

 PDF
PDF Citation
Citation Print
Print



Introduction
Sugammadex is a prime antagonist of aminosteroidal neuromuscular blocking agents (NMBAs), especially rocuronium; However, before introducing sugammadex, it should be noted that the indirect mechanism of antagonism of the rocuronium-induced neuromuscular block has a ceiling effect, and is limited by the depth of the neuromuscular block at the time of reversal [1].
In the clinical setting, recovery of neuromuscular blockade is monitored through responses of the innervated muscle under indirect neuronal stimulation, particularly that of the ulnar nerve at the wrist level and the adductor pollicis muscle [2]. Patients with partial neuromuscular block usually show a train-of-four (TOF) ratio of < 0.7. TOF ratio of > 0.9 by mechanomyography or an electromyography-type monitor or > 1.0 by an acceleromyography-type monitor are considered as complete recovery [2]. However, these definitions were mostly established when neuromuscular blockade recovery was done by anticholinesterases. During blockade with nondepolarizing NMBAs, TOF fading occurs as a result of presynaptic cholinergic autoreceptor activity, which is influenced by concentrations of NMBAs and acetylcholine (ACh) at the neuromuscular junction [3]. Anticholinesterase administration was the main method of antagonism of neuromuscular blockade which cause an increase in ACh at neuromuscular junctions, which thereby outcompetes rocuronium at postsynaptic nicotinic acetylcholine receptors (nAChR) [4,5]. In contrast, sugammadex-induced antagonism is unlelated to ACh release or cholinergic activity [1]. Sugammadex directly encapsulates and inactivates rocuronium in a 1 : 1 ratio at the molecular level [6,7]. As such, we assumed that the TOF ratio recovery pattern by sugammadex might be different from those of anticholinesterase-induced recovery from neuromuscular blockade because sugammadex has no effect on ACh release or metabolism at the neuromuscular junction [1]. During sugammadex-induced neuromuscular recovery, the TOF ratio recovers to nearly 1.0 immediately after injecting sugammadex. However, patient complaints of muscle weakness have been reported even when extubation is performed after securing a TOF ratio of > 0.9. Indeed, it has been reported on several occasions that the TOF ratio recovery preceded twitch recovery in clinical settings during sugammadex-induced neuromuscular recovery [8–10].
As such, we hypothesized that at different doses of sugammadex for neuromuscular blockade reversal, rocuronium will be eliminated at different rates from neuromuscular junction, and the different affinities of rocuronium to the pre- and post-synaptic AChRs may affect recovery of both T1 and the TOF ratio, which may be hindered during anticholinesterase-induced recovery from neuromuscular blockade. The primary objective of this study was to assess recovery progressions of T1 and the TOF ratio after administrating different doses of sugammadex, and to compare the results obtained with those at spontaneous recovery. The secondary objective was to examine inter-group differences of TOF ratio recovery at the same T1 twitch tension during recovery from rocuronium-induced neuromuscular blockade.
Materials and Methods
Basic study design and sample preparation
This ex-vivo study protocol was approved by the Ethics Committee of the Laboratory of Animal Research of the Asan Institute of Life Science (Seoul, Korea) on July 1, 2017 (Protocol No. 2017-13-114). All animals were bred at a constant ambient temperature of 22°C under a regular diurnal cycle, and food and water were supplied ad libitum. The phrenic nerve-hemidiaphragm tissues were immersed in Krebs buffer solution (120 mM NaCl, 2.5 mM CaCl2, 4.7 mM KCl, 1.2 mM MgSO4, 1.2 mM KH2PO4, 25 mM NaHCO3, and 11 mM α-D-glucose) and maintained at 35°C with continuous bubbling of a mixture of 95% O2 and 5% CO2 to ensure tissue viability throughout the whole study. The sizes and weights of the tissues were measured and compared between groups (Table 1). In all experiments, sugammadex, rocuronium (Bridion® and Esmeron®, respectively; MSD Korea, Korea), and afaxalone (Alfaxan®; Careside Co. Ltd, Korea). were used.
