

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Antiorthostatic suspension (AOS) is one of well-known ground based model of simulated microgravity [1]. We could perform AOS experiment by simply hanging the tail of mouse so that the hindlimbs are not bearing weight. Using their forelimbs on the ground, animal could move and get food and water [1]. Several researchers have suggested that AOS could reproduce the effect of real microgravity on the cardiovascular, musculoskeletal, and more importantly, immune systems of a living organism [2-4].
Acute lung injury is one of common clinical complication of pulmonary infection, and is the cause of respiratory failure [5]. It could be characterized as inflammation of lung parenchyme, including infiltration of neutrophils into the alveoli and increase of several pro-inflammatory cytokines such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6 [6]. As the era of ‘space travel’ is near at hand, patients with pulmonary disorders as well as healthy individuals could be exposed to long-term microgravity during space travel. However, there is still no study about the effect of long-term simulated or actual microgravity on the clinical course of acute lung injury.
Therefore, we evaluated the effect of simulated microgravity using AOS in a murine model of acute lung injury by lipopolysaccharide (LPS), by measuring: (1) weight gain before and after AOS; (2) the number of inflammatory cells and titers of cytokines such as IL-1β, IL-6, IL-10, TNF-α, and interferon (IFN)-γ in bronchoalveolar lavage (BAL) fluid; (3) titer of myeloperoxidase (MPO) in serum and lung homogenate, and finally by (4) histopathologic examination of lung tissue.
Go to : 
MATERIALS AND METHODS
Animals
Thirty BALB/c mice, 4 weeks old and free from specific murine pathogens, were used (Orient Bio, Seongnam, Korea). Throughout the whole experiment period, all mice were raised in a specific-pathogen-free environment with regular light-dark cycle and unlimited access to ovalbumin-free water and food. The experimental protocol of our study was approved by the Animal Care and Use Committee of Inha University (INHA 140919-327).
Antiorthostatic suspension
For performing AOS, we suspended a tail of mouse so that only hindlimbs are unloaded, and forelimbs remained weight-bearing (forelimbs on the ground). In order to prevent tail necrosis during AOS, we sealed around tail with sponge and tape, and protected it from being gnawed by placing a plastic tube around the tail. The angle of suspension was about 30°. As we connected a swivel pulley near the tip of tail and suspended it on a steel bar, mice could move freely along the axis of steel bar with free rotation in any direction using their forelimb.
During the period of 4 weeks, mice in control group (n=10) were raised in individual cages without any restraint during the experimental period. Mice in restraint group (n=10) were suspended by their tail, but their hindlimbs were not unloaded (touching the ground). Finally in AOS group (n=10), mice were exposed to continuous AOS.
Induction of acute lung injury
After 4 weeks, mice in restraint group and AOS group got intranasal instillation of 50 μL of 20 mg/kg lipopolysaccharide (LPS) isolated from Escherichia coli (O111:B4, catalogue #L4130; Sigma-Aldrich, St. Louis, MO, USA). Mice in control group were given intranasal challenge with 50-μL phosphate-buffered saline (PBS) with no LPS. Twenty-four hours after PBS or LPS challenge, all mice were sacrificed.
Weight gain & differential cell counts in BAL fluid
Before and after 4 weeks of experimental period, we measured the weight of each mouse. Weight gain was calculated by subtracting initial weight from the final weight after 4 weeks. Immediately after sacrifice, we collected serum by aortic puncture technique. In order to get BAL fluid, we inserted a polyethylene tube into the trachea. Then we performed lavage with sterile saline (about 3 mL) through intra-tracheal tube. We stained centrifuged preparations with Diff-Quik (Baxter Scientific, Miami, FL, USA) according to the guideline of manufacturer, and determined differential cell count for inflammatory cells such as eosinophils, neutrophils, lymphocytes and macrophages in high power field (×1,000). Two impartial, blinded practitioners counted the number of cells in each slide, and the mean value of them was used.
Cytokines in BAL fluid and myeloperoxidase assay
We measured the titers of IL-1β, IL-6, IL-10, TNF-α, and IFN-γ in BAL fluid by using appropriate ELISA kits (BioSource International, Camarillo, CA, USA), according to manufacturer’s instruction. We also performed MPO assay for serum and lung homogenates using mouse myeloperoxidase DuoSet ELISA kit (R&D systems, Minneapolis, MN, USA) according to manufacturer’s guide.
