

 PDF
PDF Citation
Citation Print
Print


Introduction
Ionizing radiation therapy is an effective approach to local tumor control. It plays an indispensable role in the treatment of non-small cell lung cancer (NSCLC) and other types of cancer. However, for metastatic lesions, radiotherapy is always limited to palliative symptoms and improvement of quality of life. Stereotactic ablative radiotherapy (SABR) optimizes local control by providing tumor-ablative doses with minimal damage to adjacent normal tissues and organs [1]. Accumulating evidence has shown that, SABR, delivered 1 to 3 fractions, can alter the inflammatory tumor microenvironment to trigger adaptive immune responses [2], which finally mediates tumor regression. One such observation is defined as “abscopal effect” and referred to nonirradiated tumor partial or complete eradication, suggesting an indirect systemic antitumor effect primed by local SABR [3]. This effect has been reported to date in NSCLC [4], melanoma [5], and other tumor species [6].
However, the incidence of abscopal effect is infrequent, and it has only been reported in a few cases. This may be because individual radiotherapy is generally unable to overcome the immunosuppressive factors of tumor-bearing host, including regulatory T cells (Tregs), myeloid-derived suppression cells (MDSC), and immuno-checkpoint (programmed death 1/programmed death-ligand 1 [PD-1/PD-L1] axis and cytotoxic T-lymphocyte antigen 4) [7]. Current strategies include radiotherapy combined with other regimens, such as Tregs depletion [8], cytokine addition [9], and in particular, combined with the PD-1/PD-L1 inhibitors. Given the recent success of cancer immunotherapy, SABR-induced abscopal effects may become widely seen [10,11].
Apatinib, a novel small molecule tyrosine kinase inhibitor (TKI), exerts its antitumor effects by specifically acting on vascular endothelial growth factor receptor 2 (VEGFR2), and it showed tolerance to a single-agent toxicity profile [12,13]. In 2014, apatinib was approved by the Chinese State Food and Drug Administration for the treatment of chemotherapy-refractory gastric cancer [14]. It also manifested satisfactory efficacy among patients with NSCLC [15]. Emerging data demonstrated that vascular endothelial growth factor (VEGF)/ VEGFR2, which is a pro-angiogenic factor, not only plays a critical role in promoting tumor angiogenesis but also participates in modulating tumor-induced immunosuppression (accumulation of Tregs, MDSC, and immature dendritic cells) that enables the tumor to evade host immunosurveillance [16]. Therefore, we here hypothesize that apatinib may have the ability to change the tumor tolerogenic microenvironment.
This study addressed the issue of whether SABR-elicited antitumor immunity could be significantly enhanced with a novel TKI-apatinib, then further leading to the suppression of both irradiated and nonirradiated (primary and secondary tumor, respectively) tumor growth.
Materials and Methods
1. Cell lines and animals
Mouse-derived Lewis lung carcinoma (LLC) cells and colon adenocarcinoma cell line (MC38) were purchased from American Type Culture Collection (ATCC) and cultured as described [8]. C57BL/6 mice (male, 4- to 5-week-old) were purchased from Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China).
2. Tumor challenge and treatment
C57BL/6 mice were injected subcutaneously with LLC cells or MC38 cells as indicated in each experiment. Tumor volumes were calculated as length×width2×0.5. Before irradiation, all mice were lightly anesthetized by intraperitoneal injection of propofol (150 mg/kg). Then, mice were positioned on a customized restraining device, and the whole body was protected by lead shielding except for the tumor area to be irradiated. Mice received a single dose of 15 Gy at a rate of 600 cGy/min using 12 MeV electrons with Siemens Oncor Impression (Siemens Medical Systems, Concord, CA). Apatinib was provided by Hengrui Pharmaceutical Group Co. Ltd. (Lianyungang, China) and administered by intragastric administration (i.g.) to mice once a day and for a total of 7 days.