Protocol for the main experiment
Sixty male Sprague-Dawley rats weighting an average of 354.8 ± 36.9 g (range 298.5–438.9 g) were used in the study. The rats were anesthetized with an intraperitoneal injection of 10 ml/kg of alfaxalone. The thoracic cages were immediately isolated and phrenic nerve-hemidiaphragm tissues were obtained. The tissues were fixed to a frame with electrodes and subsequently immersed in a 100 ml organ bath containing 75 ml of oxygenated Krebs buffer solution. For each specimen, the tendinous portion of the diaphragm was connected to a Grass FT03 Force Transducer (Grass Technologies, USA), and a resting tension of 40 mN was applied. The phrenic nerve was fixed to a platinum bipolar electrode and stimulated using a Grass S88 Stimulator (Grass Technologies, USA). Regardiing TOF simulation, supramaximal stimulation was delivered using a square wave pulse of 0.2 ms at 20-second intervals at 2 Hz for a total duration of 2 s. All waveforms were displayed and stored using the PowerLab 4/26 data acquisition system (AD Instruments, Australia) and Lab-Chart 7 software (AD Instruments, USA).
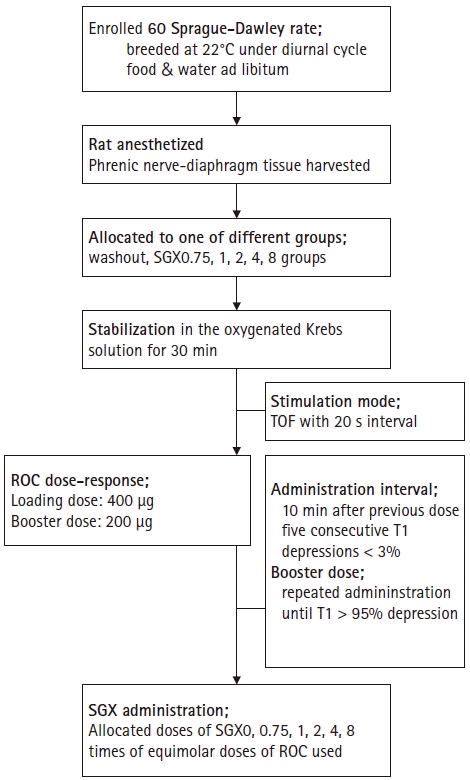
The phrenic nerve-hemidiaphragm tissues were randomly allocated to either a control group (washout) or one of five groups of different sugammadex doses (0.75, 1, 2, 4, or 8 times equimolar doses of rocuronium to produce > 95% T1 depressions; SGX0.75, SGX1 SGX2, SGX4, and SGX8, respectively) using random numbers generated by Microsoft Office Excel 2013 (Microsoft, USA). We sorted the groups into two categories: a high-dose group, with sugammadex at ≥ 2 times the equimolar dose of rocuronium; and a low-dose group, with sugammadex equal to or less than the equimolar dose of rocuronium. Twitch tensions and TOF ratio were serially monitored during a 30 min-stabilization time. After the stabilization period, 400 µg rocuronium was added to the organ bath. Subsequently, 200 μg booster doses of rocuronium were added when the five consecutive T1 depressions were either ≤ 3% of the previous T1 twitch tension, or 10 minutes after the previous dose. Booster dosing was stopped when T1 depression of ≥ 95% was achieved. The loading dose was set as the amount that produced no change in T1 twitch tension but changed the TOF ratio within 3% of that before the loading dose. Booster doses were set as the level of the first booster that produced a change of the T1 twitch tension, and the total numbers of boosters administerd was ≤ 10. The study protocol is summarized in Fig. 1.
