

 PDF
PDF Citation
Citation Print
Print



Introduction
Chemotherapeutic agents such as vincristine, paclitaxel, and oxaliplatin are widely used to treat several types of malignant tumors. However, these anti-cancer agents are also associated with peripheral neuropathic pain [1]. Chemotherapy-induced neuropathic pain is the most common side effect of cancer treatment, limiting the effectiveness of various anti-cancer agents and eventually impacting overall survival [2]. The paresthesia and dysesthesia induced by the vinca alkaloid vincristine occur in the early stage of vincristine treatment. Vincristine-induced peripheral neuropathy (VIPN) is often resistant to standard analgesics in humans; therefore, rodent models of VIPN have been developed to elucidate these pain mechanisms [3]. Pharmacological studies using these models have indicated that the mechanisms underlying allodynia and hyperalgesia after vincristine treatment are complex [4].
Adenosine is an endogenous purine compound, and it functions as an extracellular signaling molecule in the peripheral and central nervous systems [5]. Adenosine is locally released at tissue sites in response to ischemia, trauma, and interactions with specific receptors. There exists abundant experimental data demonstrating the role of adenosine in the modulation of nociceptive transmission at the spinal cord level [6]. To date, four types of adenosine receptors have been identified and cloned, namely A1, A2A, A2B, and A3 [5]. It is known that the antiallodynic and motor dysfunction effects of adenosine are mediated through the activation of spinal A1 and A2 adenosine receptors, respectively [7]. Activation of the adenosine A1 receptor has been shown to reduce allodynia in neuropathic pain animal models [7,8]. After spinal nerve ligation, intrathecal administration of adenosine A1 receptor agonists, including the R-(-) isomer of N6-(2-phenylisopropyl)-adenosine (R-PIA), is known to exert an antiallodynic effect mediated by the spinal adenosine A1 receptor system in rats [7,8]. Preconditioning with R-PIA has been shown to protect the brain and neuronal tissue against ischemic damage [9]. This protective mechanism, which is mediated via adenosine A1 receptor activation, activates a cascade of intracellular pathways, including adenosine triphosphate-sensitive potassium channel opening [9].
However, studies investigating the correlation between spinal adenosine A1 receptors and VIPN are limited. Therefore, the present study was designed to investigate the antinociceptive and anti-inflammatory effects of an intrathecally administered adenosine A1 receptor agonist, R-PIA, in a rat model of VIPN.
Materials and Methods
Experimental animals
A total of 50 male Sprague-Dawley rats (200–250 g, Orient, Seoul, Korea) were used across all experiments. Animals were housed in groups of 2–3 rats per cage in a room maintained at 22 ± 0.5°C with an alternating 12 h light-dark cycle. Food and water were available ad libitum. The animals were allowed to adapt to the laboratory environment for at least 2 h before testing. Experiments were performed during the light phase of the cycle (10 am–5 pm). All animal procedures and study protocols were approved by the Institutional Animal Care and Use Committee of the Asan Institute of Life Sciences, Seoul, Korea (IACUC Number: 2010-13-155).
Drugs and chemicals
Vincristine, the adenosine A1 receptor agonist R-PIA, and the adenosine A1 receptor antagonist 1,3-dipropyl-8-cyclopentylxanthine (DPCPX) were purchased from Sigma-Aldrich Inc., St. Louis, MO, USA. All drugs were dissolved in sterilized saline (0.9% sodium chloride solution).
Induction of peripheral neuropathy with vincristine
Vincristine sulfate (100 μg/kg/day) was intraperitoneally administered for a period of 10 days (two 5-day cycles with a 2-day pause in between) and peripheral neuropathy was induced [10]. Pain assessment and behavioral examination were conducted on days 0 (before vincristine administration), 14, and 28 (Fig. 1).
