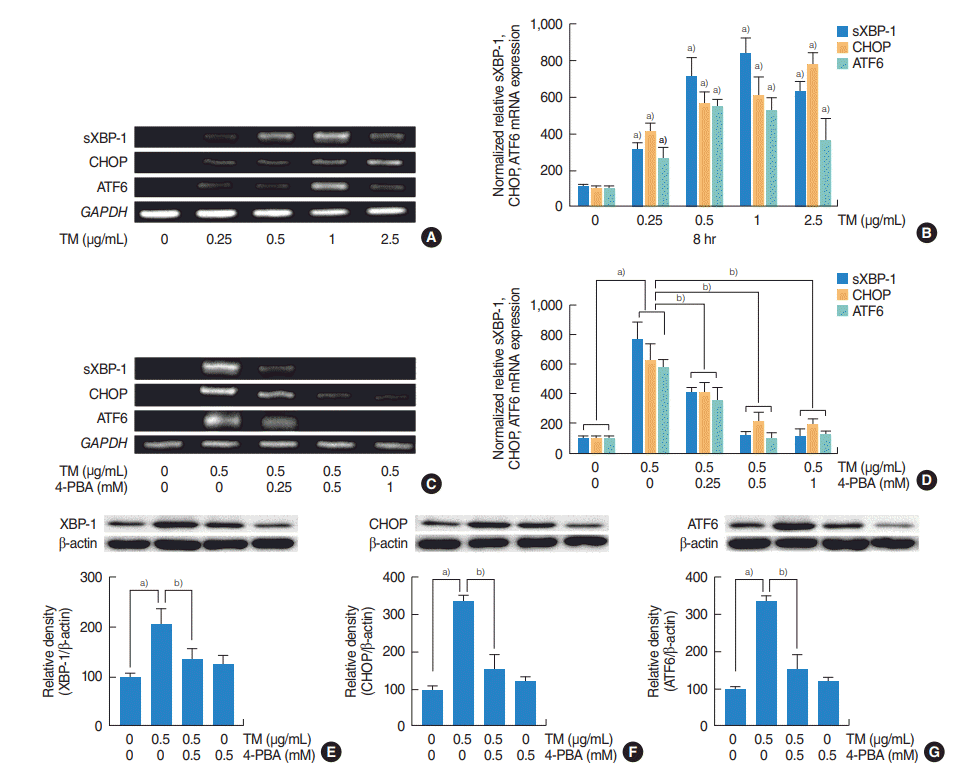 | Fig. 2.The roles of the three major endoplasmic reticulum stress-mediated signaling pathways (IRE1/XBP-1, PERK/ATF4/CHOP, and ATF6) on the expressions of MUC5AC and MUC5B in human nasal epithelial cells. (A-D) Reverse transcriptase-polymerase chain reaction (RT-PCR) and real-time PCR showed that the mRNA expressions of sXBP-1, CHOP, or ATF6 were significantly increased at all doses of tunicamycin (TM; 0.25, 0.5, 1, 2.5 µg/mL) administered and that these increases were attenuated by 4-phenylbutyric acid (4-PBA). (E-G) Western blot also showed that TM induced XBP-1, CHOP and ATF6 and attenuated by 4-PBA. The images shown are representative of three separate experiments performed in triplicate. Bars represent the averages±standard deviation of three independent experiments performed in triplicate. sXBP-1, spliced X-box binding protein 1; CHOP, CCAAT-enhancer-binding protein homologous protein; ATF6, activating transcription factor 6; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; XBP-1, X-box binding protein 1; IRE1, inositol requiring enzyme 1; PERK, RNA-activated protein kinase-like endoplasmic reticulum kinase; ATF4, activating transcription factor 4. a)P<0.05 vs. baseline. b)P<0.05 vs. TM (0.5 µg/mL) alone.