3. Immunohistochemical staining
Immunohistochemistry was performed as previously described [17]. Sections were incubated with primary antibodies overnight at 4°C, including phospho-STAT3 (p-STAT3), CD4, CD8, p-VEGFR2, hypoxia-inducible factor-1α, and PD-L1 (all from Cell Signaling Technology, MA). Then, the sections were incubated with the secondary horseradish peroxidase conjugated antibody (Absin, Shanghai, China) for 30 minutes at room temperature. Targeted proteins were visualized with diami-nobenzidine (Zhongshan Golden Bridge, Beijing, China). Five random fields were selected for estimating the staining intensities by three independent observers individually.
4. Flow cytometric analysis
To obtain single-cell suspensions, the spleen was first scraped with scissors, and the homogenate was then passed through a 0.45 μm nylon mesh. Separated splenocytes were immuno-stained with anti-mouse CD3, CD4, CD8, Foxp3, and CD25 (all from BD Pharmingen, San Diego, CA) for 30 minutes at 4°C. Samples were collected on a FACSCalibur Flow Cytometer (BD Pharmingen), and data were analyzed with FlowJo software (BD Biosciences, San Diego, CA). An example of the gating strategy used for selection of CD4+ and CD8+ T cells and CD4+ CD25+ Foxp3+ Tregs cells is presented in S1 Fig.
5. Interferon-γ ELISPOT assay and enzyme-linked immunosorbent assay
Interfeon γ (IFN-γ)–producing lymphocytes were detected with ELISPOT according to the manufacturer’s instructions (Dakewe Biotech Co. Ltd., Shenzhen, China). Spleen single cells (2×105) from tumor-bearing mice were cultured with 2×103 LLC cells or MC38 cells in 96-well tissue culture plates precoated with an anti–IFN-γ antibody. Each positive spot represents an active lymphocyte. The supernatants were removed and stored at –80°C for further enzyme-linked immunosorbent assay against cytokines.
6. Statistical analysis
Data were analyzed with Prism 7.0 software (GraphPad Software, San Diego, CA). Data are represented as the mean±standard error of mean for all figures panels in which error bars are shown. Statistical analyses were carried out with Student's t test for two groups. For comparison among treatment groups, statistical analysis was performed using analysis of variance followed by Dunnett t test and log-rank test. The results were considered statistically significant at p < 0.05 and 0.01.
Results
1. Apatinib synergizes with SABR in the LLC mouse model
Previous research indicated that apatinib was able to downregulate the PD-L1 expression through the inactivation of STAT3 [18,19]. In this study, we found that the PD-L1 expression of LLC cell could be reduced by apatinib in a dose-dependent manner (S2A Fig.). As shown in S2B Fig., when VEGFR2 was activated by vascular endothelial growth factor (VEGF) as evidenced by increased p-VEGFR2 level, the expression of p-STAT3 and PD-L1 was elevated, and the total STAT3 expression was unchanged. Additionally, the elevation of PD-L1 was neutralized after deactivating STAT3 by S3I-201, an inhibitor of STAT3. These results suggested that apatinib could downregulate PD-L1 expression through VEGFR2/STAT3 pathway.
It has been proved that PD-1/PD-L1 axis restrains the effect of SABR [10,11,20], the function of apatinib on the SABR treatment was further examined in a LLC mouse model. LLC cells were injected into C57BL/6 mice at two separate sites as illustrated (Fig. 1A). For the single 15 Gy modality without apatinib, tumor growth delay was observed only at the irradiated lesion (primary tumor). When SABR and apatinib were combined, significant retardation of primary tumor growth could be observed. Especially, the growth of secondary tumor volume was different between SABR plus apatinib of 200 mg/kg group and apatinib 200 mg/kg group (p < 0.05) (Fig. 1B); however, all the secondary tumors in these two groups were not received radiotherapy, while the primary tumors in the combined group received radiotherapy, indicating that the irradiation of the primary tumor had an impact (abscopal effect) on the tumor shrink-age of the secondary tumor.
