

 PDF
PDF Citation
Citation Print
Print


Introduction
Indigo carmine, a blue dye that is concentrated by the kidney and excreted primarily in the urine, is used as a marker dye in localizing ureteral orifices, severed ureters and fistulous communication [1]. The intravenous administration of indigo carmine during anesthesia has been reported to produce transiently increased blood pressure in patients [2-5]. The commonly encountered cardiovascular effect of intravenously administered indigo carmine during anesthesia might be associated with transient alpha-adrenoceptor stimulation, which leads to increased total peripheral resistance and hypertension [4,5]. Indigo carmine inhibits endothelium-dependent nitric oxide (NO)-mediated relaxation and produces superoxide radicals [6,7]. However, the detailed effect of indigo carmine on a phenylephrine-induced contraction in an isolated vessel has not been investigated. Therefore, the goals of this in vitro study were to examine the effects of indigo carmine on the phenylephrine-induced contraction in the isolated rat aorta and to determine the associated cellular mechanism with particular focus on the endothelium-derived vasodilators (NO and prostacyclin) [8]. Based on previous reports [6-8], this study tested the hypothesis that indigo carmine would increase the phenylephrine-induced contraction in endothelium-intact aorta via the inactivation of NO due to the increased production of reactive oxygen species (ROS) including the superoxide radical.
Go to : 
Materials and Methods
All experimental procedures and protocols were approved by the Institutional Animal Care and Use Committee of Gyeongsang National University, and were performed in accordance with the Guide for the Care and Use of Laboratory Animals prepared by the National Academy of Sciences.
Experimental preparation
Male Sprague-Dawley rats (n = 37) weighing 250-350 g each were anesthetized by the intraperitoneal administration of pentobarbital sodium (50 mg/kg). The descending thoracic aorta was dissected free, and the surrounding connective tissue and fat were removed under microscopic guidance while the blood vessels were bathed in Krebs solution with the following composition: 118 mM NaCl, 4.7 mM KCl, 1.2 mM MgSO4, 1.2 mM KH2PO4, 2.4 mM CaCl2, 25 mM NaHCO3, 11 mM glucose and 0.03 mM EDTA. The aorta was then cut into 2.5 mm rings, which were suspended on Grass isometric transducers (FT-03, Grass Instrument, Quincy, MA, USA) with a resting tension of 4.0 g in 10 ml temperature-controlled baths (37℃) containing Krebs solution that had been gassed continuously with 95% O2 and 5% CO2. The rings were equilibrated at a resting tension of 4.0 g for 120 min, during which time the bathing solution was changed every 30 min. In some aortic rings, the endothelium was removed intentionally by inserting a 25-gauge needle tip into the lumen of the rings and rubbing the ring gently for a few seconds. Only one concentration-response curve elicited by phenylephrine was made for each ring in the experiments. The contractile response induced by isotonic 60 mM KCl was measured in all aortic rings. Once the isotonic 60 mM KCl-induced contraction had stabilized, acetylcholine (10-5 M) was added to assess the integrity of the endothelium. The endothelial integrity was confirmed by an observation of > 25% relaxation by acetylcholine (10-5 M) from the 60 mM KCl-induced precontraction [9]. The endothelium-denuded aortic rings failed to respond to acetylcholine (10-5 M). In the preliminary experiments, in terms of the magnitude, the > 25% relaxation induced by 10-5 M acetylcholine in the rings precontracted with 60 mM KCl was similar to the > 80% relaxation response to 10-5 M acetylcholine in the rings precontracted with 10-7 M phenylephrine [9].
Study design
The first series of these experiments examined the effect of indigo carmine on the contractile response induced by the alpha-1 adrenoceptor agonist, phenylephrine, in the endothelium-intact and -denuded rings. Indigo carmine was added directly to the organ bath 15 min before the addition of phenylephrine. The effect of indigo carmine on the concentration-response curves for phenylephrine (10-9 to 10-5 M) was assessed by comparing the contractile response in the presence or absence of indigo carmine (10-5 M).