The concentration of rocuronium required to obtain a reduction of T1 of > 95% was noted in the control and all sugammadex groups. For the comparisons, rocuronium dose-responses were plotted, regression curves obtained, and group-wise comparisons of the values performed. TOF ratios were obtained while monitoring T1 depression, and the regression curves were compared among the groups. Following that, inter-group progressions of the TOF ratios by % recovery of the T1 at each different sugammadex dose were compared among the groups.
Statistical analysis
The sample size was calculated based on the previous experiment and pilot study, which suggested that 10 samples per group were sufficient at α = 0.05, power = 0.80, and a dropout rate of 10%. Results are expressed as mean ± SD. All doses are expressed as μM. Statistical analysis was carried out using SPSS ver. 13.0 software (IBM Corp., USA). Recovery data were plotted by fitting nonlinear regression curves to the group data. An equation model was selected when the R2 > 0.8 by using curve estimation in SPSS. To describe recovery of T1 and the TOF ratio, the following equation was used: y = Ωx + b; where y represents TOF ratio progression, x represents T1 recovery, and Ω represents the slope of the regression curve (R2 = 0.87). For simultaneous group-wise comparison of recovery progression of T1 and the TOF ratio by time, a variable (TOFR/T1 product; which contained data of TOF ratio over T1 at specific time-point) was calculated using the following equations: y = λx, or y = λ*1/x, where y and x represent the TOFR/T1 product and time, respectively, while λ represents the slope of the regression curve (R2 = 0.91, 0.83 in the low- and high-dose groups, respectively). Different Ω and λ values correspond to the speed of T1 recovery to > 95%, TOF ratio to > 0.9, or speed of increment of decay of the TOFR/T1 product. The mean group values of Ω and λ were compared using the Mann Whitney U test. Statistical significance was accepted at P values of < 0.05.
Results
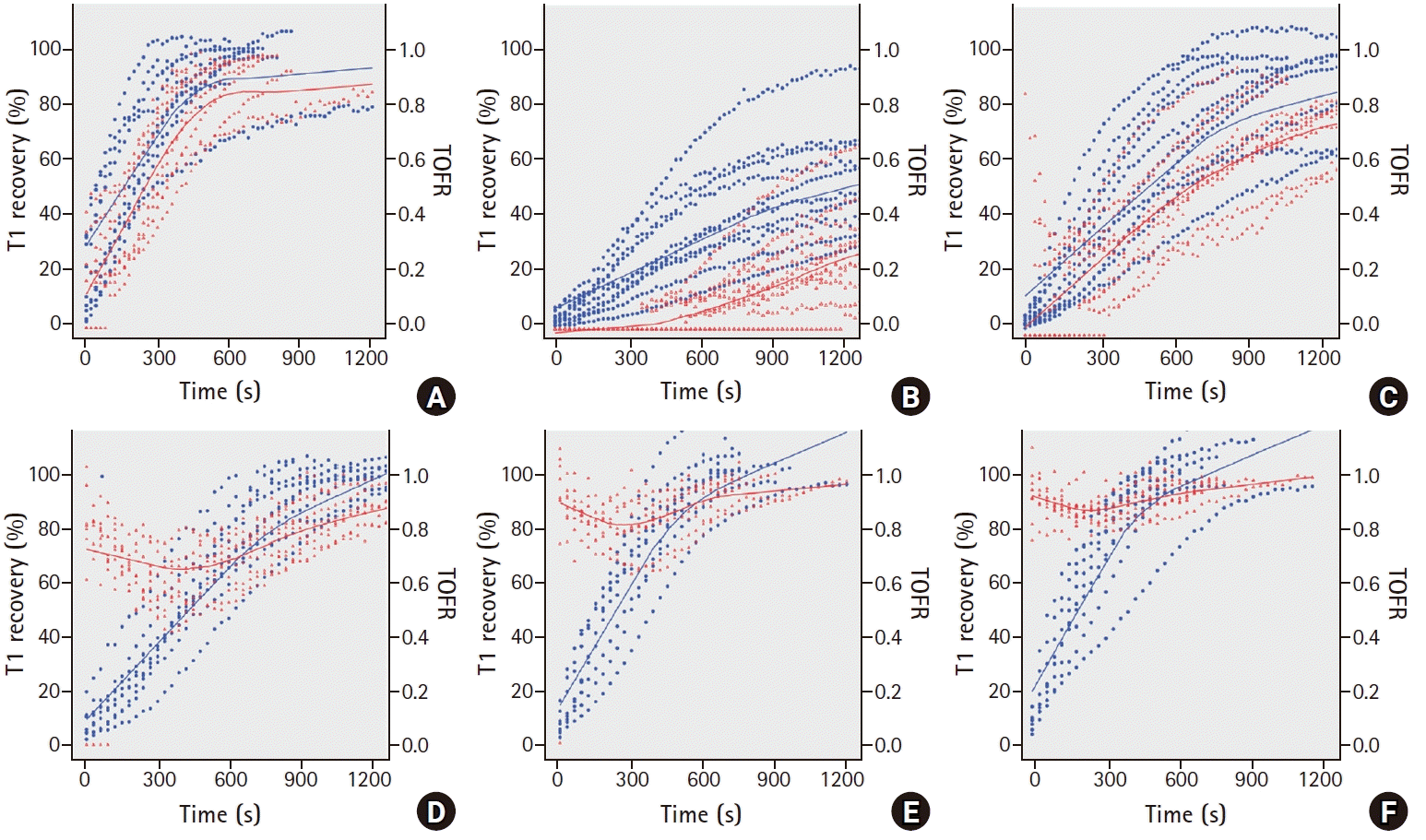
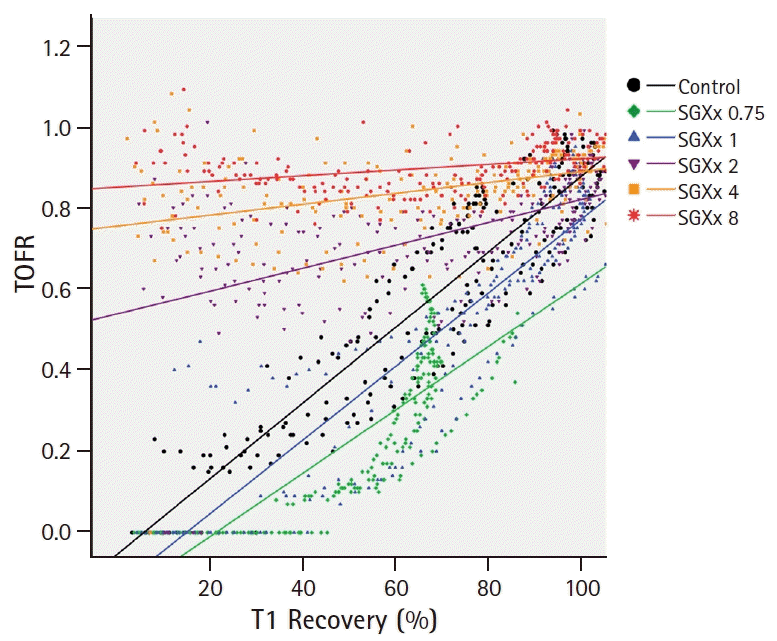
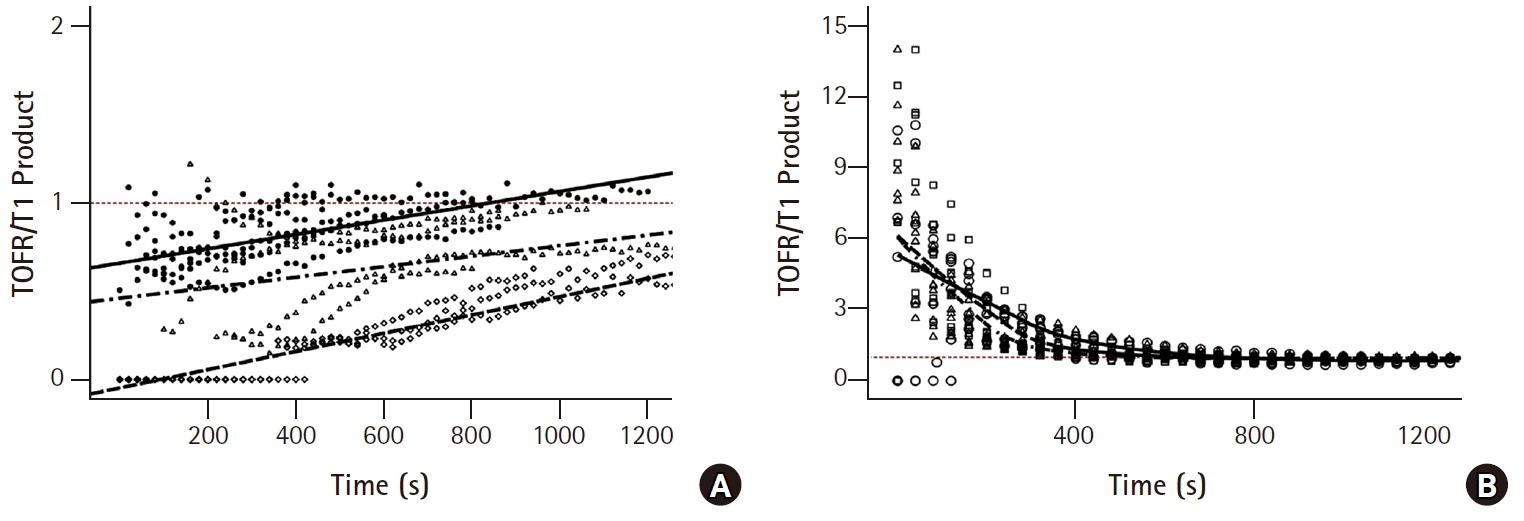
The specimen sizes and weights were similar in the six groups (Table 1). In the control, SGX0.75, and SGX1 groups, reappearance of T1 before the TOF ratio (Figs. 2A, 2B, and 2C) and of T2, T3, and T4 was observed. However, in the SGX2, SGX4, and SGX8 groups, simultaneous reappearance of T1 and T4 was observed and a TOF ratio of ≥ 0.7 was obtained from the start of recovery (Figs. 2D, 2E, and 2F). Inter-group comparison was conducted on the TOF ratio progression by T1 recovery. Based on the results in Fig. 2, the slopes (Ω) of the control, SGX0.75, and SGX1 groups (Fig. 3A) were steeper than those of the SGX2, SGX4, and SGX8 groups (Fig. 3B). The combined T1 recovery and TOF ratio was expressed as a single variable (TOFR/T1 product), which was plotted against time (Fig. 4). In the control, SGX0.75, and SGX1 groups, regression curves were fitted using the following equation: y = λx; where y represents the TOFR/T1, x represents the recovery time with 5% T1 recovery as the zero point, and λ represents the slope (Fig. 4A). In these groups, no statistically significant differences in λ were observed. In the SGX2, SGX4, and SGX8 groups, the regression curves were fitted using the following equation: y = λ*1/x, where the variables represent the same parameters as those for the equation, y = λx (Fig. 4B).
Discussion
This ex-vivo experiment demonstrates that the recovery patterns of the TOF ratio of the SGX2, SGX4 and SGX8 groups were significantly different from the control, SGX0.75, and SGX1 groups.
