

 PDF
PDF Citation
Citation Print
Print



Introduction
After the first report of unintentional neural damage following anesthesia during childhood, there is now a large amount of data regarding neurotoxicity in the developing brain [12]. While it is relieving to know that recent clinical studies suggest that a single, short exposure may not affect neurodevelopment [34], there is still not enough evidence to rule out the possibility of the negative effects of anesthesia on neuro development. Accordingly, the FDA has recently reported two safety announcements focusing on multiple, lengthy sedation or general anesthesia in children and pregnant women (https://www.fda.gov/Drugs/DrugSafety/ucm532356.htm). However, Andropoulos and Greene [5] have also reported concerns of such announcements, since it may also delay necessary surgical and diagnostic procedures that require anesthesia, resulting in adverse outcomes for patients [5].
There are very few clinical studies focusing on the effects of anesthesia in the fetal brain. Immense neurodevelopment, such as neurogenesis and neural migration, begins eight weeks after conception and continues throughout the second trimester of pregnancy [6]. Since these processes are regulated through GABA and glutamate, it is possible that anesthetic agents may disrupt this highly regulated process [7]. Studies in gestational day (GD) 14–20 pregnant mice, which represent the late-first or early-second trimester of human pregnancy (http://www.translatingtime.net/home) [8], report conflicting results regarding long-term behavioral consequences after fetal anesthetic exposure [9101112]. Unfortunately, there is little understanding on why such conflicting results exist.
Among the many factors that may contribute to the conflicting results regarding long-term behavioral consequences from fetal anesthesia, we first focused on the age of mice at which behavioral assays were actually performed, based on the fact that behavioral consequences after fetal anesthesia were mostly reported in young postnatal day (PND) 30 mice, while older mice do not show differences [711121314]. We also examined how sex-dependent neurodevelopment influences neurotoxicity during fetal anesthesia by separately comparing young male and female offspring mice [1516]. Thus, in order to evaluate the importance of age and sex when studying the long-term consequences of early anesthesia during pregnancy, pregnant mice were exposed to sevoflurane as previously described and fear chamber tests were conducted with male and female offspring mice at PND 30–35 [12].
Materials and Methods
Animals
All experiments were performed after being approved by the Committees on Animal Research in our institute (no. 014-A0009). C57BL/6J mice were housed in a regulated environment (22℃, 24 hour light cycle) and had access to food and water ad libitum. Mice were weaned at 3 weeks and caged in groups of three to five.
Anesthesia
Sevoflurane exposure was performed as previously described [12]. Briefly, pregnant female mice (GD 14) were randomly divided into 2 groups. Mice in the control group were placed in a 1-l plastic chamber and exposed to 100% oxygen (4 L/min) for 2.5 hours for 3 consecutive days (GD 14-16). Sevoflurane group mice were treated identically, except that 2.5% sevoflurane was added during the first two hours. The anesthesia chamber was placed in a water bath (36℃) for temperature control and gas was analyzed using a S/5 compact anesthetic monitor and an e-CAiO gas analyzer module (Datex-Ohmeda, Helsinki, Finland).
Behavior tests
Behavioral tests were performed with two cohorts of offspring mice obtained from eight pregnant mice that received multiple exposures of oxygen or sevoflurane (four mice each). The open field test and fear chamber test were performed as previously described with both male and female offspring at PND 30–35 [121718]. There was at least a one day resting period between tests. All tests were recorded and later analyzed with a tracking software (Ethovision XT, Noldus Information Technology, Waxgeningen, the Netherlands & Freezeframe, Actimetrics, Wilmette, IL, USA).
Open field test
General activity was evaluated after placing the mice in the center of a white cylinder (diameter 30 cm, height 40 cm). Locomotion was analyzed for 60 min with a tracking software (Ethovision).
Fear test
The fear test was conducted with a fear chamber (Coulbourn Instruments, Holliston, MA, USA) as previously described. Fear conditioning was performed 5-minutes after placing mice in the fear chamber: three trials of a conditioned stimulus (CS, 20-second long, 3 kHz, 80 dB tone) paired with an unconditioned stimulus (US, 1 second long, 0.5 mA electrical shock) at a 60-second interval. Contextual-fear memory was measured 24 hours later after being placed in the same chamber for 5 minutes. Cue-memory was analyzed after another 24 hours by placing the mice in a novel chamber. CS was applied for 3 minutes after a 5-minute habituation period.
