

 PDF
PDF Citation
Citation Print
Print



Introduction
Several types of receptors for modulating acetylcholine (ACh) release exist at the neuromuscular presynaptic membranes [1,2]. These receptors function together to control the release of ACh during rest or evoked neuronal stimulation [2,3]. Among them, muscarinic and purinergic (adenosine, [ADO]) receptors interact at the presynaptic neuromuscular junction. Of these receptors, the adenosine A1 and A2A receptors in the motor neurons contribute to the efficiency of neuromuscular transmission [4–6]. These ADO receptors, together with presynaptic muscarinic ACh receptors (mAChRs), modulate ACh release upon neuronal firing and regulate each other as a facilitatory or inhibitory receptors [5–7]. Adenosine A1 and A2A receptors are activated at different ADO concentrations, exerting different modulatory functions. Presynaptic inhibitory A1 and facilitatory A2A receptors mediate the fine-tuning mechanism upon tonic activation of M1 and M2 subtype of mAChRs that have facilitatory and inhibitory functions, respectively [5,8].
The train-of-four (TOF) stimulation, the conventional neuromuscular monitoring mode, is induced via indirect 2 Hz stimulation conducted for 2 s, and the TOF ratio (TOFR) corresponds to the ratio of the fourth twitch tension over the first twitch tension [9]. At such low-frequency stimulation, the inhibitory A1 receptors have the dominant function [6]; however, when the stimulation frequency is increased to 50 Hz or higher, which is often used to determine the depth of block during deep neuromuscular blockade [9–11], the amount of ADO accumulated at the synapse site drastically increased, and eventually reached levels capable of activating the excitatory ADO A2A receptors, which counteracts the M1 receptors and potentiates M2 inhibitory receptors [6].
Sugammadex, a γ-cyclodextrine derivative, can reverse the effect of aminosteroidal neuromuscular blocking agents by encapsulating and inactivating them [12–14]. This effect does not influence the amount of ACh at the neuromuscular synaptic junction, which is different from the mechanism of classic reversal strategy to increase the ACh by using cholinesterase inhibitors such as neostigmine [15]. As such, we hypothesized that, during sugammadex-induced recovery from the neuromuscular blockade, the muscle tension might be influenced if the presynaptic release of ACh is modified.
Accordingly, the primary objective of this study was to assess the changes in the first twitch tension after TOF stimulation (T1) and TOFR obtained during rocuronium-induced nuromuscular blockade after activating neuromuscular presynaptic A1/A2A receptors. Furthermore, we obtained recovery profiles during the sugammadex-mediated recovery from the rocuronium-induced neuromuscular blockade. Therefore, we investigated whether the A1 receptor-specific agonist, 2-chloroadenosine (CADO), contributes to the effective concentration for 95% blockade of T1 (EC95) and TOFR by the rocuronium. Moreover, we studied whether CADO contributes to sugammadex-induced recovery from the neuromuscular blockade.
Materials and Methods
General study design and sample preparation
The study protocol for this experiment was approved by the Ethics Committee of the Laboratory of Animal Research, Asan Institute of Life Science, Seoul, Korea, on September 1, 2015 (Protocol No. 2015-14-076). All animals were raised at a constant temperature of 22°C with a regular diurnal cycle and food and water supplied ad libitum. The phrenic nerve-hemidiaphragm tissues were obtained and immersed in Krebs buffer solution (120 mM NaCl, 2.5 mM CaCl2, 4.7 mM KCl, 2.1 mM MgSO4, 1.2 mM KH2PO4, 25 mM NaHCO3, and 11 mM α-D-glucose) maintained at 35°C and 95% O2 and 5% CO2 with continuous bubbling to ensure tissue viability throughout the experiment. Sizes and weights of each tissue were measured and compared between groups (Table 1). CADO was purchased from Sigma-Aldrich Korea LTD (Korea). The CADO stock solution was stored at −20°C and thawed only before using. Stock solutions were discarded after two weeks of their preparation.