During onset and recovery of neuromuscular blockade, the action of NMBAs is determined by competitive binding of ACh and NMBAs to the postsynaptic nAChRs at the neuromuscular junctions [11]. The conventional strategy for neuromuscular blockade reversal involves the use of pharmacological tools to increase ACh levels, thereby increasing binding at postsynaptic nAChRs by inhibiting acetylcholinesterase near postsynaptic nAChRs. In contrast, sugammadex binds and chelates the ‘guest’ molecules at a 1 : 1 ratio; thus, sugammadex has no effects on the release and levels of ACh at the neuromuscular junctions, while reversing neuromuscular blockade [1]. Several receptors mediate and modulate the release of ACh at neuromuscular junctions. In general, ACh release in variable neuronal signal freuqencies is modulated through fine tuning of several receptors at the presynaptic membrane by ACh and adenosine [12]. The TOF ratio is the most common and valuable measurement during neuromuscular monitoring, which is performed by four stimuli of 2 Hz [2]. With this low-frequency stimulation, presynaptic muscarinic M1AChRs and adenosine A1 receptors are predominant and modulate the release of adequate amounts of ACh [12,13]. ACh then transmits the signal to the post synaptic junction via postsynaptic nAChRs, which affects the presynaptic neuronal nAChRs, which in turn have a positive feedback on ACh release. During non-depolarizing NMBA-induced neuromuscular blockade, the TOF fade and TOF ratio are both considered phenomena related to presynaptic neuronal nAChRs [3,14,15]. A recent study reported that these phenomena are related to the postsynaptic receptor type [15]. Those authors conducted an in-vivo experiment; the TOF fade occurred only under blockade of the postjunctional nAChRs with α-bungarotoxin (α-BTX) or α-conotoxin, but not under that of the presynaptic nAChR with specific blockers alone. Moreover, co-administration of α-BTX or α-conotoxin during presynaptic nAChR blockade resulted in a prominent TOF fade. Other studies reported that the presynaptic nAChR regulates the amount of ACh per neural stimulus [15,16]. Faria et al. [15] reported that blockade with dihydro-β-erythrodine (DhβE) was effective in decreasing the level of prejunctional ACh release, which was consistent with the findings from another study using a cell culture model. Decrease of ACh release causes the onset of a neuromuscular blockade, which was slow under blockade of only the postjunctional nAChRs, but accelerated under that of prejunctional AChRs with DhβE [17]. Therefore, the postsynaptic and presynaptic actions of NMBAs on nAChRs affect the efficacy of neuromuscular blockade [18]. Considering these results, the relationship between pre- and postsynaptic receptor function is an important determinant of the TOF fade and the TOF ratio. The conventional strategy for neuromuscular block reversal with anticholinesterase administration is not capable of eliminating NMBAs at the neuromuscular junction, leading to prolonged neuromuscular blocking activities after the initial administration of anticholinesterase. The TOF fade and the TOF ratio are prominent for the duration of NMBA action at the postsynaptic nAChRs. Sugammadex is not able to inactivate neuromuscular blockade at the neuromuscular junction [1,7], but causes immediate reduction of the NMBA concentration outside the neuromuscular junction. This causes rapid transfer of NMBAs, due to the concentration gradient across the neuromuscular junction [1]. Thus, reduction of NMBA concentrations and their activities at the neuromuscular junction attenuates the TOF fade and increases the TOF ratio, which explains the results of our ex-vivo experiment. We sorted the six groups into two categories to enable clear description and easy understanding for readers: a high-dose group, with sugammadex at ≥ 2 times the equimolar dose of rocuronium; and a low-dose group, with sugammadex at the level of or less than the equimolar dose of rocuronium. In the high-dose groups, a rapid drop of rocuronium concentration in the Krebs buffer solution allowed rapid exit of rocuronium molecules from the neuromuscular junction, and thereby, achieved attenuation and disappearance of TOF fade even in the early recovery period. In the low-dose group, rocuronium molecules remained at the neuromuscular junction under sugammadex, and the TOF fade and the TOF ratio were maintained until rocuronium, was adequately decreased, due to the concentration gradient across the neuromuscular junction. We obtained similar results in the low-dose groups to that of spontaneous recovery. In Fig. 3A, as the TOF ratio recovered in parallel with T1 recovery and showed prominent TOF fade in the low-dose group, y-axis values (TOF ratio) often start low and converge at 1. In contrast, TOF fade was attenuated and the TOF ratio was high even in the early recovery of T1 in the high-dose group. As such, the slopes of regression curves, Ω, of low-dose groups were steeper than those of high-dose groups. In Fig. 4, we demonstrated the simultaneous progression of % T1 recovery and the TOF ratio by time. We generated one value (TOFR/T1 product) by using T1 recovery and the TOF ratio, which were converted by % value. In the low dose group, as the T1 recovery preceded the TOF ratio, TOFR/T1 was ≤ 1. As such, it was well-represented by the equation of y = λx. In contrast, those in the high dose group were ≥ 1 and showed a decay pattern, because TOF fade was attenuated and the TOF ratio was higher than T1 recovery.