|
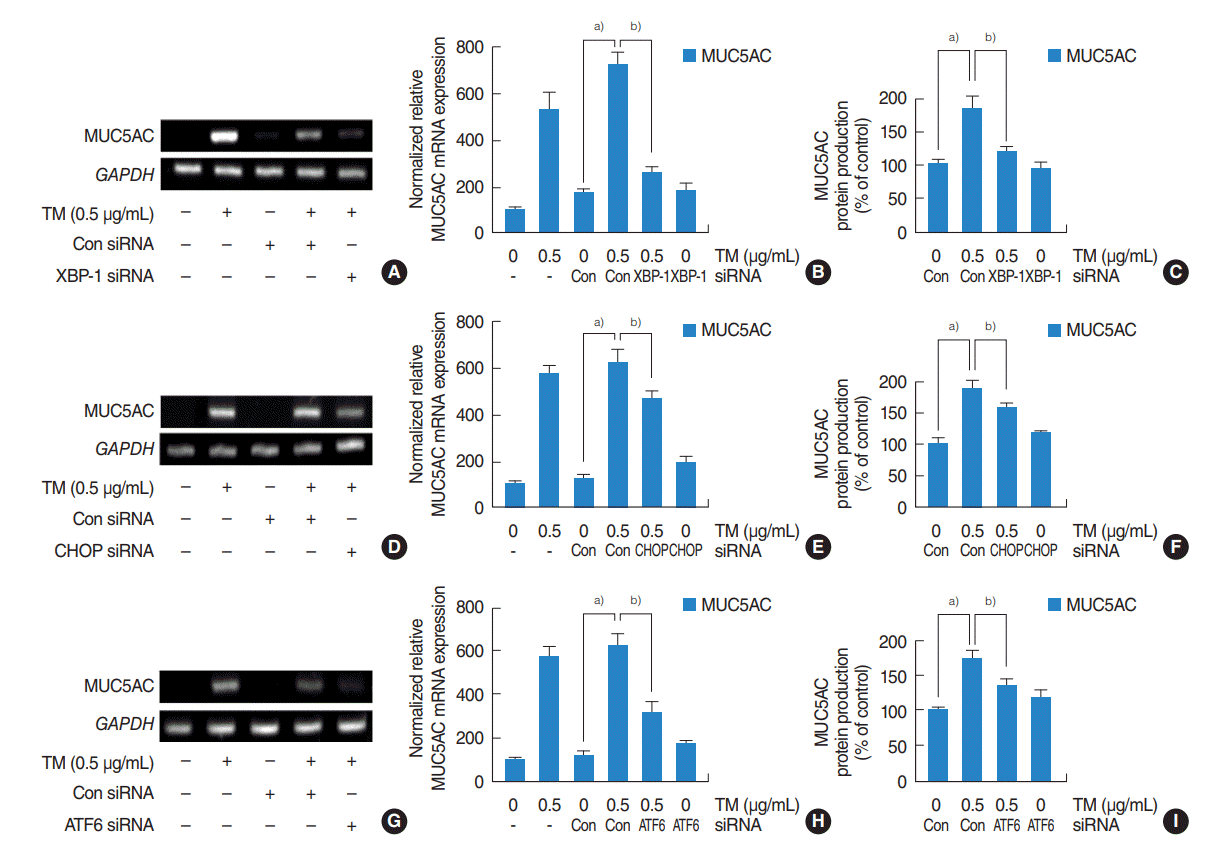 | Fig. 3.Contributions of the three major pathways to the upregulation of MUC5AC expression by endoplasmic reticulum stress in human nasal epithelial cells. (A-I) Reverse transcriptase-polymerase chain reaction (RT-PCR), real-time PCR and enzyme-linked immunosorbent assay showed that small interfering RNA (siRNA) knockdowns of XBP-1, CHOP, or ATF6 significantly blocked the endoplasmic reticulum stress-induced MUC5AC mRNA expression and glycoprotein production. (A-C) Inhibition by XBP-1 siRNA was greatest. (D-I) Inhibitions by CHOP and ATF6 siRNA were limited, and inhibition by CHOP siRNA was weakest. The images shown are representative of three separate experiments performed in triplicate. Bars represent the averages±standard deviation of three independent experiments performed in triplicate. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; TM, tunicamycin; XBP-1, X-box binding protein 1; Con, control; CHOP, CCAAT-enhancer-binding protein homologous protein; ATF6, activating transcription factor 6. a)P<0.05 vs. control siRNA. b)P<0.05 vs. TM (0.5 µg/mL) with control siRNA.