Statistical analysis
Statistical software R (3.1.2; R Core Team, Vienna, Austria) was used for analysis as previously described [121718]. Continuous variables were tested for the assumption of normality and homogeneity of variance. Kruskal-Wallis test, Whelch's t test, and t test were performed accordingly. Welch's t test was performed when homogeneity of variance was unmet, and the Kruskal-Wallis test was performed if normality was unmet. Student's t test was performed only when both conditions were met. Mixed effect modeling was used for analyzing repeatedly measured data (fear conditioning during the fear chamber test). Differences with P < 0.05 were considered statistically significant.
Results
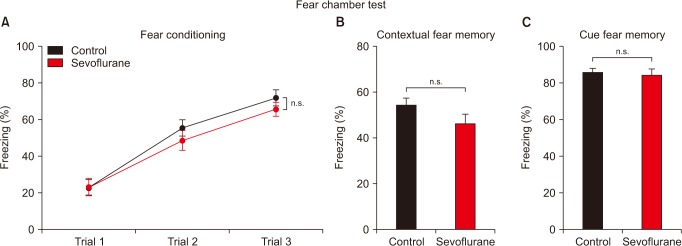
In order to evaluate the possibility that sevoflurane exposure during pregnancy induces age-dependent learning and memory impairments, we performed the fear chamber test in PND 30–35 mice. Previous studies regarding neurotoxicity during the fetal period have reported long-term behavioral impairments in this particular age, while several other studies focusing on older mice (over eight weeks) showed normal cognitive function. However, young mice that received multiple sevoflurane exposures (sevoflurane group) displayed similar learning and memory throughout the fear chamber test (Fig. 1). Fetal exposures to sevoflurane did not affect fear conditioning in young offspring mice (Fig. 1A), which is used to examine learning ability by analyzing the increase of fear behavior during three trials of a CS (20-s long, 3 kHz, 80 dB tone) and an US (1 mV shock) [12]. Long-term memory was assessed by measuring contextual-, cue-memory during the 2 following days [12]. Contextual-fear memory (measuring freezing behavior while exposing the mice to the same fear chamber) and cue-fear memory (measuring freezing behavior in a new context after applying a CS) were also not affected from fetal sevoflurane exposures in young offspring mice (Figs. 1B and 1C).
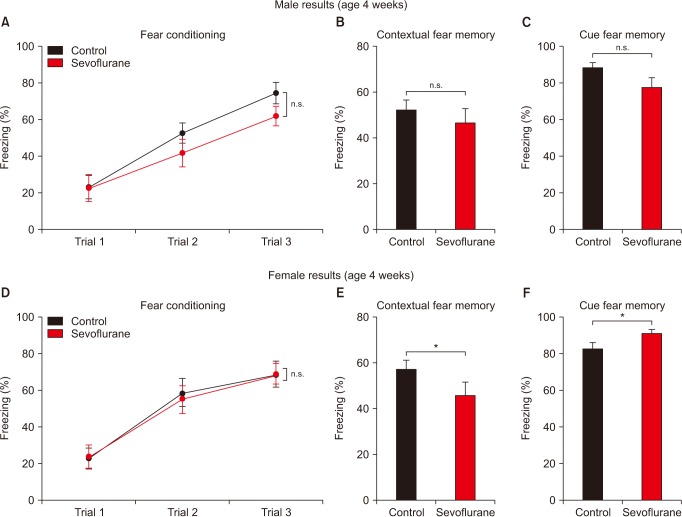
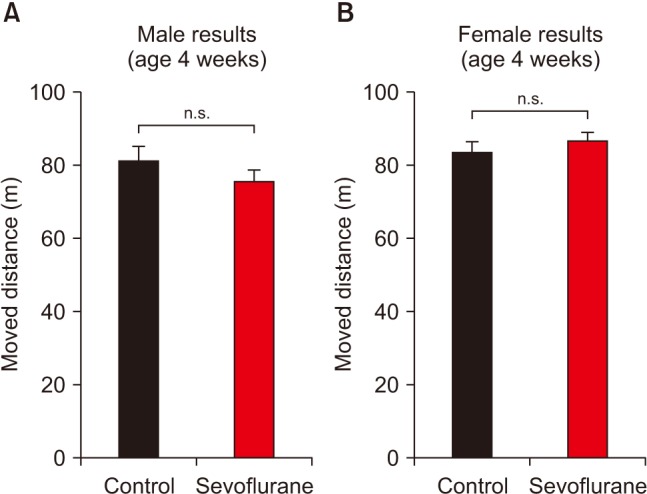
Sex has also been implied to be an important factor for anesthetic neurotoxicity [1516]. Thus, in order to exclude sexual differences, we performed additional analysis after separating male and female mice. Both male and female mice show normal learning during fear conditioning (Figs. 2A and 2D). Interestingly, while male mice show normal long-term memory (Figs. 2B and 2C), sevoflurane exposed female mice display significant changes in fear memory compared to control group female mice (Figs. 2E and 2F). Fetal sevoflurane exposures significantly impaired contextual-fear memory in young females (Fig. 2E). Unexpectedly, cue-fear memory was significantly increased in young female mice who received early anesthesia (Fig. 2F). The difference between male and female mice was not due to differences in general activity, as shown in the open field test (Fig. 3).