The second series of experiments investigated the involvement of the endothelium-derived vasodilators (NO and prostacyclin) in the indigo carmine-induced potentiation of the contractile response induced by phenylephrine. The effect of indigo carmine on the concentration-response curve for phenylephrine in the endothelium-intact rings pretreated independently with the nitric oxide synthase (NOS) inhibitor, Nω-nitro-L-arginine methyl ester (L-NAME: 10-4 M), and cyclooxygenase inhibitor, indomethacin (10-5 M), was assessed by comparing the contractile response in the presence or absence of indigo carmine (10-5 M). There was a 20 min incubation period for each inhibitor plus indigo carmine or each inhibitor alone before the phenylephrine-induced contraction.
The third series of experiments evaluated the participation of ROS including superoxide radicals in the indigo carmine-induced potentiation of the contractile response induced by phenylephrine. The effect of indigo carmine on the concentration-response curve for phenylephrine in the endothelium-intact rings pretreated with a low-molecular-weight superoxide anion scavenger, 4,5-dihydroxy-1,3-benzene-disulfonic acid disodium salt monohydrate (tiron: 10-2 M) was examined by comparing the contractile response in the presence or absence of indigo carmine (10-5 M). There was a 20 min incubation period for tiron plus indigo carmine or tiron alone.
Finally, the participation of NO in the indigo carmine-induced potentiation of the contractile response induced by phenylephrine in the endothelium-intact rings was studied. Indigo carmine was added directly to the organ bath 40 min before the phenylephrine (10-5 M)-induced maximum contraction in the endothelium-intact rings. L-NAME (10-4 M) was added directly to the organ bath after the phenylephrine-induced maximum contraction reached stable tension in the presence or absence of indigo carmine (10-5 M). The effect of indigo carmine on the L-NAME-induced contraction in the endothelium-intact rings with the stable maximal contractile state induced by phenylephrine (10-5 M) was assessed by comparing the L-NAME-induced contractile response in the presence or absence of indigo carmine (10-5 M).
Cell culture
Rat aortic vascular smooth muscle cells (VSMC) were isolated from free-floating explants of the aorta, as described previously [10]. Briefly, the thoracic aortae were dissected from adult male Sprague-Dawley rats, cut longitudinally and the endothelial cells were removed. The isolated medial membrane was cut into small pieces and incubated for 1 day. After supplementation with fresh medium, the tissue was incubated for a few days. The VSMCs were removed by trypsinization and maintained in Dulbecco's modified Eagle's medium (DMEM) containing 100 U/ml penicillin and 100 µg/ml streptomycin, supplemented with 10% heat-inactivated fetal bovine serum (FBS), at 37℃ in an atmosphere of 95% air and 5% CO2. The cells from passages 4 to 10 were used for the experiments.
Measurement of ROS level
The production of intracellular ROS was monitored spectrofluorometrically using dichlorodihydrofluorescein diacetate as a fluorescent dye. The cells were trypsinized 12 h after serum deprivation, and the cell suspension was treated with dichlorodihydrofluorescein diacetate at a final concentration of 10-5 M in the medium for 30 min. Oxidation of dichlorodihydrofluorescein by ROS yields dichlorofluorescein (DCF). The fluorescence of DCF was monitored at excitation and emission wavelengths of 485 and 530 nm, respectively, using a fluorescence plate reader (20 cycles per 30 s at 37℃) (Tecan®, Tecan US Inc., USA). The data is expressed as the relative changes to the initial fluorescence.
Reagents
All the drugs were of the highest purity commercially available: phenylephrine HCl, L-NAME, indomethacin, tiron, 2', 7'-dichlorodihydrofluorescein diacetate (Singma Chemical, St. Louis, MO, USA), indigo carmine (Korea United Pharm. INC, Seoul, Korea). DMEM, FBS, penicillin and streptomycin were supplied by Gibco-BRL (Rockville, MD, USA). All drug concentrations are expressed as the final molar concentration in the organ bath. Indomethacin was dissolved in dimethyl sulfoxide (final organ bath concentration: 0.1%). Unless stated otherwise, all other drugs were dissolved and diluted in distilled water.
