

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Respiratory epithelial cells are the first mucosal cells exposed to the environmental stimuli that are associated with airway inflammation. These cells defend against allergens and microorganisms through physical barriers such as the tight junction, and they produce glycoconjugates, defensive proteins and enzymes that together constitute a protective barrier. They also play an active role in inflammation through the production of a variety of lipid mediators, cytokines, chemokines, the extracellular matrix and growth factors, in addition to being a major source of inflammatory chemical mediators such as granulocyte-macrophage colony stimulating factor (GM-CSF), regulated on activation normal T expressed and secreted, eotaxin, interleukin-6 (IL-6) and IL-8; all these are involved in the modulation of the local inflammatory reaction (1, 2).
House dust mites (HDM) are complex organisms that produce thousands of different proteins and other macromolecules. Over thirty different proteins of HDM can induce IgE antibody in patients who are allergic to HDM. Dermatophagoides pteronyssinus (DP) and dermatophagoides farina (DF) are major causes of allergic disease and they have been strongly implicated in the expanding prevalence of allergies. Nineteen groups of HDM allergens have been described and among the19 allergens, major IgE binding has been reported for the group 1, 2, 3, 9, 11, 14, and 15 allergens (3, 4). HDMs present to the airway by inhalation, and they then contact the airway epithelium. After this they became hydrated and discharge their allergenic contents onto the mucosal barrier, which is specialized to prevent invasion by foreign proteins. Proteases produced by the HDMs are able to play a role in a number of potentially significant pathophysiological processes; they decrease the barrier function of the epithelial cell layer, alter mucosal permeability, activate kallikrein and degranulate mast cells by a mechanism that is independent of IgE crosslinkage (4-6).
Many immune cells are located in close proximity to the epithelial cells. These immune cells and their inflammatory mediators can regulate the epithelial function. One constant feature of chronic airway inflammatory disease (bronchial asthma, sinusitis and allergic rhinitis) is the presence of tissue inflammation, and this is characterized by the infiltration of mononuclear leukocytes and notably T cells, monocytes and macrophages. Peripheral immune cells have been widely studied to provide insight into the immunophysiological regulation of the epithelial cell function. There is data that suggests that immune cells newly recruited to the site of inflammation from the circulation may be more active in the disease process than the resident cells (7, 8).
Bidirectional communication between the structural resident cells and the cells of the innate and adaptive immune system is important for the protection and maintenance of the airway function. In this study, we used an in vitro co-culture system of respiratory epithelial cells and peripheral blood mononuclear cells (PBMCs), which was a mixed immune cell population consisting of T cells, B cells and monocytes, to examine the effect of epithelial cells on the immune response of PBMCs.
Go to : 
MATERIALS AND METHODS
Nasal polyp epithelial cell culture and activation with HDM
Nasal polyps were obtained from eight patients who were undergoing endoscopic sinus surgery. Patients were excluded if they had an allergy or they had received systemic or topical steroids or antibiotics during the four weeks preceding the study. This study was approved by the Institutional Review Board of Daegu Catholic University Medical Center and each patient signed a consent form that outlined the objectives of the research and experiments. The specimens were placed in Ham's F-12 medium supplemented with 100 international units penicillin, 100 µg/mL streptomycin and 2 µg/mL amphotericin B, and they were transported to the laboratory. The nasal polyp epithelial cells were isolated using protease digestion methods. The polyps were rinsed with Ham's F-12 medium supplemented with antibiotics and they were incubated with Dispase (Roche, Mannheim, Germany) in Ham's F-12 medium for 16 hr at 4℃. After incubation, the nasal epithelial cells were isolated by gentle agitation. The cell suspensions were filtered through a No. 60 mesh cell dissociation sieve and then centrifuged at 1,000 rpm for 5 min at room temperature. The cell pellet was then resuspended in Ham's F-12 medium supplemented with antibiotics, 150 µg/mL glutamine, 5 µg/mL transferrin, 25 ng/mL, epithelial growth factor, 15 µg/mL endothelial cell growth supplement, 5 IU/mL insulin, 200 pM triiodothyronin, 100 nM hydrocortisone and 15% fetal calf serum (FCS). The cell suspensions (106 cells/mL) were plated on to six-well culture plates and these were placed in a 5% CO2 humidified incubator at 37℃. The culture medium was changed after 24 hr and then every two days thereafter. When the epithelial cell cultures reached confluence, Ham's F-12 medium was changed to RPMI-1640 medium supplemented with 10% FCS and 50 µg/mL gentamycin. The nasal epithelial cells were activated with endotoxin removed DP and DF at 10, 25, and 50 µg/mL, respectively, (Greer Lab, Lenoir, NC, USA). After 48 hr of incubation, the supernatants and epithelial cells were harvested and stored at -70℃ until they were assayed. Cytokine production (IL-8, GM-CSF) was quantified using commercially available ELISA kits (R&D System, Minneapolis, MN, USA).