Behavioral examinations
Cold allodynia (acetone drop test)
Cold allodynia was measured using the acetone drop method at the hind paw as described by Choi et al. [11], with small modification. The rat was placed on top of a wire mesh grid, and access to the hind paws was allowed. Acetone (0.1 ml) was applied to the plantar surface of the left hind paw by using a pipette with polyethylene tube and the duration of paw withdrawal was recorded in seconds using a stopwatch. The minimum and maximum values were 0.1 s and 20 s, respectively. This was conducted three times, with an interval of 5 min between applications, and the total individual withdrawal durations were then calculated.
Mechanical hyperalgesia (pin-prick test)
Mechanical hyperalgesia of the paw was evaluated using the pin-prick test, as previously described by Erichsen and Blackburn-Munro [12]. The surface of the injured hind paw was touched with the point of a bent gauge needle (at a 90° angle to the syringe) at an intensity sufficient to produce a reflex withdrawal response. Paw withdrawal latency was then recorded in seconds using a stopwatch with a minimum value of 0.1 s.
Thermal hyperalgesia (hot plate test)
The thermal nociceptive threshold, employed as an index of thermal hyperalgesia, was assessed by the hot plate method, using a plate maintained at a temperature of 52.5 ± 1.0°C. The rat was placed on the hot plate, and the withdrawal latency with respect to licking of the hind paw was recorded in seconds. A cut-off time of 15 s was used [13].
Mechanical dynamic allodynia (paint brush test)
The ‘paint brush’ behavioral test is well-established for investigating dynamic responses to a mechanical stimulus. Normal rats never withdraw from a smooth paint brush, so a response to this stimulus is described as allodynia. The rat was placed in a cylinder with a wire mesh floor and a smooth paint brush was used to rub the plantar surface of the hind paw from the heel to the toes. This stimulus was applied five times, with intervals of 5 s, and the number of withdrawals was noted (between 0 and 5). The same procedure was repeated twice, with a gap of 5 min, and the number of withdrawals across the three tests was summed to obtain a single cumulative score of mechanical dynamic allodynia. The minimum and maximum values were 0 and 15, respectively [14].
Mechanical static allodynia (von Frey filament test)
For assessment of mechanical state allodynia, the rat was placed in an individual plastic cage with a wire mesh bottom. After 20 min, the mechanical threshold was measured by applying a series of eight calibrated von Frey filaments (0.4, 0.6, 1.0, 2.0, 4.0, 6.0, 8.0, and 15.0 g; Stoelting Co., USA) to the mid-plantar surface of the hind paw, ipsilateral to the nerve injury. This was conducted until a positive sign of pain behavior was elicited, at which point it was then held for 6 s. A brisk withdrawal or paw flinching was considered a positive response, in which case the next lower force filament was then applied. In the absence of such a response, the next greater force filament was then applied. In the absence of a response at 15.0 g of pressure, this cut-off value was assigned to the animal. The mechanical stimulus producing a 50% likelihood of withdrawal was determined using the up-down method [15].
Intrathecal administration of drugs
Drugs were injected in 10 µl volumes by a direct lumbar puncture method between the L5 and L6 vertebrae [16]. Briefly, rats were anesthetized with sevoflurane in oxygen delivered via a nose cone. The lumbar region was shaved and prepared with Betadine solution, and the intervertebral spaces were widened by placing the animal on a 50 ml syringe body. Then, animals were administered the test drug at the L5-6 interspace using a 0.5-inch 30-gauge needle (Becton-Dickinson, USA) connected to a Hamilton syringe (Microliter™ #702, Hamilton Co., USA). The Hamilton syringe was filled with 10–20 µl of the test drug. The needle plunger was then slowly pushed over a 30 s period, delivering a volume of 10 µl, and the needle was immediately removed. The exact subarachnoid positioning of the tip of the needle was monitored using a tail- or paw-flick test [16]. Then, the animals recovered in their home cage before behavioral testing was conducted. All pain treatment groups and drugs were tested in a randomized order. The experimenters were blind to the drug treatment groups.