Data analysis
The values are expressed as the mean ± SD. The contractile responses to phenylephrine are expressed as the percentage of their own relevant maximum contraction to isotonic 60 mM KCl. The logarithm of the drug concentration (ED50) eliciting 50% of the maximum contractile response was calculated using nonlinear regression analysis by fitting the concentration-response relationship for phenylephrine to a sigmoidal curve using commercially available software (Prism version 5.0 for windows, GraphPad Software, San Diego, CA, USA). The data was fitted to a sigmoid dose-response curve using the following equation Y = Bottom + (Top - Bottom)/(1 + 10^ ((LogED50 - X) × Hill Slope)). The phenylephrine-induced maximum contractions were calculated as the percentage of their own maximum contraction to isotonic 60 mM KCl. The L-NAME (10-4 M)-induced maximum contraction was calculated as the percentage change from the phenylephrine (10-5 M)-induced maximum contraction in the presence or absence of indigo carmine (10-5 M). Statistical analysis for comparing the ED50 and maximum contraction between the no-drug and indigo carmine-treated groups was performed using a Student's t-test. The effect of indigo carmine on the phenylephrine-induced contractions at the same concentration was analyzed by two-way analysis of the variance with a Bonferroni posttest (Prism version 5.0 for windows). P values < 0.05 were considered significant. N refers to the number of thoracic aortic rings used in each experiment.
Go to :
Results
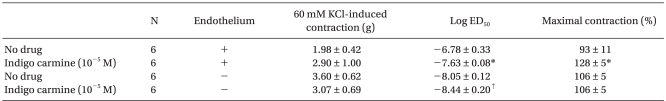
In the endothelium-intact rings, indigo carmine (10-5 M) produced a significant leftward shift (ED50: P < 0.001) in the phenylephrine concentration-response curves, and increased (P < 0.001) the phenylephrine-induced maximum contraction compared with the rings not treated with indigo carmine (Fig. 1A, Table 1). However, in the endothelium-denuded rings, indigo carmine (10-5 M) had no effect on the phenylephrine-induced maximum contraction and produced a slightly significant leftward shift (ED50: P < 0.01) in the phenylephrine concentration-response curve (Fig. 1B, Table 1).
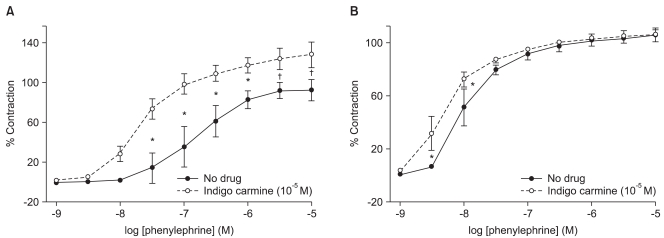 | Fig. 1Effect of indigo carmine on the phenylephrine concentration-response curves in the endothelium-intact (A) and -denuded (B) rings. The data is shown as the mean ± SD, and is expressed as the percentage of the maximum contraction induced by isotonic 60 mM KCl. (A) *P < 0.001 versus no drug at 3 × 10-8 to 10-6 M phenylephrine, †P < 0.01 versus no drug at 3 × 10-6 and 10-5 M phenylephrine. (B) *P < 0.001 versus no drug at 3 × 10-9 and 10-8 M phenylephrine.
|
In the endothelium-intact rings pretreated with 10-5 M indomethacin, indigo carmine (10-5 M) produced a significant leftward shift (ED50: P < 0.001) in the phenylephrine concentration-response curves, and increased (P < 0.01) the phenylephrine-induced maximum contraction compared with the rings not treated with indigo carmine (Fig. 2A, Table 2). However, indigo carmine had no effect on the phenylephrine-induced maximum contraction in the endothelium-intact rings pretreated with 10-4 M L-NAME and produced a slightly significant leftward shift (ED50: P < 0.05) in the phenylephrine concentration-response curve (Fig. 2B, Table 2).