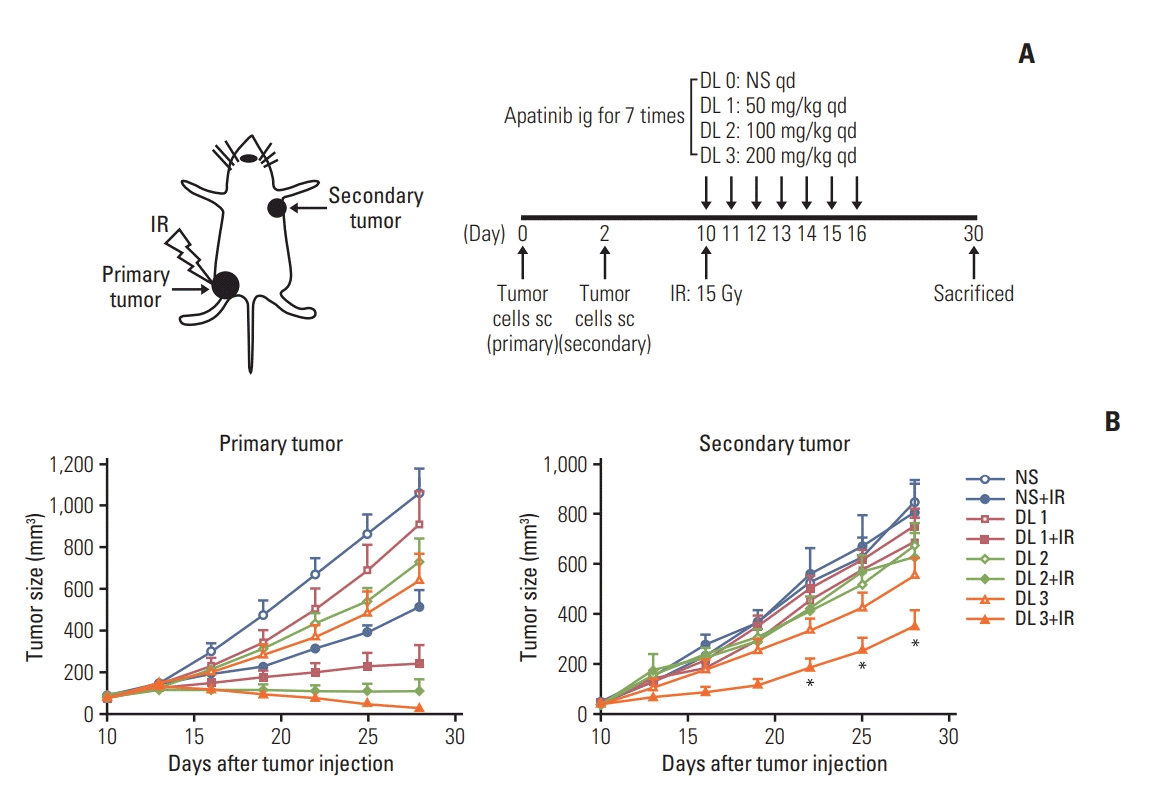
Fig. 1.
High dose of apatinib synergizes with stereotactic ablative radiotherapy in the Lewis lung carcinoma (LLC) mouse model. (A) C57BL/6 mice were injected subcutaneously with 5×105 LLC cells in the left hindlimb (primary tumor; irradiated) on day 0, and in the right flank (secondary tumor; nonirradiated) on day 2. On day 10, mice were administered intragastrically with varying doses of apatinib (NS; dose level [DL] of 50 mg/kg, 150 mg/kg, and 200 mg/kg) daily for 7 days. A single dose of 15 Gy radiotherapy to the primary tumor was administrated to half of the mice in each DL group on day 10. Tumor growth was evaluated every 3 days until day 30. (B) Tumor growth delay of primary tumors (left) and secondary tumors (right) in mice. Representative data are shown from two experiments conducted with five mice per group. *p < 0.05, **p < 0.01. IR, irradiation; NS, normal saline; qd, once daily; sc, subcutaneous administration.