Behavioral examinations on the 28th day (cold and mechanical static allodynia)
On the 28th day, after behavioral examination (cold allodynia, mechanical hyperalgesia, thermal hyperalgesia, mechanical dynamic allodynia, and mechanical static allodynia), the antinociceptive and anti-inflammatory effects of intrathecal R-PIA were estimated as follows. A pretreatment dose of saline (10 μl) or DPCPX (10 μg/10 μl) was intrathecally administered 15 min before intrathecal administration of R-PIA (0.3, 1.0, and 3.0 μg/10 μl). Measurements of cold and mechanical static allodynia were performed 15, 30, 45, 60, 90, 120, and 180 min after intrathecal R-PIA administration. Thirty minutes after the final measurements of cold and mechanical static allodynia (i.e., at 210 min), the animals were sacrificed. The sciatic nerve was then obtained, and biochemical estimations were conducted (Fig. 1). The doses of R-PIA and DPCPX administered in this study were chosen based on previous in vivo study involving rat models of neuropathic pain [17].
Biochemical estimations
Estimation of tumor necrosis factor-alpha level
After euthanasia, sciatic nerve samples were utilized for the determination of the tumor necrosis factor-alpha (TNF-α) level. TNF-α levels (sensitivity: 25 pg/ml) were determined using a rat TNF-α ELISA kit (R&D Systems, Inc., USA), following the manufacturer’s instructions. Testing of sciatic nerve homogenate samples was performed in duplicate. Recombinant anti-rat TNF-α was used to generate a standard curve (range: 0–20,000 pg/ml) as per the diagnostic kit. Absorbance was determined spectrophotometrically at 450 nm. The results were expressed as pg of TNF-α per mg of total protein in the supernatant.
Estimation of myeloperoxidase activity
Myeloperoxidase (MPO) activity was measured using a method described by Jain, Jaggi, and Singh [13]. After euthanasia, the sciatic nerve was obtained, and the nerve was minced and homogenized using a tissue homogenizer in an ice-cold potassium phosphate buffer (pH 7.4). The prepared homogenate was centrifuged at 5000 g for 10 min at 4°C. MPO activity in the supernatant was determined spectrophotometrically at 460 nm in the presence of hydrogen peroxide and 3,3,5,50-tetramethylbenzidine. The measured MPO values were expressed as MPO units (U)/mg of protein.
Experimental protocol
Eight groups were involved in the present study (Fig. 1).
Group I: normal control animals (n = 7)
The rats were subjected to no treatment and kept for 14 days. Behavioral tests were conducted on days 0, 7, 14, and 28. On the 28th day, after behavioral testing (cold and mechanical static allodynia), the animals were sacrificed. The sciatic nerve was then obtained and biochemical estimations were conducted.
Group II: vincristine-treated control animals (n = 7)
After induction of VIPN, behavioral examinations were conducted on days 0 (before treatment), 14, and 28. On the 28th day, a 10 μl pretreatment dose of saline was intrathecally administered 15 min before intrathecal administration of 10 μl saline. Measurements of cold and mechanical static allodynia were performed 15, 30, 45, 60, 90, 120, and 180 min after the intrathecal doses of saline. Thirty min after the final measurements of cold and mechanical static allodynia (i.e., at 210 min), the animals were sacrificed. The sciatic nerve was then obtained and biochemical estimations were conducted.
Groups III, IV, and V: R-PIA-administered (0.3 μg/10 μl, 1.0 μg/10 μl, and 3.0 μg/10 μl, intrathecal) vincristine-treated animals (n = 6 /each group)
After induction of VIPN, same behavioral examinations were conducted. On the 28th day, a 10 μl pretreatment dose of saline was intrathecally administered 15 min before intrathecal administration of R-PIA (0.3, 1.0, or 3.0 μg/10 μl). After measurements of cold and mechanical static allodynia, the animals were sacrificed, and the sciatic nerve was obtained for biochemical estimations.
Groups VI, VII, and VIII: DPCPX- (10 μg/10 μl) and R-PIA-administered (0.3, 1.0, or 3.0 μg/10 μl, intrathecal) vincristine-treated animals (n = 6 /each group)
All procedures were performed in the same serial order as the Groups III, IV, and V. However, a pretreatment dose of DPCPX (10 μg/10 μl) was intrathecally administered instead of saline.