Histopathologic examination
Lung tissues were fixed in a 10% formalin solution for 3 weeks, and then embedded in paraffin. For prepared sections with 4-μm thickness, we performed hematoxylin and eosin staining and Sirius red staining. We counted the number of neutrophils infiltrated in a 1 mm2 of lung parenchyme. Again, two blinded practitioners independently counted the number of neutrophils. Then we used the mean value as the number of infiltrated neutrophils.
Statistical analysis
For the comparison of weight gain, differential cell counts and titers of cytokines in BAL fluid, titers of serum and lung MPO and the number of infiltrated neutrophils into the lung parenchyme, we adopted non-parametric tests such as Kruskal-Wallis test and the Mann-Whitney U-test. We expressed all results as means±standard deviations. We used IBM SPSS ver. 19.0 (IBM Co., Armonk, NY, USA) and a P-value of less than 0.05 was considered statistically significant.
Go to :
RESULTS
Prolonged AOS caused significant weight loss
After 4 weeks of orthostatic suspension, restraint group had significantly less weight gain compared with control group (P<0.001). After prolonged suspension of 4 weeks, AOS group had remarkable weight loss, with statistical significance compared to control or restraint group (P<0.001) (Fig. 1).
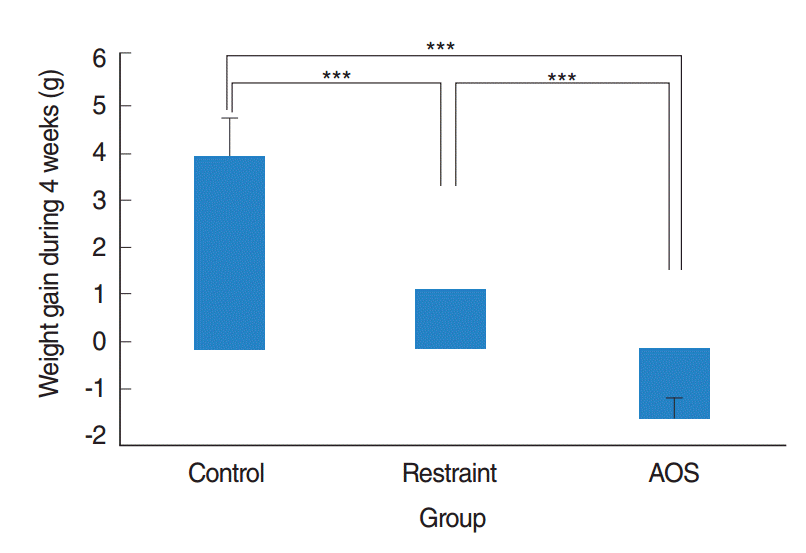 | Fig. 1.Weight gain after 4 weeks. Control group: free movement without tail suspension. Restraint group: tail suspended, but hindlimb not unloaded. Antiorthostatic suspension (AOS) group: hindlimb unloaded by tail suspension. Error bar: standard deviation. KruskalWallis and Mann-Whitney U-tests. ***P<0.001.
|
Change of neutrophils, lymphocytes, and macrophages in BAL fluid after prolonged AOS
The total cell count in BAL fluid was 340.5×103/mL in control group, 741.4×103/mL in restraint group, and 601.0×103/mL in AOS group. Restraint group showed significantly increased neutrophils, slightly decreased lymphocytes, and remarkably decreased macrophages in BAL fluid compared to control group. On the other hand, AOS showed significant increase of neutrophils and decrease of lymphocytes and macrophages, compared with control group (P<0.001 for neutrophils and macrophages and P=0.023 for lymphocytes). Comparing restraint and AOS group, there was modest tendency of decrease of all inflammatory cells in BAL fluid, although not reaching statistical significance (P>0.05) (Fig. 2).