![]()
In the toxicity analysis, we found that ionizing radiation combined with apatinib 200 mg/kg did not lead to significant weight loss or treatment-related death (Fig. 2A). Blood analysis was performed on day 30 when the mice were sacrificed. The results showed a trend toward mildly reduced frequency of white blood cells counts in mice treated with both single modality of apatinib (200 mg/kg) and combined strategy with SABR, without showing any other myelosuppressive toxicity (Fig. 2B).
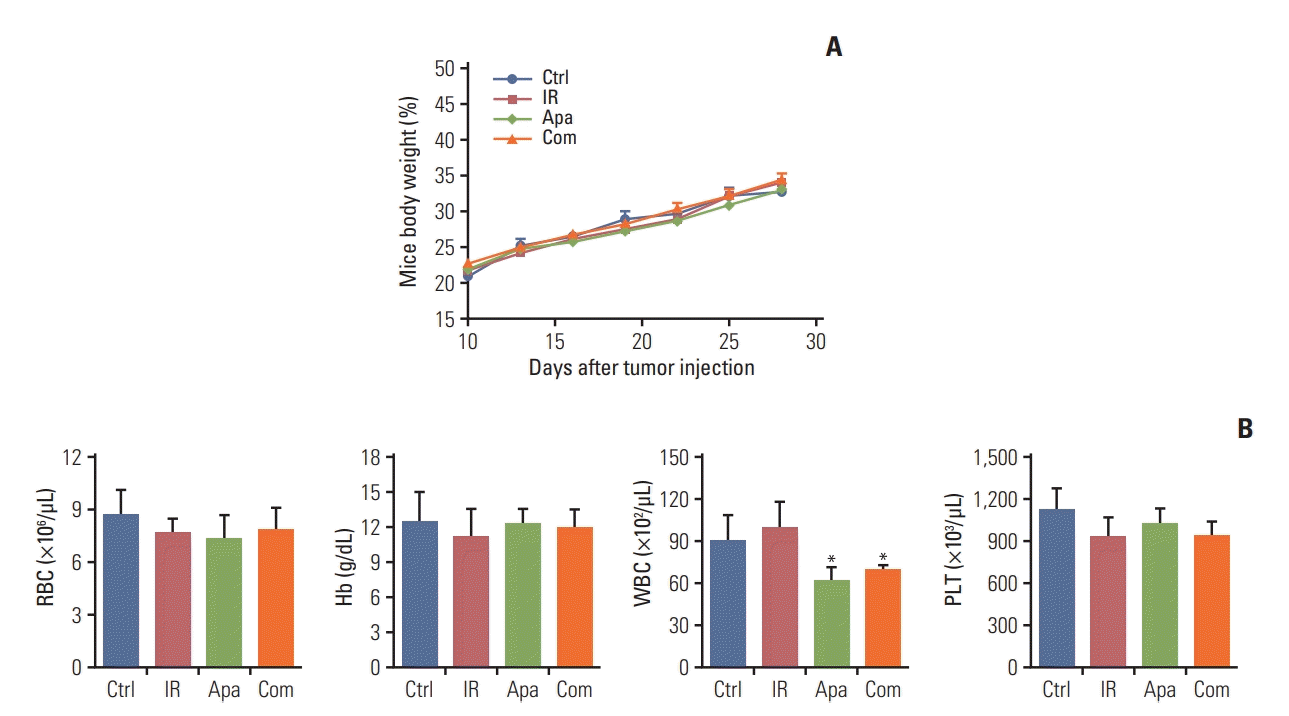
Fig. 2.