Statistical analysis
Data are expressed as mean ± standard error of mean. Data from the behavioral tests were analyzed using the one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test, using SigmaPlot® Version 11 software (Systat Software Inc., USA). A P value < 0.05 was considered to be statistically significant.
Results
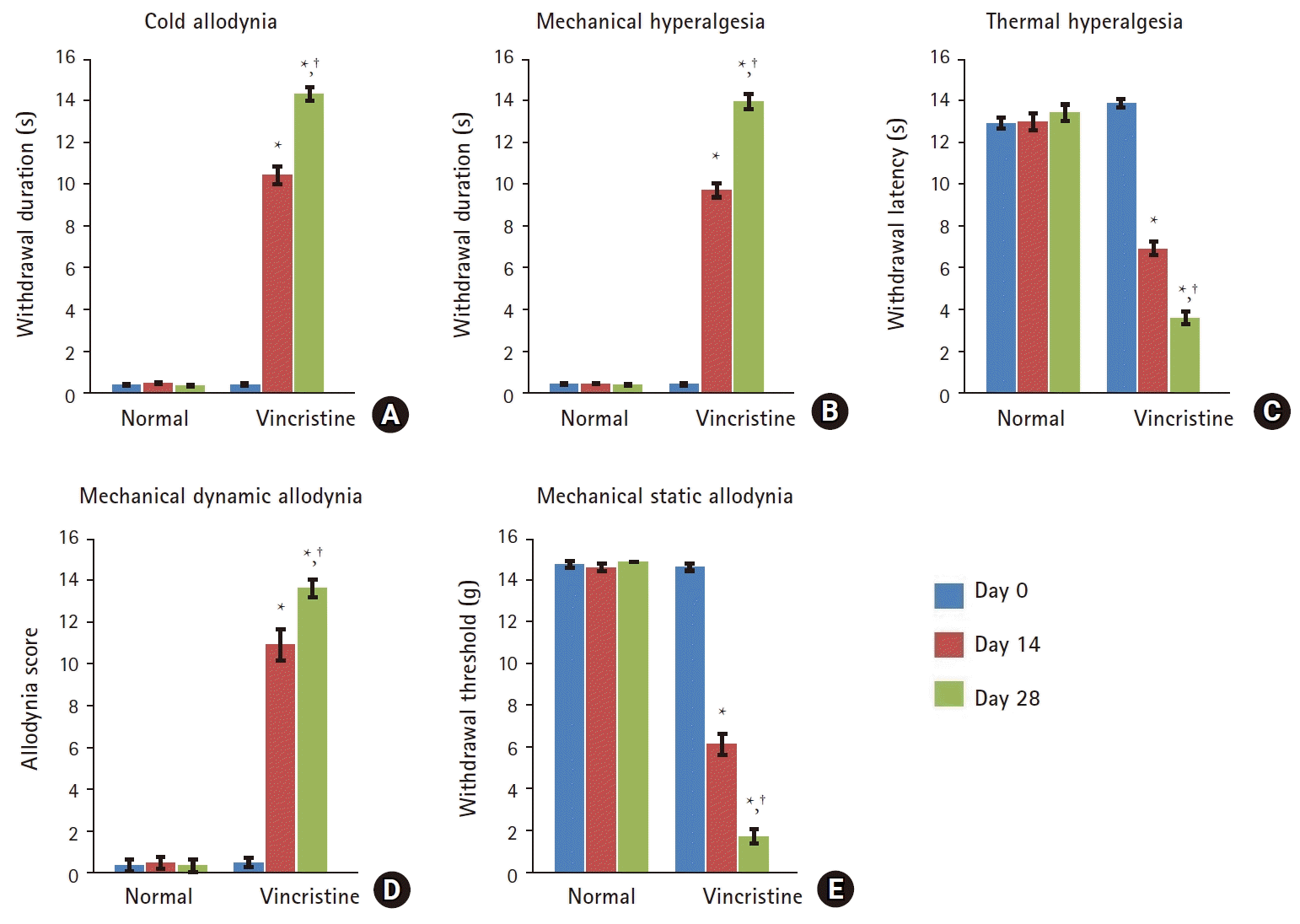
Effect of vincristine on hyperalgesia and allodynia
Compared with the normal group, administration of vincristine resulted in a significant increase in acetone- and pin prick-evoked paw withdrawal duration, indicating the development of cold allodynia (Fig. 2A) and mechanical hyperalgesia (Fig. 2B), respectively. Moreover, the withdrawal latency in the hot plate test was significantly decreased, indicating the development of thermal hyperalgesia (Fig. 2C). The allodynia score in the paint brush test was significantly increased, signifying the development of mechanical dynamic allodynia (Fig. 2D). In addition, the paw withdrawal threshold in response to von Frey filaments was significantly decreased, indicating the development of mechanical static allodynia (Fig. 2E).