|
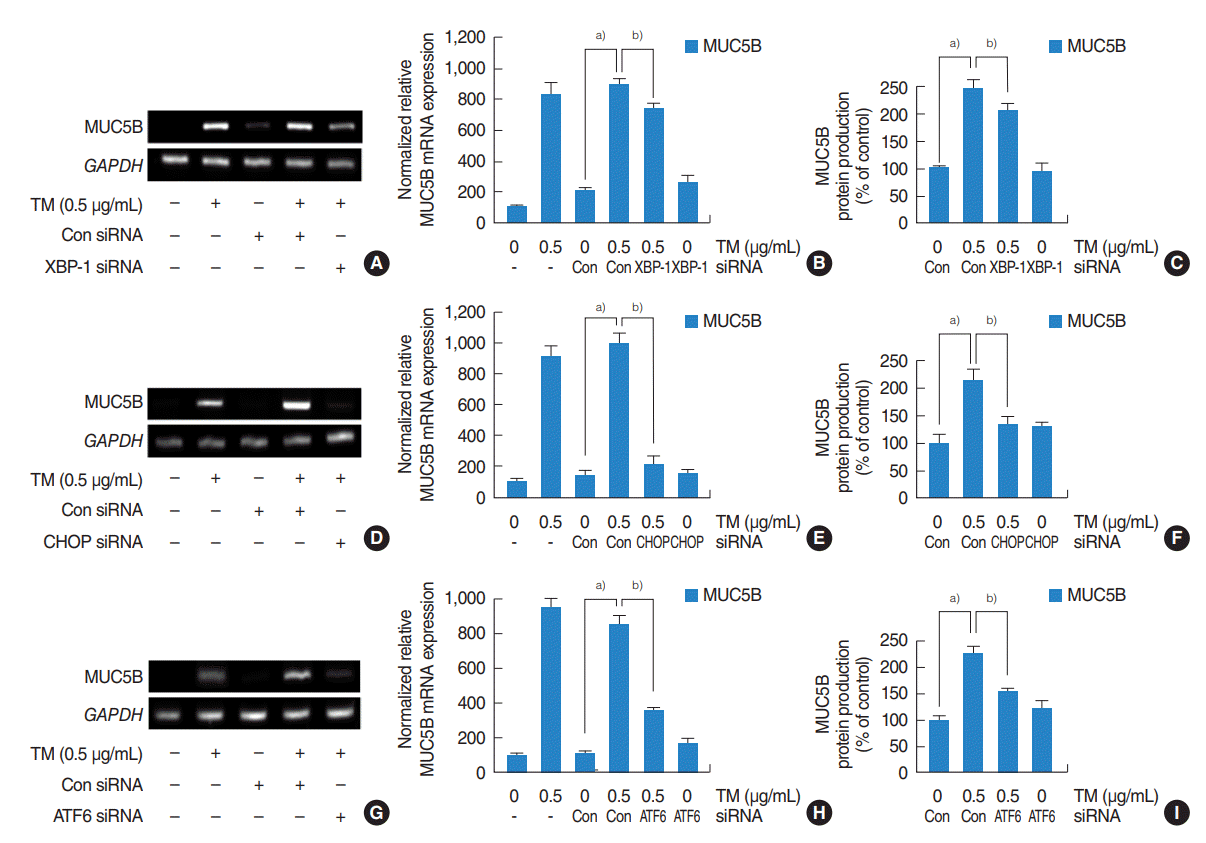 | Fig. 4.Contributions of the three major pathways to the upregulation of MUC5B expression by endoplasmic reticulum stress in human nasal epithelial cells. (A-I) Reverse transcriptase-polymerase chain reaction (RT-PCR), real-time PCR and enzyme-linked immunosorbent assay showed that small interfering RNA (siRNA) knockdowns of XBP-1, CHOP, or ATF6 significantly blocked the endoplasmic reticulum stress-induced MUC5B mRNA expression and glycoprotein production. (D-F) Inhibition by CHOP siRNA was greatest. (A-C, G-I) Inhibitions by XBP-1 and ATF6 siRNA were limited, and inhibition by XBP-1 siRNA was weakest. The images shown are representative of three separate experiments performed in triplicate. Bars represent the averages±standard deviation of three independent experiments performed in triplicate. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; TM, tunicamycin; Con, control; XBP-1, X-box binding protein 1; CHOP, CCAAT-enhancer-binding protein homologous protein; ATF6, activating transcription factor 6. a)P<0.05 vs. control siRNA. b)P<0.05 vs. TM (0.5 µg/mL) with control siRNA.
|
Go to :
DISCUSSION
ER is an organelle found in eukaryotic cells, which serves multiple functions, being important particularly in the biosynthesis, processing, and transport of proteins and lipids. ER stress is a response to mismatched protein folding, which lead by an accumulation of misfolded or unfolded proteins in ER lumen [1,2]. ER stress activates three ER transmembrane receptors, that is, IRE1, PERK, and ATF6, to maintain ER homeostasis [15]. ER stress reduces protein synthesis via the activation of PERK-eukaryotic initiation factor 2 alpha (eIF2α), but increases XBP-1- and ATF6-mediated chaperone transcription to attenuate ER stress. However, when ER stress persists, ER stress induces apoptosis by activating the PERK-ATF4-CHOP signal pathway [15,16].