Toxicity analysis for treatments with stereotactic ablative radiotherapy (SABR) and apatinib in C57/BL6 mice bearing Lewis lung carcinoma. (A) Total body weight of high-dose level (200 mg/kg) apatinib-treated plus SABR mouse groups was monitored every 3 days. (B) Blood analysis of mice after various treatments. *p < 0.05. (A, B) Representative data are shown from two experiments conducted with five mice per group. Ctrl, control; IR, irradiation; Apa, apatinib; Com, combination treatment; RBC, red blood cells; Hb, hemoglobin; WBC, white blood cells; PLT, platelets.
![]()
2. Increased PD-L1 expression in tumor tissue following SABR could be overcome by concurrent apatinib therapy
The mechanism underlying the enhanced antitumor effects of SABR mediated by apatinib was further explored. We investigated whether PD-L1 expression could be down-regulated with apatinib treatment in vivo. PD-L1 expression of irradiated tumors was significantly greater after SABR (p < 0.01) (S3 Fig.). Interestingly, an increased expression of PD-L1 was also observed on nonirradiated secondary tumors after irradiation, compared with those of observed in control group (p < 0.05) (Fig. 3). However, both of which were reversed by additional apatinib therapy through reduced the phosphorylation levels of VEGFR2 and STAT3 (Fig. 3, S3 Fig.). Therefore, the result partially explains the synergistic mechanism of the combined radiotherapy and apatinib strategy.
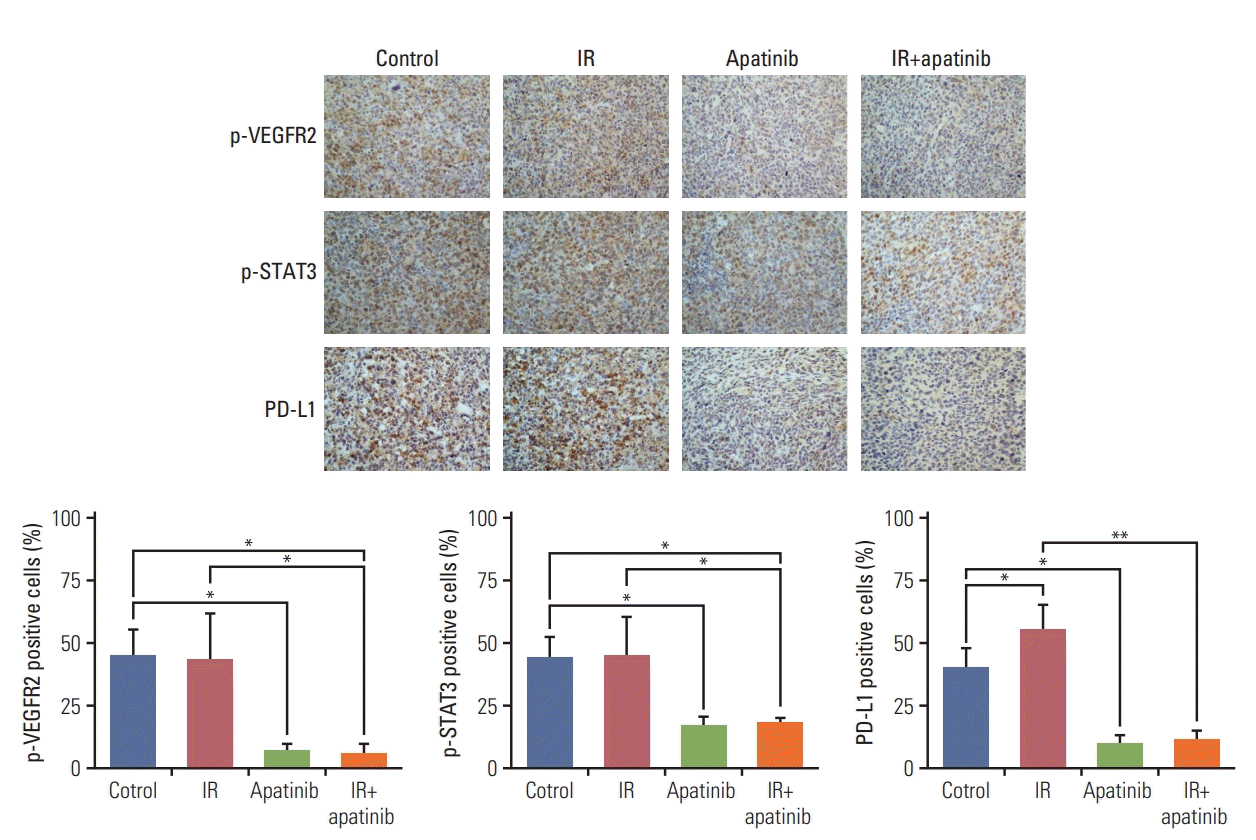
Fig. 3.