Effect of intrathecal R-PIA administration on cold and mechanical static allodynia in vincristine-treated rats
Administration of vincristine resulted in a significant increase in acetone-evoked paw withdrawal duration (13.2 ± 0.4 s) compared with the normal group (0.5 ± 0.1 s) on the 28th day. Treatment with intrathecal R-PIA (1.0 and 3.0 μg/10 μl) resulted in a dose-dependent antiallodynic effect against cold allodynia. The antiallodynic effect was higher at the 3.0 μg/10 μl dose of R-PIA (0.7 ± 0.3 s) than at the 1.0 μg/10 μl dose (4.2 ± 0.7 s) (Table 1). Treatment with 0.3 μg/10 μl intrathecal R-PIA, however, did not result in an antiallodynic effect. This attenuating effect peaked 15 min after intrathecal administration of R-PIA (Table 1). After this time, the antiallodynic effect gradually decreased until 180 min. Pretreatment with DPCPX 15 min before intrathecal R-PIA administration significantly reversed the antiallodynic effect of R-PIA (11.4 ± 0.4 g and 11.1 ± 0.3 g for 1.0 and 3.0 μg/10 μl R-PIA, respectively). The data showing the effect of R-PIA administration on cold allodynia are presented in Table 1.
Administration of vincristine resulted in a significant decrease in the paw withdrawal threshold in response to von Frey filaments (1.8 ± 0.3 g), compared with the normal group (15.0 ± 0.0 g). Treatment with intrathecal R-PIA resulted in a dose-dependent antiallodynic effect against mechanical static allodynia. This antiallodynic effect was higher at the 3.0 μg/10 μl dose of R-PIA (14.5 ± 0.3 g) compared with the 1.0 μg/10 μl dose (10.8 ± 0.8 g) (Table 2). Treatment with 0.3 μg/10 μl intrathecal R-PIA did not result in an antiallodynic effect. This attenuating effect peaked 15 min after intrathecal administration of R-PIA (Table 2). After this time, the antiallodynic effect gradually decreased until 180 min. Pretreatment with DPCPX (10 μg/10 μl) 15 min before intrathecal R-PIA administration significantly reversed the antiallodynic effect of R-PIA (3.6 ± 0.4 g and 3.9 ± 0.3 g for R-PIA 1.0 and 3.0 μg/10 μl, respectively). The data showing the effect of R-PIA administration on mechanical static allodynia are presented in Table 2.