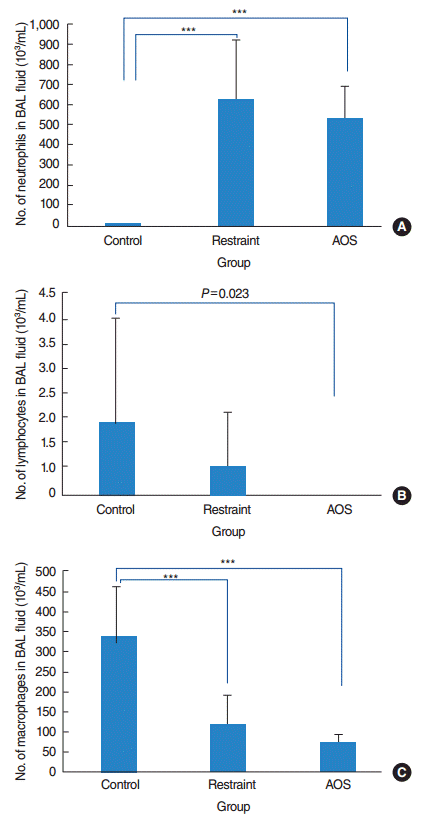 | Fig. 2.Number of neutrophils (A), lymphocytes (B), and macrophages (C) in bronchoalveolar lavage (BAL) fluid after intranasal challenge with lipopolysaccharide. Control group: free movement without tail suspension. Restraint group: tail suspended, but hindlimb not unloaded. Antiorthostatic suspension (AOS) group: hindlimb unloaded by tail suspension. Error bar: standard deviation. KruskalWallis and Mann-Whitney U-tests. ***P<0.001.
|
Significantly decreased IL-1β in response to LPS after prolonged AOS
After LPS instillation, restraint group showed significant increase of IL-1β, IL-6, and TNF-α in their BAL fluid, compared to control group. However, after prolonged suspension, AOS group showed significantly lower IL-1β compared with restraint group (P=0.049). Although IL-6 and TNF-α had modest tendency of decrease after AOS, there was no statistical significance (P>0.05) (Fig. 3).
 | Fig. 3.Titers for interleukin (IL)-1β, IL-6, IL-10, tumor necrosis factor (TNF)-α, and interleukin (IFN)-γ in bronchoalveolar lavage (BAL) fluid after intranasal challenge with lipopolysaccharide. Control group: free movement without tail suspension. Restraint group: tail suspended, but hindlimb not unloaded. Antiorthostatic suspension (AOS) group: hindlimb unloaded by tail suspension. Error bar: standard deviation. ***P<0.001.
|
AOS caused marked decrease of serum MPO but no decrease of lung MPO
Restraint group showed significant increase of serum MPO titers after LPS instillation, compared to control group (P<0.001). In AOS group, prolonged hindlimb unloading significantly reduced serum MPO titers caused by LPS instillation (P=0.004) (Fig. 4A). However, there was no significant difference of MPO in lung homogenates between restraint and AOS group (P>0.05) (Fig. 4B).
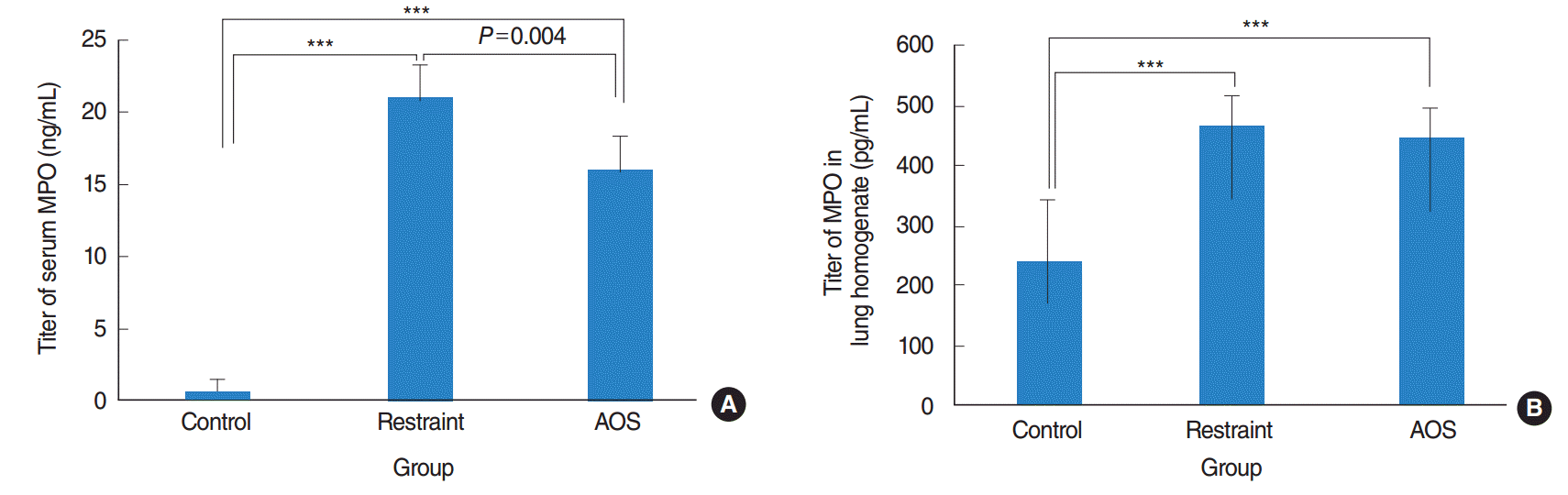 | Fig. 4.Titer for serum myeloperoxidase (MPO; A) and MPO titer in lung homogenate (B). Control group: free movement without tail suspension. Restraint group: tail suspended, but hindlimb not unloaded. Antiorthostatic suspension (AOS) group: hindlimb unloaded by tail suspension. Error bar: standard deviation. Kruskal-Wallis and Mann-Whitney U-tests. ***P<0.001.