Increased programmed death-ligand 1 (PD-L1) expression in nonirradiated tumor tissue following stereotactic ablative radiotherapy could be overcome by concurrent apatinib therapy. The expression levels of PD-L1, phospho–vascular endothelial growth factor receptor 2 (VEGFR2), and phospho-STAT3 in nonirradiated tumor were detected by immunohistochemical (×40). Representative data are shown from two experiments conducted with five mice per group. *p < 0.05, **p < 0.01. IR, irradiation.
![]()
3. Apatinib combined with SABR could modulate the composition of immune cells and elicit activation of T lymphocyte
Subsequently, the lymphocyte population in the spleens of tumor-bearing mice was assessed. We found that the overall percentage of CD4+ and CD8+ T cells was significantly increased with the increase of apatinib dose. However, the percentage of CD4+ CD25+ Foxp3+ Tregs was significantly decreased, all in a dose-dependent fashion (Fig. 4A). Analysis of the presence of tumor-infiltrating lymphocytes (TILs) in secondary tumors indicated that, treatment with concurrent SABR and apatinib led to a significant increase in CD4+ and CD8+ TILs (p < 0.01, compared with all other groups) (Fig. 4B). ELISPOT assay showed that the number of activated IFN-γ–secreted lymphocyte was significantly greater only in mice receiving combined treatment (p < 0.05, compared with all other single groups) (Fig. 4C). The overall level of IFN-γ secreted from splenic lymphocytes was significantly greater in combination therapy, compared with that of single apatinib or SABR treatment (Fig. 4D, left). On the contrary, interleukin 10, which was involved in tumor-mediated immune suppression, was significantly decreased in the combined group (Fig. 4D, right).
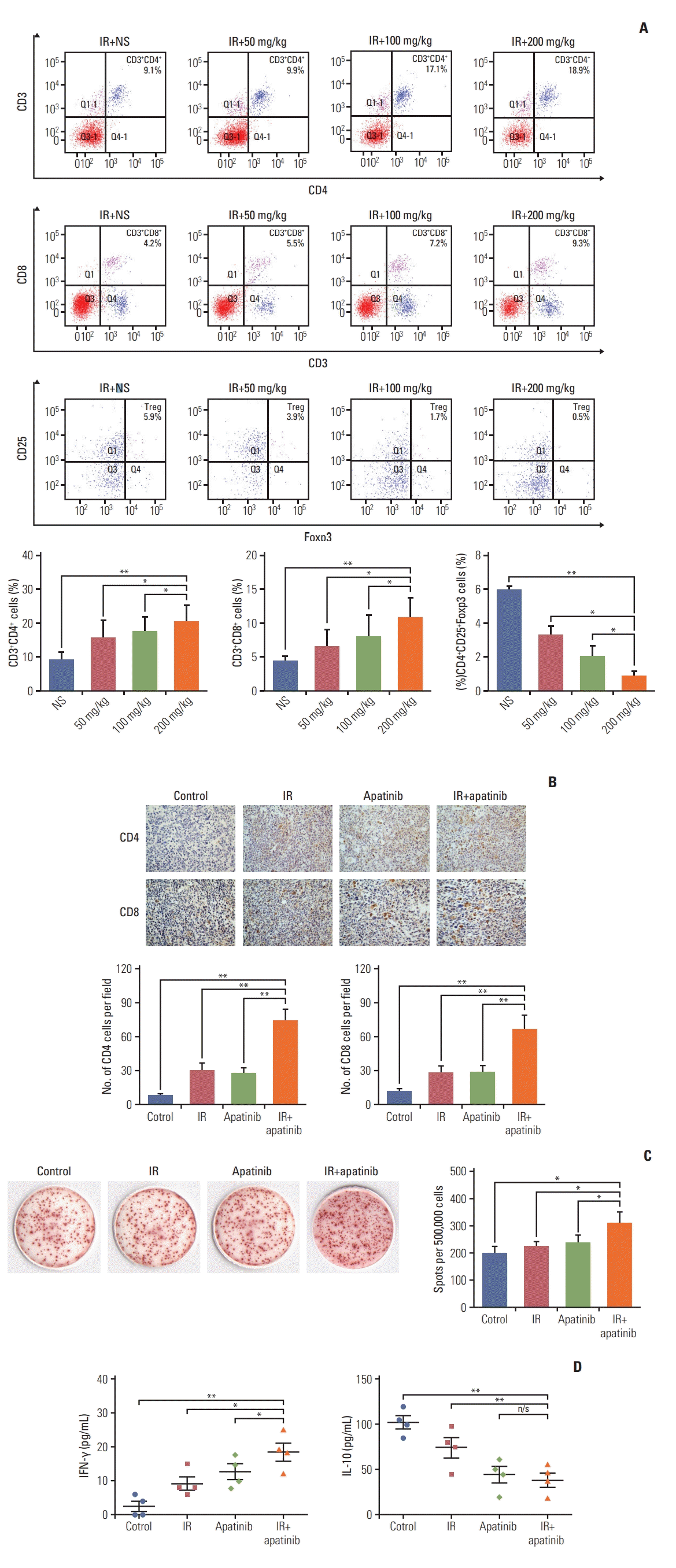
Fig. 4.