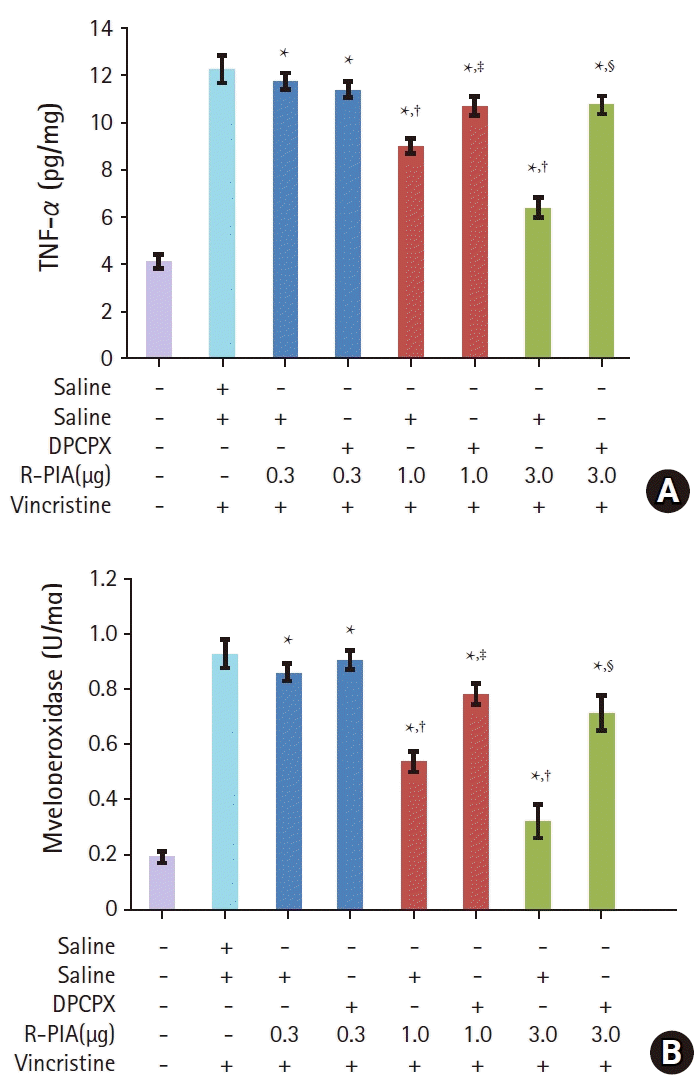
Effect of intrathecal R-PIA on TNF-α level and MPO activity in vincristine-treated rats
Administration of vincristine resulted in a significant increase in TNF-α level in tissue from the sciatic nerve (12.3 ± 0.6 pg/mg) compared with the normal group (4.1 ± 0.3 pg/mg). Intrathecal administration of R-PIA significantly reversed this increase in TNF-α level in a dose-dependent manner (9.0 ± 0.3 and 6.4 ± 0.4 pg/mg for R-PIA 1.0 and 3.0 μg/10 μl, respectively). Intrathecal administration of 0.3 μg/10 μl R-PIA did not affect the TNF-α level (Fig. 3A). Pretreatment with DPCPX before treatment with intrathecal R-PIA significantly reversed this attenuating effect of R-PIA (10.7 ± 0.4 and 10.7 ± 0.4 pg/mg for R-PIA 1.0 and 3.0 μg/10 μl, respectively) (Fig. 3A).
Administration of vincristine also resulted in a significant increase in MPO activity in tissue from the sciatic nerve (0.9 ± 0.1 U/mg) compared with the normal group (0.2 ± 0.0 U/mg). Intrathecal administration of R-PIA significantly reversed this increase in MPO activity in a dose-dependent manner (0.5 ± 0.0 and 0.3 ± 0.1 U/mg for R-PIA 1.0 and 3.0 μg/10 μl, respectively) (Fig. 3A). Intrathecal administration of 0.3 μg/10 μl R-PIA did not affect MPO activity (Fig. 3B). Pretreatment with DPCPX before treatment with intrathecal R-PIA significantly reversed this attenuating effect of R-PIA (0.8 ± 0.0 and 0.7 ± 0.1 U/mg for R-PIA 1.0 and 3.0 μg/10 μl, respectively) (Fig. 3B).