Discussion
With the growing awareness of potential negative effects after anesthesia in children and pregnancy, many parents and health givers express concerns of possible long-term neurodevelopmental complications after anesthesia during childhood. Even though recent studies suggest that a short, single episode of general anesthesia does not affect neurodevelopment [34], there still remains the possibility of negative effects after multiple or lengthy exposure to anesthetics [19]. We have recently reported that both single and multiple sevoflurane exposures during the fetal period does not affect behavior at an adult age in male mice [12]. Our results may cause confusion, since they do not agree with previous studies that used the same anesthetic procedure [910]. On the other hand, by understanding the cause of such conflicting results can help us gain valuable insight regarding anesthesia induced neurotoxicity.
We first focused on the actual age at which behavior was assessed. We considered age as an important factor based on several facts. Most studies that show negative long-term behavioral consequences from fetal anesthesia were performed at PND 30, while studies performed at a later age reported no differences [711121314]. Secondly, learning and memory impairments after early exposure have been shown to be reversible by raising the mice in a stimulating environment [10]. It is possible that anesthesia-induced cognitive impairments may self-resolve as the mice continue to grow and develop. However, multiple sevoflurane exposures during the fetal period did not induce changes in young offspring mice (Fig. 1).
Neurodevelopment is dependent on many factors. Although often disregarded, sex greatly influences diverse biological functions including neurodevelopment [202122]. Interestingly, sexual dimorphic neurodevelopment has also been implied to be involved with neurodevelopmental disorders and various neurotoxic insults [2123242526], including neurotoxicity from anesthesia [15162728]. Thus, we next examined how sexually dimorphic neurodevelopment affects learning and memory after multiple sevoflurane exposures in pregnant mice (Fig. 2). Interestingly, our results suggest that early sevoflurane exposures during the fetal period impairs long-term contextual memory only in young female mice, which agrees with a previous study suggesting that female mice may be more vulnerable to anesthesia-induced neurotoxicity [27]. Thus, our study emphasizes the importance of age and sex when studying the long-term effects of sevoflurane exposures. However, our results also show that cue-fear memory is increased in young female mice who received early anesthesia, complicating the interpretation of our data. Since estrogen has been shown to affect cue-fear memory, our conflicting results may be due to the influence of the estrous cycle [29], which can occur by PND 30 [30]. Unfortunately, we are presently unable to present a clear explanation why such differences occurred.
There are several limitations in the present study. Although we are now able to suggest that age does not affect fear chamber tests in male mice after early anesthesia, we are still unable to suggest the same in female mice. It is still possible that older female mice may show impaired learning and memory. Also, we still do not know if our results can be replicated in other learning and memory behavioral assays such as the Morris water maze.
In conclusion, multiple sevoflurane exposures in the second trimester of pregnancy affects learning and memory only in young female mice. We suggest that age and sex must be considered when examining long-term effects of early exposures to anesthetics. As great interests are being focused on gender differences in many clinical fields, more detailed study designs focusing on the sexual difference in the form of inflammatory markers, cytokines, hormonal levels, and neurodevelopment may help understand the sex-dependent findings of the present study.
 XML Download
XML Download