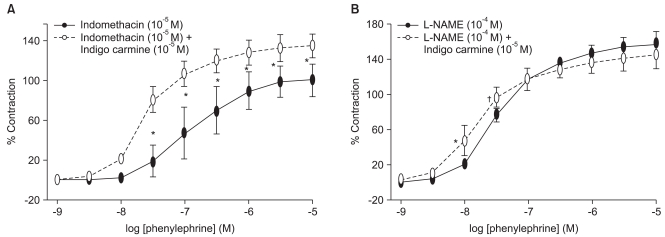 | Fig. 2Effect of indigo carmine on the phenylephrine concentration-response curves in the endothelium-intact rings pretreated with 10-5 M indomethacin (A) and 10-4 M Nω-nitro-L-arginine methyl ester (L-NAME) (B). The data is shown as the mean ± SD, and is expressed as the percentage of the maximum contraction induced by isotonic 60 mM KCl. (A) *P < 0.001 versus no drug at 3 × 10-8 to 10-5 M phenylephrine. (B) *P < 0.001 versus no drug at 10-8 M phenylephrine, †P < 0.05 versus no drug at 3 × 10-8 M phenylephrine.
|
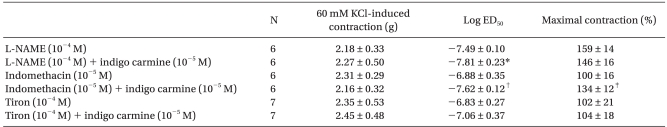
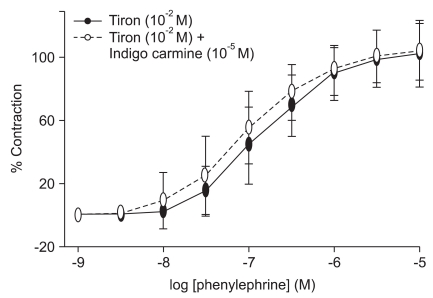
In the endothelium-intact rings treated with the low-molecular-weight superoxide anion scavenger, tiron (10-2 M), indigo carmine (10-5 M) did not alter phenylephrine-induced contraction significantly (Fig. 3, Table 2).
In the endothelium-intact rings that developed a stable phenylephrine (10-5 M)-induced maximum contraction in the presence or absence of indigo carmine, the L-NAME (10-4 M)-induced contractile response (no drug = 73 ± 38%, 10-5 M indigo carmine = 9 ± 6%) was attenuated (P < 0.001) in the endothelium-intact rings pretreated with indigo carmine compared with the rings without 10-5 M indigo carmine (Fig. 4).
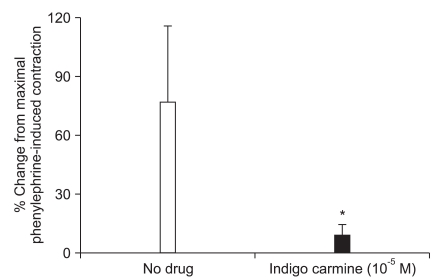 | Fig. 4After the phenylephrine (10-5 M)-induced maximum contraction in the endothelium-intact rings reached a stable tension in the absence or presence of indigo carmine (10-5 M), 10-4 M Nω-nitro-L-arginine methyl ester (L-NAME) was added directly to the organ baths. The L-NAME-induced contraction was assessed in the absence or presence of indigo carmine (10-5 M). The data is shown as the mean ± SD, and is expressed as the percentage change from the maximum contraction induced by 10-5 M phenylephrine (10-5 M phenylephrine-induced maximum contraction: 1.87 ± 0.49 g [N = 9] and 3.42 ± 1.17 g [N = 9] for the rings not treated with indigo carmine and the indigo carmine [10-5 M] pretreated rings, respectively). *P < 0.001 versus no drug.
|
Indigo carmine (10-5 M) increased the fluorescence of oxidized DCF (P < 0.01), whereas tiron completely abolished the indigo carmine-induced increase in the fluorescence of oxidized DCF (P < 0.05) (Fig. 5).
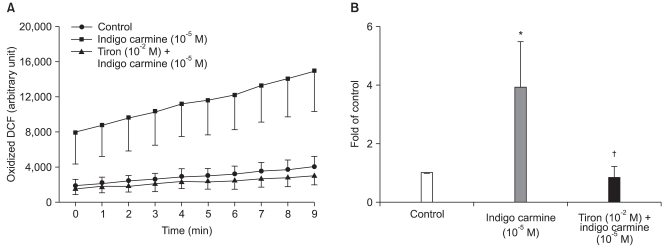 | Fig. 5Effect of indigo carmine (10-5 M) with or without tiron (10-2 M) on the fluorescence of oxidized dichlorofluorescein (DCF) in rat aortic vascular smooth muscle cells. The cells were incubated with indigo carmine for 10 min after pretreatment with tiron for 30 min. The cells were then loaded with 10-5 M dichlorodihydrofluorescein diacetate for 30 min. The data is shown as the mean ± SD (n = 4) and expressed as the relative changes to the initial fluorescence (A) or fold of the control at 9 min (B). *P < 0.01 versus the control. †P < 0.05 versus indigo carmine (10-5 M).
|
Go to :
Discussion
This report presents new information suggesting that indigo carmine (10-5 M) increases the phenylephrine-induced contraction mainly via an endothelium-dependent mechanism involving the inactivation of NO by ROS, which is produced by indigo carmine.