Co-culture of the epithelial cells and PBMCs
The confluent nasal epithelial cells were incubated with the DP and DF for 48 hr, and then they were treated with 1% paraformumaldehyde in phosphate buffer solution for 1 hr at room temperature to inhibit the synthesis of cytokines that maintain the intercellular interaction. The nasal epithelial cells were co-cultured with PBMCs for 72 hr. PBMCs were isolated from healthy volunteers after they gave their informed consent, which was approved by the Institutional Review Board of Daegu Catholic University Medical Center. The PBMCs were isolated by density gradient centrifugation on Histopaque (Sigma, St. Louis, MO, USA). The interphase cells were harvested, washed and resuspended at 1×106 cells/ml in RPMI-1640 medium (Gibco, Rockville, MD, USA) and they were supplemented with 10% FBS. After 72 hr, the supernatants were harvested to measure the tumor necrosis factor (TNF)-α, IL-5 and interferon (INF)-γ using commercially available ELISA kits (R&D system).
The intercellular adhesion molecule 1 (ICAM-1) expression of the nasal epithelial cells and the inhibition with ICAM-1 antibody
After stimulated with the HDMs, the cell pellets were placed in cryo-tubes and 1 mL of Trizole reagent was added. RNA was extracted according to the manufacture's instructions (Roche). The RNA's purity and concentration were measured using a spectrophotometer (Beckman, Mountain View, CA, USA). One microliter of RNA was used for the reverse transcription PCR amplifications, which were performed in a PerkinElmer (Norwalk, CT, USA) thermal cycler. The primers used in this study were as follows: ICAM-1 (sense, AGG TCC ACC ACT GAC ACG TT, antisense CCA ATA TGA TTC CAC CCA, 348 bp) and β-actin (sense, CTG GCC GGG ACC TGA CT, antisense, GCA GCC GTG GCC ATC TC, 268 bp). The amplification reaction consisted of an initial denaturation step at 94℃ for 2 min, followed by three-step cycling: denaturation at 94℃ for 30 sec, annealing at a temperature optimized for each primer pair (ICAM-1: 58℃ and β-actin: 58℃) for 30 sec, and extension at 72℃ for 2 min, all for 32 cycles. The amplification products were electrophoresed on 2% agarose gels and they were visualized by ethidium bromide staining. The band densities were measured using the multi Gauge ver. 2.02 (Fujifilm, Tokyo, Japan). The band intensities were expressed as a percentage of the density of β-actin.
Immunocytochemical staining was performed with the monoclonal anti-ICAM-1 antibody (Sigma). The cytospin preparations of the nasal epithelial cells were obtained with a sample application of 1×105 cells per slide. The cytospin slides were air-dried and treated with 3% H2O2. Non-specific binding was blocked by 10% goat serum. Incubation with the diluted specific anti-ICAM-1 antibody was performed at 37℃ for 1 hr, and then this was again done overnight at 4℃ in a moist chamber. After incubation with the corresponding diluted secondary antibody, the antigen-antibody complex was visualized using a commercial AEC kit (DAKO Co., Carpentaria, CA, USA) in accordance with the manufacturer's instructions. The immunostained cell samples were counted and three visual fields on every slide were evaluated (Fig. 1). The results were expressed as the percentages of positive cells. Immunolabelling was carried out in a single series to minimize the batch-to-batch variation.
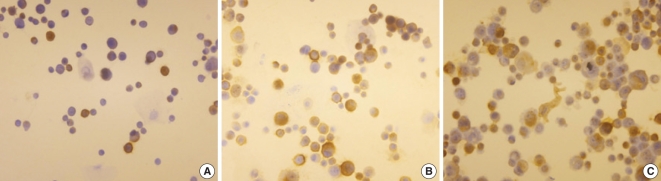 | Fig. 1The immunocytochemical findings of the nasal polyp epithelial cells treated with dermatophagoides farina (DF). (A) The nasal epithelial cells were not stimulated with DF. (B, C) The nasal epithelial cells were stimulated with 10 and 50 µg/mL of DF and the number of immune stained cells was increased.
|
Epithelial ICAM-1 was neutralized with the monoclonal antibody anti-CD54 (mAb 1379; Chemicon, Hampshire, UK). An IgG2a isotype antibody was used as a control. ICAM-1 and isotype control antibodies were added 1 hr prior to co-culture with the PBMCs. The cultured supernatants were collected after 72 hr to measure the cytokines.