|
Change of neutrophilic infiltration in response to LPS after prolonged AOS
Compared to control group, restraint and AOS group had significantly more infiltration of inflammatory cells in their pulmonary parenchyme (Fig. 5A). Counting the number of infiltrated neutrophils in 1 mm2 of lung tissue, we could find that there was no significant difference between restraint and AOS group (P>0.05) (Fig. 5B).
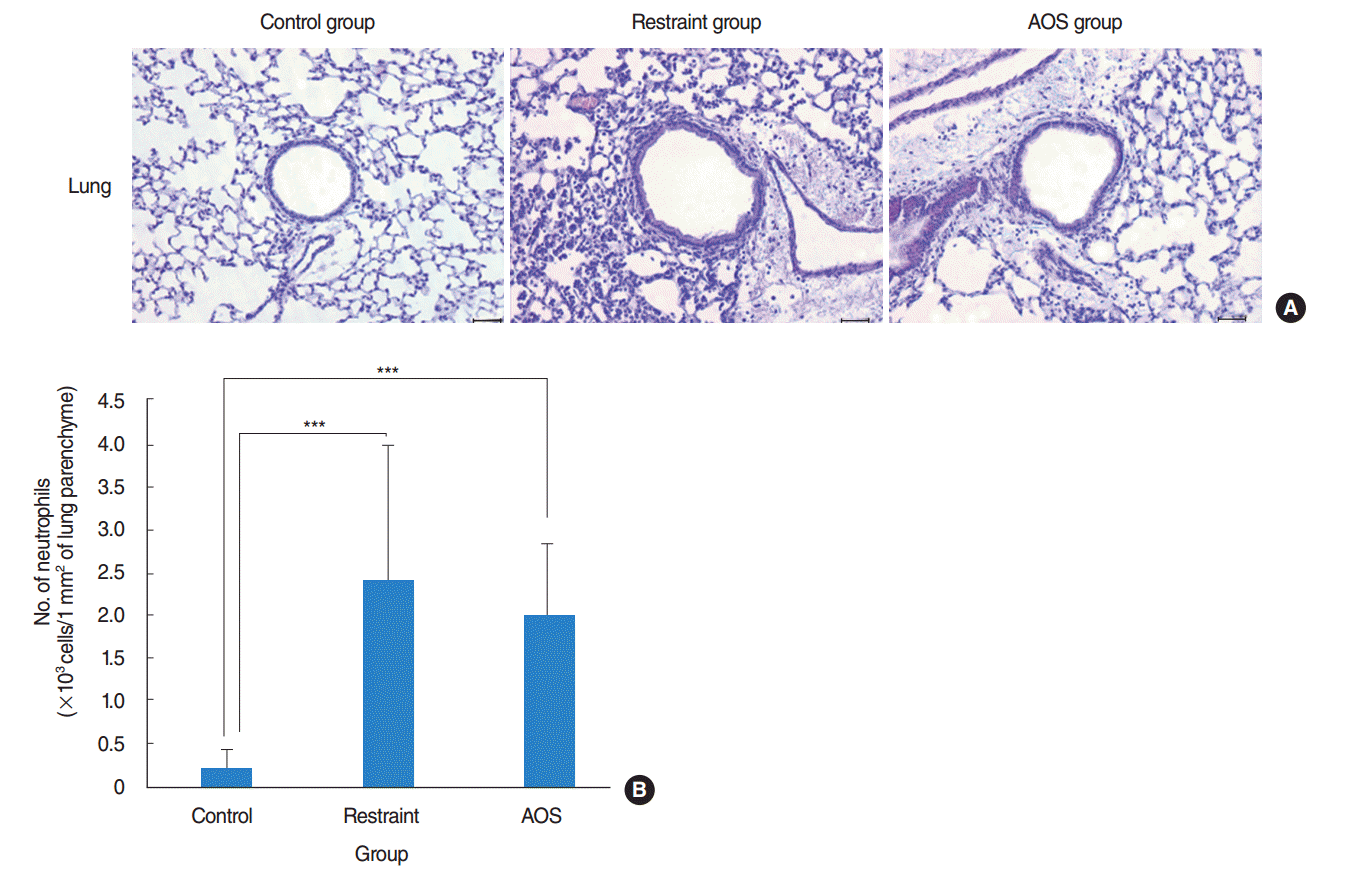 | Fig. 5.(A) Histopathologic examination of lung tissue (H&E, ×200; 50 μm). (B) Number of infiltrated neutrophils in 1 mm2 of lung parenchyme. Control group: free movement without tail suspension. Restraint group: tail suspended, but hindlimb not unloaded. Antiorthostatic suspension (AOS) group: hindlimb unloaded by tail suspension. Error bar: standard deviation. ***P<0.001.
|
Go to :
DISCUSSION
In our study, both restraint and AOS groups had significantly less weight gain compared to free control group. It is so remarkable that AOS group had significant weight loss, which was significantly different compared to restraint group. These results well concurs with previous findings. Felix et al. [2] reported significant loss of body weight after 3 weeks of AOS in experimental animals. Lew et al. [7] also found significantly less weight gain in AOS-exposed mice and suggested that the exact pathophysiologic mechanism about the effect of microgravity condition on the energy metabolism had not been solved yet. The possible cause of weight loss in AOS mice could be: (1) reduced food in take [8,9], (2) decrease of muscle mass due to disuse atrophy [10,11], and (3) increased energy expenditure [7]. In fact, space travelers suffer from anorexia and weight loss in microgravity condition [12-14]. Therefore, it is essential for him or her to increase food intake and exert muscular exercise to minimize negative energy balance.
There have been some in vitro studies about the effect of simulated or actual microgravity on the function of immune cells. However, to the best of our literature review, we could not find any in vivo study that evaluated different immune cells in BAL fluid. In our study, AOS group had significant decrease of lymphocytes in BAL fluid, which was not in the case in restraint group. It had been known that the blastogenesis of lymphocyte was down-regulated [15,16] and bone marrow cells had less capacities to respond to colony stimulating factors [17]. According to Murdaca et al. [18], when healthy volunteers took a bed rest in antigravity posture for 3 consecutive days, the proliferation of lymphocytes in their serum was significantly decreased. To our best knowledge, our study is the first one to investigate the similar change of lymphocytes in lungs, which is the main target organ in allergic asthma.