Indigo carmine (10-5 M) produced a significant leftward shift in the phenylephrine concentration-response curve and increased the phenylephrine-induced maximum contraction in the endothelium-intact aorta, whereas indigo carmine had no effect on the maximal contraction and produced a slightly leftward shift in the phenylephrine concentration-response curve in the endothelium-denuded aorta. Similar to a previous report [6], the magnitude of the indigo carmine-induced enhancement of the contraction evoked by phenylephrine was higher in the endothelium-intact aorta than in the endothelium-denuded aorta. This suggests that the indigo carmine-induced potentiation of the contraction evoked by phenylephrine was mainly endothelium-dependent. Endothelial NO, which is one of the most important endothelium-derived vasodilators, increases the level of cyclic guanosine monophosphate, leading to vasodilation [8]. Indigo carmine inhibits endothelium-dependent vasodilation by interfering with the acetylcholine-induced NO-mediated relaxation [6]. The superoxide radical is produced by the autoxidation of the leuco form of indigo carmine, which is produced by NADPH-cytochrome P450 reductase [7]. ROS, particularly superoxide anion (O2·-) and hydrogen peroxide (H2O2), are important signaling molecules in cardiovascular cells [11]. Enhanced production of O2·- increases NO inactivation [11], which leads to a decrease in endothelium-dependent NO-mediated relaxation. Endothelial NO was reported to inhibit the contractile agonist (e.g., phenylephrine and norepinephrine)-induced contraction of the rat aorta [12,13]. A low-molecular-weight superoxide anion scavenger tiron abolished the indigo carmine-induced potentiation of contraction evoked by phenylephrine. These results, along with previous reports [6,7,11-13], suggest that indigo carmine enhances the phenylephrine-induced contraction in the endothelium-intact rings via NO inactivation induced by indigo carmine-induced ROS production (Fig. 6). Reinforced with our results from isometric tension measurements in current in vitro study, indigo carmine increased the fluorescence of oxidized DCF, which is considered to be probe of ROS, such as H2O2 and O2·-, whereas a low-molecular-weight superoxide anion scavenger, tiron, abolished the indigo carmine-induced increase in the fluorescence of oxidized DCF [15,16]. In addition, indigo carmine was reported to induce the production of superoxide radical [7]. Considering a previous report [7], these results suggest that the decreased NO bioavailability due to the indigo carmine-induced production of superoxide radicals might be involved in the augmented response to phenylephrine in endothelium-intact rings (Fig. 6). As the typical cardiovascular response to the intravenous administration of indigo carmine appears to be the stimulation of the alpha-1 adrenoceptor [5], the indigo carmine-induced enhancement in the endothelium-denuded aorta and L-NAME-treated endothelium-intact aorta might be associated with the indigo carmine-induced potentiation of the phenylephrine-induced alpha-1 adrenoceptor-mediated contraction of the vascular smooth muscle.