Discussion
Vincristine is a well-known chemotherapeutic agent widely used for the treatment of several malignancies such as breast cancer, leukemia, lymphoma, and primary brain tumors [18]. Nevertheless, sometimes neurotoxicity of the peripheral nerve fibers or sensory-motor neuropathy can develop during vincristine treatment. In the present study, intraperitoneal administration of vincristine (100 μg/kg) for 10 d caused the development of cold allodynia, mechanical dynamic and static allodynia, and mechanical and heat hyperalgesia, evaluated on the 14th and 28th day of the experiment. Mechanical dynamic allodynia is induced by light touching of the skin with a soft brush. Meanwhile, mechanical static allodynia is elicited by static pressure stimulation applied to the skin. Clinically, mechanical dynamic allodynia is a more common symptom than mechanical static allodynia. Therefore, the measurement of mechanical dynamic allodynia is preferred over mechanical static allodynia. It has been known that mechanical dynamic allodynia is mediated by peripheral low threshold, large myelinated Aβ-fibers [19]. The behavioral alterations observed in our study are consistent with those in previous studies demonstrating the development of pain symptoms following chronic vincristine administration [20]. Vincristine administration (0.1 mg/d) for 10 days (two 5-day cycles with a 2-day pause in between) was shown to produce persistent mechanical hyperalgesia and allodynia 5–8 days after peritoneal administration of vincristine, with this effect peaking after two weeks [10].
In the present investigation, treatment with intrathecal R-PIA after the development of vincristine-induced neuropathy significantly reversed behavioral changes, including paw cold allodynia and mechanical static allodynia. The antiallodynic effect of intrathecal R-PIA was significantly reversed by pretreatment with intrathecal DPCPX. Therefore, it is thought that the antiallodynic effect of intrathecal R-PIA may be mediated through adenosine A1 receptors in the spinal cord.
Since initial observations in the 1970s, numerous studies have demonstrated that systemic administration of adenosine A1 receptor agonists causes pain-reducing effects in animal models, such as inflammatory and neuropathic models exhibiting hyper-responsiveness. Additionally, the potential of adenosine A1 receptor agonists as therapeutic agents for pain has been considered in previous reports [6,21]. Subsequent preclinical studies have further illustrated the antinociceptive and/or antihyperalgesic actions of adenosine A1 receptor agonists in a diverse range of pain models. These models include the formalin model of inflammation, the carrageenan model of arthritis, hyperalgesia following surgical incision, the chronic constriction injury model of neuropathic pain, the spinal nerve ligation model of neuropathic pain, pain following spinal cord injury, and the streptozotocin model of diabetic neuropathy. This is an ongoing field of research, and reviews on the analgesic action of adenosine A1 receptor agents have been recently published [22].
Adenosine A1 receptors are located on peripheral sensory nerve endings [23], within the superficial layers of the dorsal horn of the spinal cord [24] and at specific supraspinal sites within the pain signaling neuraxis [25]. In addition to their neuronal localization, adenosine A1 receptors have also been identified on microglia [26], and their inhibition has been shown to contribute to antinociception in instances where the pain state involves glial activation and hypertrophy [26]. Adenosine A1 receptor signaling at the cellular level acts via the activation of an associated G protein; inhibition of cyclic AMP/PKA and interactions with Ca2+ and K+ channels are mediated via the Gαi subunit, and interactions with the PLC/IP3/DAG pathway via the Gα or βγ subunits. Additionally, adenosine A1 receptor signaling can involve β-arrestin-mediating receptor uncoupling and downregulation [27]. Several alternative clinical applications for adenosine A1 receptor agents in cardiovascular, respiratory, neuroprotective, and metabolic conditions are currently being explored [28].
Several studies have reported that intrathecal administration of adenosine A1 receptor agonists produces pain-reducing effects in animal models such as the spinal nerve ligation model of neuropathic pain and the formalin model of inflammation [18,29]. In the present study, intrathecally administered R-PIA at a dose of 1.0–3.0 μg/10 μl also produced an antiallodynic effect in a rat model of VIPN. Considering the dose of intraperitoneal R-PIA was 20–25 μg/kg (= 4.0–6.25 μg) [30,31] in previous studies, the intrathecal dose of R-PIA was only 25–48% of the intraperitoneal dose of R-PIA. A smaller dose is utilized when R-PIA is administered intrathecally; therefore, side effects are reduced. This is the first report suggesting the neuropathy-attenuating potential of intrathecal R-PIA in the VIPN model of rats.