Restraint group and AOS group showed significant decrease of macrophages in BAL fluid compared with control group. However, there was no significant difference between restraint and AOS group in the number of macrophages in BAL fluid. Macrophages are the major cell population in BAL fluid of healthy animal. However, neutrophils are the predominant cells in BAL fluid. Therefore, the relative proportion of macrophages generally decreases as the proportion of neutrophils increases in BAL fluid. This relative decrease of macrophages in comparison with neutrophils is a natural phenomenon in acute lung injury. However, it does not indicate decreased innate immunity. Miller et al. [19] suggested that the profiles of macrophage activation were increased after 4 days of AOS. However, they also suggested that these results could be highly dependent on several factors, such as the duration of AOS and ability of adaptation in each individual [19]. The duration of AOS in our study was 4 weeks, which is still longer period than previous studies.
Compared to restraint group, mice in AOS group had significantly less IL-1β in BAL fluid when challenged with LPS. According to previous literatures, human monocytic cell line cultured in simulated microgravity secreted significantly less IL-1β in response to phorbol ester challenge [20,21]. On the other hand, Wang et al. [22] argued that macrophage cell lines cultured in simulated microgravity had no difference in their secretion of IL-1β compared with those cultured in normal gravity, when they were exposed to LPS challenge. Regarding TNF-α, restraint and AOS group showed no difference when exposed to LPS challenge (although some tendency to decrease in AOS group). Hsieh et al. [23] reported that murine macrophage cell lines cultured in microgravity condition secreted less TNF-α when challenged with LPS. Our study is, to our best knowledge, the first one that evaluated the effect of simulated microgravity on change of cytokines in response to noxious stimuli in vivo. It could be possible that long term simulated microgravity could have nonspecific inhibitory effect on T helper (Th) 1 pathway. However, the exact mechanism remains unsolved yet.