Statistical analysis
All the experiments were performed at least five times and they revealed comparable results. The results are presented as the mean±SD. Statistical significance was determined using Student's t test and the Kruskal-Wallis Test (SPSS ver. 14.0; SPSS Inc., Chicago, IL, USA). A P-value of 0.05 or less was considered significant.
Go to :
RESULTS
Activation of the nasal polyp epithelial cells with HDM
In the presence of HDM extracts, the nasal polyp epithelial cells produced larger amounts of IL-8 and GM-CSF compared to the non-stimulated epithelial cells. Maximal cytokine production was found at 50 µg/mL with DF and DP. The DF showed a time dependent increase in chemical mediator production up until 48 hr, but the cytokine productions by DP were slightly decreased after 24 hr (Fig. 2).
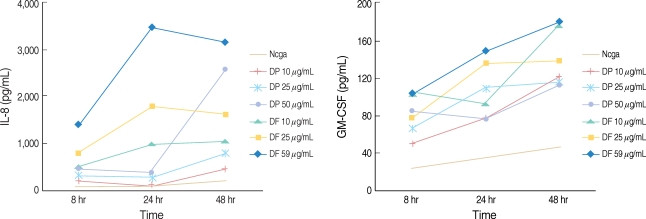 | Fig. 2Kinetic study of the interleukin-8 (IL-8) and granulocyte-macrophage colony stimulating factor production in the nasal polyp epithelial cells treated with house dust mites allergens. The cytokine productions were increased is a dose dependent manner.
DP: dermatophagoides pteronyssinus; DF: dermatophagoides farina; GM-CSF: granulocyte-macrophage colony stimulating factor.
|
Co-culture of epithelial cells and PBMCs
The activated epithelial cells were fixed with paraformumaldehyde to inhibit the cytokine secretion from the nasal epithelial cells. When the activated nasal epithelial cells were co-cultured with PBMCs, the PBMCs produced larger amounts of IL-5 and TNF-α, but not INF-γ. Compared with the co-culture of non-activated epithelial cells and PBMCs, the enhancement of TNF-α occurred at 50 and 25 µg/mL of DF, and at 25 µg/mL of DP, and the enhancement of IL-5 occurred at 50 and 25 µg/mL of DF (Fig. 3).
 | Fig. 3Production of cytokines from a co-cultrue of peripheral blood mononuclear cells and acativated nasal epithelial cells with house dust mites (HDM) allergens. Dermatophagoides farina (DF) enhanced the TNF-α and interleukin-5 production, and dermatophagoides pteronyssinus (DP) enhanced the tumor necrosis factor (TNF)-α production, but interferon (INF)-γ was not increased. The unit of the HDM extracts' concentration was µg/mL. Negative: the nasal epithelial cells were not stimulated by the HDM extracts. *P<0.05.
|
The ICAM-1 expression of the nasal epithelial cells
After stimulation with HDMs for 48 hr, relatively high values for the ICAM-1 mRNA were detected by stimulation with DP. Next, we evaluated the ICAM-1 expression on the cell membrane by performing immunocytochemical staining. After stimulation with HDMs, the epithelial cell membrane ICAM-1 expression was increased by both DP and DF (Fig. 4).
 | Fig. 4The nasal epithelial cells' expression of intercellular adhesion molecule 1 (ICAM-1) mRNA (A) and cell membrane ICAM-1 (B). The ICAM-1 mRNA expression was increased after stimulation with dermatophagoides pteronyssinus. The cell membrane ICAM-1 expression was increasd after stimulation with dermatophagoides pteronyssinus (DP) and dermatophagoides farina (DF). The unit of the house dust mites extracts' concentration was µg/mL. Negative: the nasal epithelial cells were not stimulated by the house dust mites extracts. *P<0.05.
|
To evaluate the role of ICAM-1 in the interaction between epithelial cells and PBMCs, the epithelial cells were pretreated with CD54 monoclonal antibody to block the binding of leukocyte function associated antigen-1 to ICAM-1. After 72 hr of coculture, the production of IL-5, INF-γ and TNF-α was not inhibited by ICAM-1 antibody pretreatment.