In the present investigation, the anti-inflammatory effect of intrathecally administered R-PIA was also investigated. There exist a limited number of studies that have investigated the anti-inflammatory effects of R-PIA [32–34]. In one such study, inflammatory pain was induced in rats by an injection of Complete Freund’s Adjuvant (20 μl) into the plantar surfaces of hind paws. Electrophysiological recordings then suggested that the adenosine A1 receptor may potentiate glycinergic transmission through Gαi/PKA/α3 and Gβγ/α1ins pathways [32]. Furthermore, another study concluded that the cochlea expresses adenosine A1 receptors, which mediate tonic suppression of oxidative, inflammatory, and apoptotic processes. The protective role of adenosine against cisplatin ototoxicity was shown to be enhanced by inhibition of the NOX3/STAT1 signaling pathway [33]. Adenosine, lidocaine, and Mg2+ fluid therapy attenuated systemic inflammation, platelet dysfunction, and coagulopathy after non-compressible truncal hemorrhage in another study [34].
In the present study, we measured TNF-α levels in tissue surrounding the sciatic nerve as a biochemical surrogate marker of vincristine-induced inflammation. We found that vincristine treatment increased the level of TNF-α in this tissue. Inflammatory stimuli release reactive species such as NO• and •O2-, proinflammatory factors such as TNF-α, and pronociceptive mediators such as cytokines [35]. TNF-α is a major proinflammatory cytokine, and increased TNF-α levels are associated with pathological pain [36]. This vincristine-induced elevation in TNF-α level was markedly reduced with intrathecal administration of R-PIA (1.0 and 3.0 μg/10 μl). These data suggest that intrathecal R-PIA has anti-inflammatory effects. In the present study, the MPO enzyme level in tissue around the sciatic nerve was increased after administration of vincristine for a period of 10 days. MPO is released from neutrophils and is a significant marker of inflammation. Among the various mechanisms involved in vincristine-induced neuropathy, inflammatory reaction has been described to play a main role in the pathophysiology of nerve toxicity [37]. Long-term administration of vincristine activates spinal glial cells and up-regulates cytokines in these cells, leading to hyperalgesia and allodynia [38]. Vincristine damages Schwann cells, which causes demyelination [39], and damaged Schwann cells then mediate the release of inflammatory cytokines and chemokines including monocyte chemo-attractant protein-1 [39]. These series of processes are responsible for the infiltration of leucocytes, lymphocytes, and macrophages to the site of nerve injury, further augmenting the inflammatory environment and producing nerve damage [38]. In our study, the vincristine-induced increase in MPO activity was significantly reduced following intrathecal administration of R-PIA (1.0 and 3.0 μg/10 μl). The MPO data also suggested that intrathecal R-PIA has anti-inflammatory effects. Based on both the TNF-α and MPO data, the intrathecal R-PIA-mediated decrease in allodynia (both cold and mechanical static) observed in the VIPN model may be a result of its anti-inflammatory properties. This is the first report demonstrating the anti-inflammatory effect of intrathecal R-PIA in vincristine-induced neuropathic pain in rats. Therefore, given the existing literature and the data obtained from this study, it seems that intrathecal R-PIA exerts beneficial effects in a rat model of VIPN by virtue of its anti-inflammatory actions, specifically the inhibition of TNF-α levels and MPO activity. In the present study, there are some limitations such as the relationship between the antiallodynic and anti-inflammatory effect of intrathecal R-PIA that needs to be more fully defined. Further investigations are required to evaluate the close relationship between them.
In conclusion, the results of the present study suggest that intrathecally administered R-PIA attenuates neuropathic cold and mechanical static allodynia in the VIPN model of rats, partially due to its anti-inflammatory actions. Further studies are required to evaluate the involvement of the anti-inflammatory effects of R-PIA in VIPN.

 XML Download
XML Download