There have been some controversies about the change of IL-6 in microgravity. Felix et al. [2] suggested that titer of IL-6 was significantly increased in serum and lymph nodes after 1 week of AOS. According to Lew et al. [7], hypothalamic IL-6 mRNA was up-regulated after AOS. However, Hsieh et al. [23] argued that murine macrophage cell lines cultured in microgravity secreted less IL-6 in response to LPS challenge. Different experimental protocols (in vitro or in vivo, actual or simulated microgravity, duration of exposure, and so on) could cause different results. In our study, there was no significant difference in IL-6 titers between restraint and AOS group. It could be speculated that simulated microgravity had little effect on Th2 pathway in response to toxic stimuli. Also in our study, the titers for IL-10 and IFN-γ in BAL fluid was not significantly different between groups.
Neutrophils, recruited by LPS-mediated inflammation, secrete MPO. MPO, in turn, induce production of reactive oxygen species such as superoxide anion [24]. Transcription of mRNA for various peroxide genes were significantly down-regulated in plant during a space flight [25]. In our study, serum MPO in response to LPS challenge was significantly reduced in AOS group, compared to restraint group. Along with decrease of cytokines such as IL-1β, decreased serum MPO could be responsible for less pulmonary inflammation.
By histopathologic examination of lung tissue, we could find that there was only marginal difference between restraint and AOS group in the degree of pulmonary inflammation. Although there was less infiltration of neutrophils in the lung parenchyma of AOS group compared to restraint group, that difference did not reach statistical significance. LPS is known to bind the pattern recognition receptor named toll-like receptor 4 and activate intracellular signaling pathway such as nuclear factor-kappa B. Consequently, it activates pro-inflammatory cytokines such as IL-1β, promotes recruitment of neutrophils and increases reactive oxygen species [24,26-28]. Although IL-1β in BAL fluid and serum MPO was decreased by prolonged AOS, that was not enough to significantly alleviate pulmonary infiltration significantly. In fact, there was no significant difference comparing MPO of lung homogenate between restraint and AOS group. Miller et al. [19] suggested that AOS for 4 days had protective effect for Listeria monocytogenes infection through activation of macrophages. On the other hand, Belay et al. [29] suggested that mice exposed to AOS for 2 days had less resistance and higher fatality for infection of Klebsiella pneumoniae. In fact, simulated microgravity could have different effect according to target organs. Titers for cytokine in serum could be significantly increased while those in lymph node or in spleen could show absolutely no meaningful change [2]. Further study with more mice, various duration of AOS could yield more meaningful findings in the nearby future.
Simple restraint by tail could induce stress to experimental animal, although they are not antiorthostatically suspended. Therefore, for correct comparison, we set restrain group as orthostaticaly suspended mice (hindlimb touching the ground). By this design, we could exclude the effect of stress induced by restraint itself.
The immune modulatory effect of actual or simulated microgravity could be summarized as: (1) defective and unbalanced immune response due to increase of stress hormone, (2) decrease of delayed hypersensitivity, and (3) decreased resistance to bacterial infection [29]. Furthermore, stress hormone due to the distress during the actual spaceflight or prolonged AOS could cause significant weight loss. Therefore, prolonged microgravity could have a negative effect to an individual during space travel. Further research to prevent these immunologic dysregulation should be performed.
In conclusion, 4 weeks of prolonged AOS induced (1) significant weight loss, (2) significant but borderline decrease of IL-1β, and (3) significant decrease of serum MPO (but not lung MPO). However, it was not conclusive to explain AOS-associated changes in the pulmonary inflammation.
Go to :
HIGHLIGHTS
▪ Effect of simulated microgravity was examined in acute lung injury murine model.
▪ Antiorthosatic suspension (AOS) significantly decreased body weight and lymphocytes.
▪ AOS also decreased the levels of bronchoalveolar lavage IL-1β and serum myeloperoxidase.
▪ However, prolonged AOS caused no significant change in histological sections of lung.
Go to :
 XML Download
XML Download