ER stress is closely associated with the pathogeneses of various diseases including diabetes, obesity, cancer, autoimmune diseases, and inflammatory airway and bowel diseases [1,2,17]. Moreover, it has been reported in recent studies that ER stress-mediated inflammatory reaction in the human airway plays an important role in the initiation, progression, or deterioration of most inflammatory airway diseases, including chronic obstructive airway disease, asthma, cystic fibrosis, and chronic sinusitis with nasal polyposis [3,4,18-21]. Cigarette smoke, as a major cause of chronic airway diseases, induces ER stress by activating the PERK-eIF2α pathway [18]. In addition, the pathogenesis of bronchial asthma, especially severe steroid-resistant asthma is also associated with the activation of NF-κB mediated by ER stress [3,19]. Spliced XBP-1 mRNA mediated by ER stress is responsible for ER/Ca2+ store expansion in inflamed human bronchial epithelial cells [20]. In the recent study by Kim et al. [21], they reported that Staphylococcus aureus enterotoxin B (SEB) induced ER stress via reactive oxygen species production in airway epithelial cells. And the induction of unfolded protein response by SEB was increased significantly in eosinophilic polyp compared with noneosinophilic polyp or control mucosa. Through these results, they suggested that SEB-induced ER stress may play important roles in the pathogenesis of nasal polyposis. Furthermore, recent studies indicate that ER is involved in the production of airway mucins: The ER-resident protein anterior gradient homolog 2 (AGR2) increases with mucin overproduction in individuals with asthma and in asthma mouse models. AGR2 interact with immature mucin in the ER and loss of AGR2 impairs allergen-induced MUC5AC and MUC5B overproduction. This study suggested that AGR2 is required for production of the gel-forming airway mucin MUC5AC and MUC5B [22]. And, in the study by Martino et al. [5], it has identified that the ER stress transducer IRE1β is expressed in airway mucous cells and functionally required for the production of airway mucins. Thus, it has been shown that ER is an important organelle in the regulation of the production of airway mucins and ER stress-mediated signaling pathway is also deeply involved in the production of airway mucins. However, the precise involved mechanisms are not fully understood yet. The results of the present study demonstrate that the overproduction of the major secretary airway mucins including MUC5AC and MUC5B, which observed in CRS, are mediated by tunicamycin (an ER stress inducer), and that the tunicamycin-mediated overproductions of MUC5AC and MUC5B can be attenuated by 4-PBA (an ER stress inhibitor).
Increased secretions of MUC5AC and MUC5B glycoproteins are observed from most epithelia affected by inflammatory airway disease [8-10,23,24]. But, the specific mechanisms responsible for the up- or down-regulations of the productions of MUC5AC or MUC5B are unclear. According to recent animal studies conducted by Roy et al. [10] on MUC5AC- or MUC5B-deficient or overexpressing mice, the functional roles of these two mucin glycoproteins differ in the mouse respiratory system. It was suggested mouse MUC5B is required for controlling infections in airways and the middle ear, and for maintaining immune homeostasis in mouse lungs, whereas MUC5AC is ineffective. Thus, based on the findings of this previous study, it was expected the productions of MUC5AC and MUC5B would be regulated by different mechanisms. For this reason we focused individually of the involvements of XBP-1, CHOP, and ATF6 in the ER stress-mediated overproductions of MUC5AC and MUC5B in human airway epithelial cells. It was found that all three major ER stress-mediated signaling pathways were involved in the ER stress-mediated overproductions of MUC5AC and MUC5B and that the three pathways were involved individually in the up-regulations of MUC5AC and MUC5B in human airway epithelial cells. In addition, we confirmed that the contributions of the three major pathways to the ER stress-mediated expressions of MUC5AC and MUC5B differed, that is, XBP-1 and ATF6 were mainly involved in ER stress-mediated MUC5AC expression and CHOP and ATF6 were mainly involved in ER stress-mediated MUC5B expression.
The present study has some limitations that should be mentioned. First, assessments of the activation statuses of signaling molecules generated during ER stress were insufficient. Second, due to the lack of further experimental confirmations by using with animal model or human tissue samples, we could not identify the usefulness of clinical application of ER stress-mediated substances as a therapeutic target or biomarker in human sinonasal diseases. Therefore, we suggest additional experiments be conducted to clarify the results of the present study.
In conclusion, the present study demonstrates that ER stress induces the expressions of MUC5AC and MUC5B via the activations of XBP-1, CHOP, and ATF6. And these three major ER stress-mediated signaling networks were found to be differently involved in ER stress-mediated overproductions of MUC5AC and MUC5B in human nasal epithelial cell. These novel findings suggest that precise and specific inhibitions of the three major ER stress signaling pathways offer an effective strategy for developing therapeutics to treat chronic inflammatory sinonasal diseases.
Go to :
 XML Download
XML Download