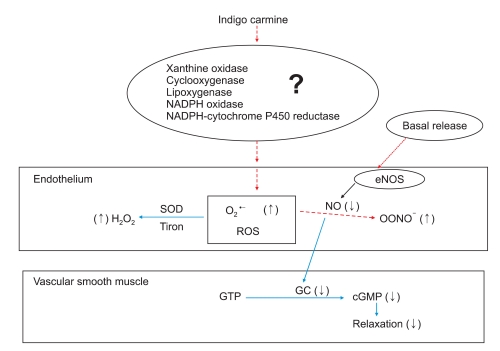 | Fig. 6Putative mechanisms responsible for the indigo carmine-induced inactivation of endothelial nitric oxide (NO), which may be involved in the indigo carmine-induced enhancement of the contraction evoked by phenylephrine in an isolated rat aorta [7,14,15]. The basal NO is produced by endothelial nitric oxide synthase (eNOS) and NO normally diffuses out of the endothelium to react with guanylyl cyclase (GC), which leads to the production of cyclic guanosine monophosphate (cGMP) and vasorelaxation. Superoxide anion (O2·-), which may be produced by enzyme systems (e.g., xanthine oxidase, cyclooxygenase, lipoxygenase, NADPH oxidase, NADPH-cytochrome P450 reductase) stimulated by indigo carmine, inactivates NO to form peroxynitrite (ONOO-), which leads to the decreased formation of cGMP and the inhibition of vasorelaxation. The decreased formation of cGMP may contribute to the enhanced response to phenylephrine. A low-molecular-weight superoxide anion scavenger, tiron, or superoxide dismutase (SOD) convert O2·- to a more stable reactive oxygen species (ROS), hydrogen peroxide (H2O2), leading to the inhibition of NO inactivation induced by O2·- and the restoration of production of cGMP, which attenuates the augmented response to phenylephrine. (↑): increase, (↓); decrease, GTP: guanosine triphosphate, NADPH: nicotinamide adenine dinucleotide phosphate.
|
Pretreatment with L-NAME completely abolished the indigo carmine-induced potentiation of the maximum contraction evoked by phenylephrine. Moreover, the L-NAME-induced contractile response was attenuated in the endothelium-intact rings that developed a stable phenylephrine-induced contraction in the rings pretreated with indigo carmine. As indigo carmine inhibits the endothelium-dependent relaxation induced by acetylcholine (receptor-mediated endothelium-dependent relaxant), calcium ionophore A23187 (nonreceptor-mediated endothelium-dependent relaxant) and endothelium-independent relaxation induced by sodium nitroprusside (endothelium-independent NO donor), the inhibitory effect of indigo carmine on the agonist-induced NO-mediated endothelium-dependent relaxation appears to be similar to the action mechanism of the nonspecific soluble guanylyl cyclase inhibitor, methylene blue [6,17,18]. These results in addition to those reported in previous studies [6,17,18], suggest that the indigo carmine-induced potentiation of the phenylephrine-induced contraction might be partly associated with an endothelium-dependent mechanism involving decreased NO synthesis. Therefore, more study on effect of indigo carmine on NO release and cyclic guanosine monophosphate formation induced by basal (unstimulated) state or contractile agonist is needed.
In the endothelium-intact rings pretreated with 10-5 M indomethacin, indigo carmine (10-5 M) increased the phenylephrine-induced maximum contraction, suggesting that the indigo carmine-induced potentiation of the phenylephrine concentration-response curve is not associated with a decrease in endothelial prostacyclin production.
Indigo carmine at 10-5 M (4.46 µg/ml), which is within the concentration encountered in a clinical setting [6], increased the phenylephrine-induced contraction. Any clinical implication of indigo carmine on the regional hemodynamics must be tempered by the fact that the aorta was used in this in vitro study, whereas the resistance vessels with the arterioles with a diameter of 100-300 µm control organ blood flow [19]. Even with this limitation, the indigo carmine-induced enhancement of the contraction evoked by phenylephrine might contribute to the transiently increased blood pressure observed in previous in vivo studies [2-5]. Intravenous indigo carmine increased the total peripheral vascular resistance, diastolic and systolic pressure and central venous pressure [4]. Generally, indigo carmine-induced hypertension is transient, not harmful and frequently undetected unless the patients are closely monitored for a brief period [4]. However, although the indigo carmine-induced hypertension is transient, it might be harmful to patients with an impaired left ventricular function. On the other hand, indigo carmine-induced hypertension might be less potent in patients with compromised endothelial integrity, such as hypertension and diabetes, than in patients with a normal endothelial integrity [20].
In conclusion, indigo carmine (10-5 M) at clinically relevant concentrations increases the phenylephrine-induced contractions mainly through an endothelium-dependent mechanism. The endothelium-dependent mechanism appears to be associated with a decrease in endothelial NO bioavailability by ROS, such as the superoxide radical, produced by indigo carmine.
Go to :
 XML Download
XML Download