Study protocol for main study
Thirty male Sprague-Dawley rats with average weight of 354.8 ± 36.9 g (range 298.5–438.9 g) were used in the study. Each rat was anesthetized using intraperitoneal injection of 50 μg/g tiletamine (Zoletil 50®, Virbac SA, France). The thoracic cage was immediately isolated, and the phrenic nerve-hemidiaphragm tissue was obtained. The tissues were fixed to a frame with electrodes and then immersed in a 100-ml organ bath containing 75-ml oxygenated Krebs buffer solution. The tendinous portion of the diaphragm of each sample was hooked to a Grass FT03 Force Transducer (Grass Technologies, USA), and 40 mN of resting tension was applied. The phrenic nerve was fixed to a platinum bipolar electrode and stimulated using a Grass S88 Stimulator (Grass Technologies, USA). Supramaximal stimulation using a square wave pulse of 0.2 ms was administered every 20 s at train of 2 Hz for 2 s (TOF stimulation). All waveforms were displayed and stored using the PowerLab 4/26 Data Acquisition System (AD Instruments, Australia) and LabChart 7 (AD Instruments, USA).
Phrenic nerve-hemidiaphragm tissues were randomly allocated to either the control group (CADO, 0 μM) or the CADO group (CADO, 10 μM). Twitch tensions were serially monitored during a 30 min stabilization time. After 30 min of stabilization, the same volumes of control fluid or CADO stock solution were added into the organ bath to obtain data on the allocated groups. One hour of reaction time was allowed in each group before adding a 300 μg loading dose of rocuronium (Esmeron®, MSD Korea, Korea). A 150 μg boost dose of rocuronium was subsequently added into the organ bath. The subsequent boost dose was added when five consecutive T1 depressions were < 3% of previous T1 twitch tension, or when 10 min had passed since the previous dose was added. The boost doses were stopped when ≥ 95% T1 depression was achieved. The loading dose was set to a dose at which the T1 twitch tension did not change after administration of rocuronium and the TOFR changes were within 3% of those obtained before administration of the loading dose. Boost dose was set to a dose at which change of the T1 twitch tension was observed and the total count of boost dose administration was within 10. After confirming that the T1 twitch tensions were disappeared, 15 min of resting time was allowed before introducing sugammadex to induce recovery from the neuromuscular blockade. The amount of sugammadex was equal to the equimolar doses of rocuronium that was used for the dose-response reaction for T1 twitch tension depression. Recovery profiles (T1 recovery time, recovery index [RI], and TOFR) were obtained during next 1 h. During that period, we set the zero time at the T1 twitch tension recovery of 10% and then the time interval from that time to the T1 tension recovery of 25%, 50%, 75%, and 95% were recorded. After confirming that the T1 twitch tensions had recovered ≥ 95%, 50 Hz tetanic stimulations were applied and the tetanic fades were recorded. This study protocol is summarized in Fig. 1.
Statistical analysis
The first aim of this study was to compare the concentration of rocuronium that was required in each group to attain a T1 depression by ≥ 95%. To achieve this goal, we had serially increased rocuronium concentration Krebs buffer by adding the loading and boost doses of the drug at regular intervals while recording twitch tension; these responses were plotted and expressed as regression curves and compared between groups. While attaining T1 depression, TOFRs were also simultaneously obtained and their plotted regression curves were compared between groups. The second aim was to compare the variables obtained during sugammadex-induced recovery from neuromuscular blockade. In clinical practice, the RI (the interval of T1 recovery from 25% to 75%) is a good marker for comparing recovery rates from neuromuscular blockade. As such, between-group comparison of the time intervals of T1 twitch tension recovery at 10%, 25%, 50%, 75%, and 95% was performed.