Go to :
DISCUSSION
HDMs are ubiquitous and they are a major cause of allergic rhinitis and bronchial asthma. They have been strongly implicated in the expanding prevalence of airway disease with the recent changes in the environment. When inhaled, HDMs impact upon the fluid-covered epithelial surfaces of the large diameter airways. The resulting hydration of the HDMs triggers the rapid and total discharge of major allergenic proteins. This results in a high local concentration of HDM allergens in the airway epithelium (9). The production of extracellular protease has been implicated as an important factor in the pathogenicity of HDMs. It is becoming increasingly evident that proteases from allergens like HDMs and fungi elicit the release of proinflammatory cytokines by the epithelial cells (10, 11). HDM extracts interact with the repiratory epithelial cells in several different ways, leading to morphological changes, cell desquamation and the production of the proinflammatory cytokiens IL-6 and IL-8 (10-12). The results of this study showed that the nasal epithelial cells produed larger amounts of IL-8 and GM-CSF with stimulation by the DF and DP extracts. The cytokine production was dose and time dependent. These results are consistent with those previous results showing that HDM allergens up-regulate the activity of transcription factors that are associated with the secretion of cytokines in the respiratory epithelial cells (13).
Respiratory epithelial cells play a role in the inflammatory response by releasing mediators that can directly affect the airways or influence the activity of inflammatory cells. The chemokines produced by epithelial cells enhance the migration of immune cells, eosinophils, neutrophils and dendritic cells (14, 15). A variety of immune cells are located in close proximity to the epitehlium. It has been established that immune cells and inflammatory mediators can regulate the epithelial function (16). The epithelial function under normal conditions, and particularly during inflammation, may be affected by sequential, additive or synergistic responses between various types of immune cells. The recruitment of immune cells from the circulation to the inflammatory site may promote inflammation. In the epithelium, immune cells are localized to microenvironments within either the epithelium or within the stroma below the epithelial basement membrane. The immune cells adjacent to the epithelia, via the selective expression of adhesion molecules and chemokine receptors, bind to their counter receptors and the chemokines expressed by the epithelial cells (8). Adhesive interactions may be important for both the localization and activation of immune cells. We hypothesized that HDMs activate nasal epithelial cells and they enhance the expression of ICAM-1 mRNA and membrane ICAM-1, and that these epithelial ICAM-1s may interact with the ligands of PBMCs such as leukocyte function associated antigen-1. The ICAM-1 mRNA expression was increased by DP and the cell membrane ICAM-1 expression was increasd by both DP and DF. However, anti ICAM-1 antibody did not inhibit cytokine production. This suggests that the interaction of the epitheila cells and the PBMCs may not be associated with an ICAM-1 medicated mechanism.
The hypothesis is that the T-helper cells response in allergy is skewed toward a Th2 response with the production of IL-4, IL-5 and IL-13. Studies on antigen-specific T-cell clones have indicated that the T-cell clones from allergic patients responsed to allergens with a predominantly Th2 type of response, whereas those T-cell clones from normal subjects responded with a Th1 type response (17). When PBMCs from healthy volunteers were activated by Der p 1, the production of IL-5 and TNF-α was not increased, yet the INF-γ production was significantly increased in a dose and time dependent manner (data not shown). The PBMCs from patients with atopic asthma show increased proliferation in response to HDM allergens, and atopic asthmatic PBMCs have enhanced Th2 cytokine responses to HDMs, as compared to those PBMCs from normal subjects (18). In this study, the nasal epithelial cells were activated with HDMs and then they were co-cultured with PBMCs. The PBMCs produced large amounts of the proinflammatory cytokine TNF-α and the Th2 cytokine IL-5. These results suggest that the activated epithelial cells interacted with the PBMCs and this enhanced the Th2 type inflammatory reaction. Comparison of the DP and DF showed that the DF appeared to be the stronger inducer of allergic inflammation. Although we did not investigate the underlying mechanism of interaction, epithelial preactivation with the HDM allergens appeared to be an important factor for the induction of allergic inflammation. The PBMCs contain lymphocytes, monocytes and dendritic cells, and further studies that will focus on these cells are needed to determine which immune cells play a key role in the interaction between the epithelial cells and the PBMCs.
In summary, we activated nasal epithelial cells with HDM antigens. The activated epithelial cells expressed greater amounts of adhesion molecule mRNA and protein. When the activated epithelial cells were co-cultured with PBMCs, the PBMCs produced larger amounts of TNF-α and IL-5. Therefore, HDM allergens might induce allergic inflammation by activating the nasal epithelial cells without any direct contact of the HDM allergens with the PBMCs.
Go to :
 XML Download
XML Download