Our study has several limitations. First, we conducted an ex-vivo experiment and disregarded the pharmacokinetic component of rocuronium action, since the phrenic nerve-hemidiaphragm tissue specimens were examined in an organ bath filled with Krebs buffer solution. The overall recovery time to > 95% T1 was > 30 minutes in the low-dose group and was shortened to < 15 minutes in the high-dose group. In clinical settings, however, the recovery time is ≤ 5 minutes [19,20] considering the dose of sugammadex used. Reports have indicated that administration of 2 mg/kg sugammadex for moderate neuromuscular blockade and 4 mg/kg for deep neuromuscular blockade achieved recovery times of ≤ 3 and 5 minutes to a TOF ratio of > 0.9, respectively [20–22]. The discrepancies of recovery patterns between the in-vivo and ex-vivo approaches suggest that the results should be interpreted differently considering that NMBA-induced neuromuscular blockade is fully and rapidly recovered in clinical settings, which might hinder our results make the blockade disappeared without notice. As such, although the postoperative residual block is still a problem, even in the new era of sugammadex-induced recovery from a neuromuscular block [23,24], we should cautiously judge the clinical implications of the findings of the current ex-vivo study. Second, this study focused on the nicotinic AChR subtype at the presynaptic and postsynaptic junctions. Sugammadex has no action at the neuromuscular junction, and rocuronium has no action on the other receptors at the neuromuscular junction; this study only focused on the nAChRs, which are the primary action site of ACh and NMBAs during neuromuscular blockade. However, as we described above, several receptors modulate ACh release in different environments [12,13], and we tried to maintain a consistent environment and neural stimulation throughout the study period. We used TOF stimulation of four stimuli of 2 Hz supramaximal stimulation, which is the same mode used for clinical neuromuscular monitoring. The method of obtaining the phrenic nerve-hemidiaphragm tissue specimen had the disadvantage of temporary hypoxia and damage to the tissue specimen during preparation due to thorax extraction from the rat. To minimize these drawbacks, we made attempts to oxygenate the rat, to remove the thorax immediately after the aorta was cut, and then performed trimming of the specimen in a petri dish containing Krebs buffer solution aerated with a mixture of 95% O2 and 5% CO2.
In conclusion, a high dose of sugammadex rapidly reversed the neuromuscular block induced by rocuronium. However, the recovery pattern of the TOF ratio differ according to the dose of sugammadex, particularly when the amount of sugammadex is high enough compared to that of rocuronium used. In that condition, a high TOF ratio may be achieved even without full recovery of the T1 twitch tension. The TOF ratio alone might be insufficient to indicate full recovery of neuromuscular blockade without full recovery of the T1 twitch. Therefore, clinicians should use an appropriate dose of sugammadex and wait for full recovery of both the TOF ratio and T1 twitch.
 XML Download
XML Download