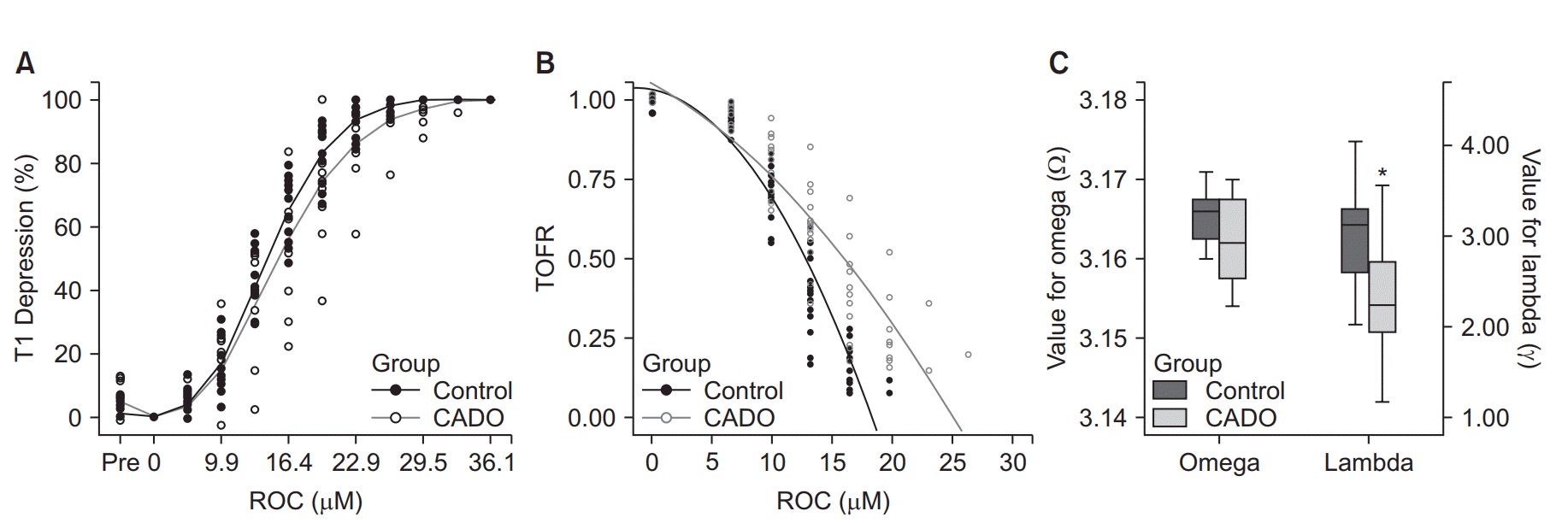
Results are expressed as mean (SD) or standard error of mean (SEM). All doses were converted and expressed as in μM. Graphs were plotted, and statistical analyses were conducted using SPSS Statistics for Windows, ver. 13.0 (SPSS Inc., USA). Rocuronium EC50 and EC95 values for twitch tension data were calculated by fitting nonlinear regression curves to group data. We used the following equation for calculating T1 depression: y = 50 + 50sin(Ωx), where y represents the T1 depression, x represents the concentration of rocuronium, and Ω represents the slope of the regression curve (R2 = 0.87). The following equation was used for calculating TOFR recovery: y = 1 − λx2, where y and x represent TOFR and rocuronium concentration, respectively, and λ represents the slope of the regression curve (R2 = 0.91, and 0.83 in the control and CADO groups, respectively). The mean group values of Ω and λ were compared using the Mann-Whitney U test. Rocuronium EC50 and EC95 values for twitch tension data were calculated by fitting linear regression curves to group data. P values < 0.05 were considered to be statistically significant.
Results
The mean size and weight of the phrenic nerve-diaphragm tissue preparations of the control and the CADO groups were not statistically different (Table 1). No significant differences were observed when comparing the rocuronium concentration-responses using the T1 twitch tension depression between the control and CADO group (P = 0.117, Fig. 2A). In addition, there were no statistical differences between the slopes of these regression lines (Table 2). One-half maximum effective concentration (EC50) and the effective concentration for ≥ 95% blockade of T1 twitch tension (EC95) were calculated using the equation for the concentration-response regression line of each group (Table 2); however, there were statistically significant differences in the TOFR change during rocuronium concentration-response. When comparing the regression line of TOFR in the control and the CADO group, there were statistically significant difference in the λs, which are the representatives of each regression curve (Figs. 2B and 2C, P = 0.002).
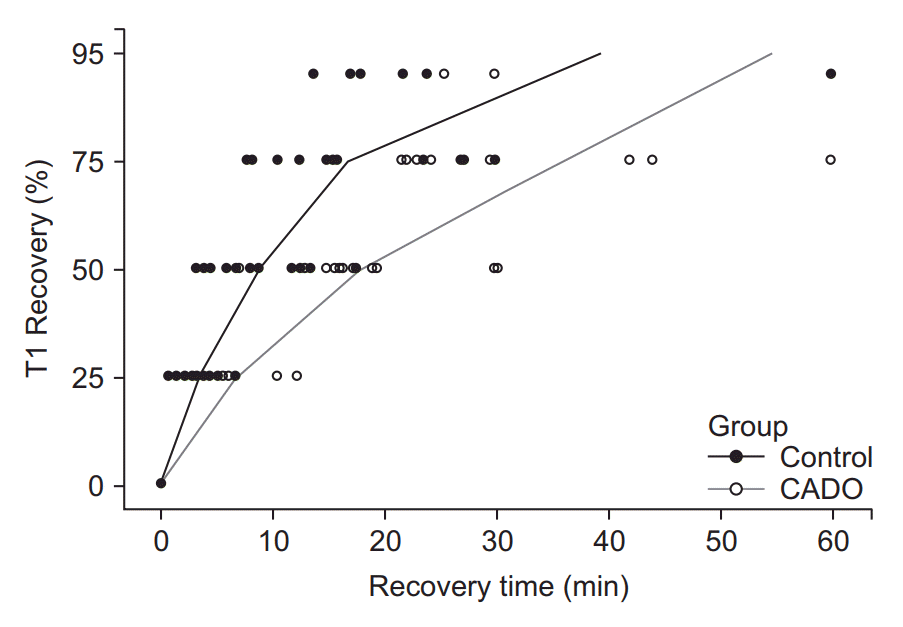
The recovery profiles were also evaluated. The zero point was set at the time at which the T1 twitch tension was recovered to 10% of initial tension, and the time intervals from this point to tension recovery up to 25%, 50%, 75%, and 95% were recorded. These time intervals in the CADO group were significantly higher than those in the control group (P < 0.01, Table 3, Fig. 3).
Discussion
The current ex-vivo phrenic nerve-hemidiaphragm study showed that presynaptic adenosine A1/A2A receptors play a role in the degree of neuromuscular blockade. CADO is listed as the selective A1 receptor agonist according to the manufacturer; however, it has often been used as a nonselective A1/A2A agonist. Although ADO acts as an A1 agonist at low concentration (300 nM), it also acts as an A2A agonist at high concentrations > 1 μM) [4]. Similarly, CADO is also used based on these concentration and result in different outcomes at different concentrations since it is a metabolically stable analogue of ADO and has the same affinity as ADO for adenosine receptors [16]. Several studies have reported conflicting results on the effect of presynaptic purinergic receptor activation on ACh release. With micromolar concentrations of ADO, the evoked quantal release or spontaneous release of ACh was decreased [16,17]; whereas, the opposite effect was exerted when submicromolar concentrations of ADO at the neuromuscular junction were made [18]. However, the inhibitory coefficients (Ki) for rat A1 and A2A receptors were 300 and 80 nM, respectively [19]. In other words, the affinity of CADO to A1 receptor is relatively lower than that to A2A receptors; hence, A2A receptor activation might occur first at lower CADO concentrations. In this study, we found that the activation of neuromuscular presynaptic A1/A2A receptors play a remarkable role in the twitch tension depression and TOF fade induced by rocuronium, which was demonstrated by the continuous activation of adenosine A1 receptors with CADO. This result is consistent with the fact that higher CADO concentrations (e.g., 10 μM) tend to reduce ACh quantal content [16], which might be the influence of the inhibitory A1 receptors. In fact, their inhibitory action was more prominent in the recovery profile in our experiment; however, other studies have reported that the equilibrium dissociation constants (Ki) of CADO at A1 and A2A receptors in the rat were 6.7 and 76, respectively [20], which contradict what we previously mentioned. Although CADO is the most commonly used agonist for this type of experiment, it is more preferable to select an agonist that is more specific to A1 receptors to better discriminate among the results.
The effect of ADO receptors on ACh release may be indicative of the potentiation of neuromuscular blockade through volatile anesthetics such as sevoflurane. Enflurane and sevoflurane have the ability to activate ADO A1 receptors in an in-vitro culture of the rat hippocampus [21]. This effect is suggested to be mediated by an interaction of volatile anesthetics with ADO transport or a key enzyme in ADO metabolism. Aminophylline is a nonselective antagonist of the adenosine receptor [22] and can decrease the sedative effects of sevoflurane [23,24] but not desflurane-induced anesthesia [25]. Sevoflurane is known as one of the most potent volatile anesthetics for potentiating the action of the neuromuscular blocking agents [26]. In our results, the recovery from neuromuscular blockade using sugammadex was delayed when the ADO receptor was activated by adding CADO in the organ bath; therefore, the potentiation effect of sevoflurane on neuromuscular blockade might be modulated by activation of the A1 receptor. The present study implicated that TOFR might be altered, showing delayed recovery from neuromuscular blockade when the A1 receptor is modulated by agonists, such as sevoflurane, or antagonists, such as aminophylline.
This study has several limitations. First, we performed this experiment in an ex-vivo environment, which lacks pharmacokinetic properties. Tissue specimens were exposed to a short period of hypoxic insult or to a hypothermic environment during tissue preparation. Attempts to prevent this exposure included shortening the manipulation time and maintaining the temperature of the buffer solution by externally circulating warm water at the organ bath. Moreover, during the experiment, circulation in the tissue was halted, and external convection current of the Krebs buffer solution made by bubbling of oxygen/CO2 mixture gas was the sole means to deliver the drugs to the neuromuscular junction. To compensate for this drawback, we allowed sufficient time after adding the study drugs; we allowed 30 min for tissue stabilization, 1 h for CADO reaction time, 10 min for rocuronium reaction, and 1 h for complete recovery from neuromuscular blockade by sugammadex. We selected CADO as the adenosine receptor agonist in this study because most experiments evaluating adenosine receptor in the neuromuscular junction have used CADO as an agonist. CADO, a nonselective agonist of all types of ADO receptors, is often used as a substitute of ADO because of its superior metabolic stability. It is soluble in dimethyl sulfoxide (DMSO) at 25 mM and relatively insoluble in water. Although CADO stock solutions were prepared in DMSO, the main component was the water-based Krebs buffer solution. To compensate for this, we used 10 μM CADO, which is less than the maximum saturated concentration of 25 mM, and allowed sufficient reaction time. We allowed 1 h of reaction time after adding CADO in the Krebs buffer solution, which was also the reaction interval used in other experiments. The last limitation of our study arose from the fact that several types of receptors (e.g., mAChRs and nAChRs) are also responsible for presynaptic ACh release at the neuromuscular junction. However, mAChRs might also have some influence postsynaptically, although they occur in smooth muscles, developing muscles, and cultured muscles [27,28]. In contrast, the ACh receptor described so far in skeletal muscle is nicotinic [29]. The presynaptic mAChRs and nAChRs also modulate the spontaneous and evoked release of ACh. By modulating the spontaneous quantal release, presynaptic M1 and M2 receptors preserve and stabilize the synaptic function [30].
The overall function of ADO receptor at the neuromuscular presynaptic side is to decrease muscle tension after neuronal stimulation; this function might be disrupted when their balance of action altered. Presynaptic A1 receptors can modulate rocuronium-induced TOF fade following pirenzepine administration. Because the A1 receptor has an inhibitory effect on ACh release when activated, its agonistic effect from CADO might decrease the amount of ACh released by repeated evoked stimulation. This, in turn, affects TOFR in the rat hemidiaphragm generated by indirect supramaximal stimulation of the phrenic nerve and delays the recovery from neuromuscular blockade after sugammadex application. T1 twitch tension and TOFR are frequently used as surrogate markers of the degree of recovery from neuromuscular blockade.
 XML Download
XML Download