The combination of apatinib and stereotactic ablative radiotherapy could modulate the composition of immune cells, elicit tumor-specific T-cell activation and result in varied cytokine expression profiles. (A) The lymphocyte population in the spleen of tumor-bearing mice were assessed by flow cytometry. NS, normal serum. (B) The CD4+ and CD8+ tumor-infiltrating lymphocytes in the secondary tumors were stained by immunohistochemical (×40). (C, D) Interferon γ (IFN-γ)–producing lymphocytes from spleen single cells were detected with ELISPOT assay; the supernatants were removed and stored at –80°C for further cytokine detection with enzyme-linked immunosorbent assay. IR, irradiation. *p < 0.05, **p < 0.01, n/s, no significant. (A-D) Representative data are shown from three experiments conducted with five mice per group.
![]()
Collectively, these results suggest that apatinib with combined SABR can modify the tumor microenvironment that may be associated with tumors outside the radiation field rejection.
4. Combined radiotherapy and apatinib therapy evoked tumor antigen–specific immune responses
To demonstrate that the applicability of combined SABR and apatinib therapy is tumor antigen–specific, LLC cells were injected into C57BL/6 mice as primary and secondary tumors and MC38 were injected as the third experimental tumor. Primary LLC were given a single dose of 15 Gy (Fig. 5A). Consistent with previous results, in both the primary irradiated and secondary nonirradiated LLC tumor sites, slowed tumor progression could be significantly induced with SABR in combination with apatinib (Fig. 5B). However, there was no difference in efficacy of MC38 tumor growth in the combined therapy group over apatinib single-agent therapy (p > 0.05) (Fig. 5B). For the response to LLC, the number of IFN-γ–producing lymphocytes in the combination treatment group was significantly higher than in the other three groups. However, for MC38, the positive spots counts were not affected by SABR, apatinib, or the combination therapy (Fig. 5C). From these observations, we conclude that the antitumor abscopal effect induced by combination radiotherapy and apatinib was tumor antigen–specific.
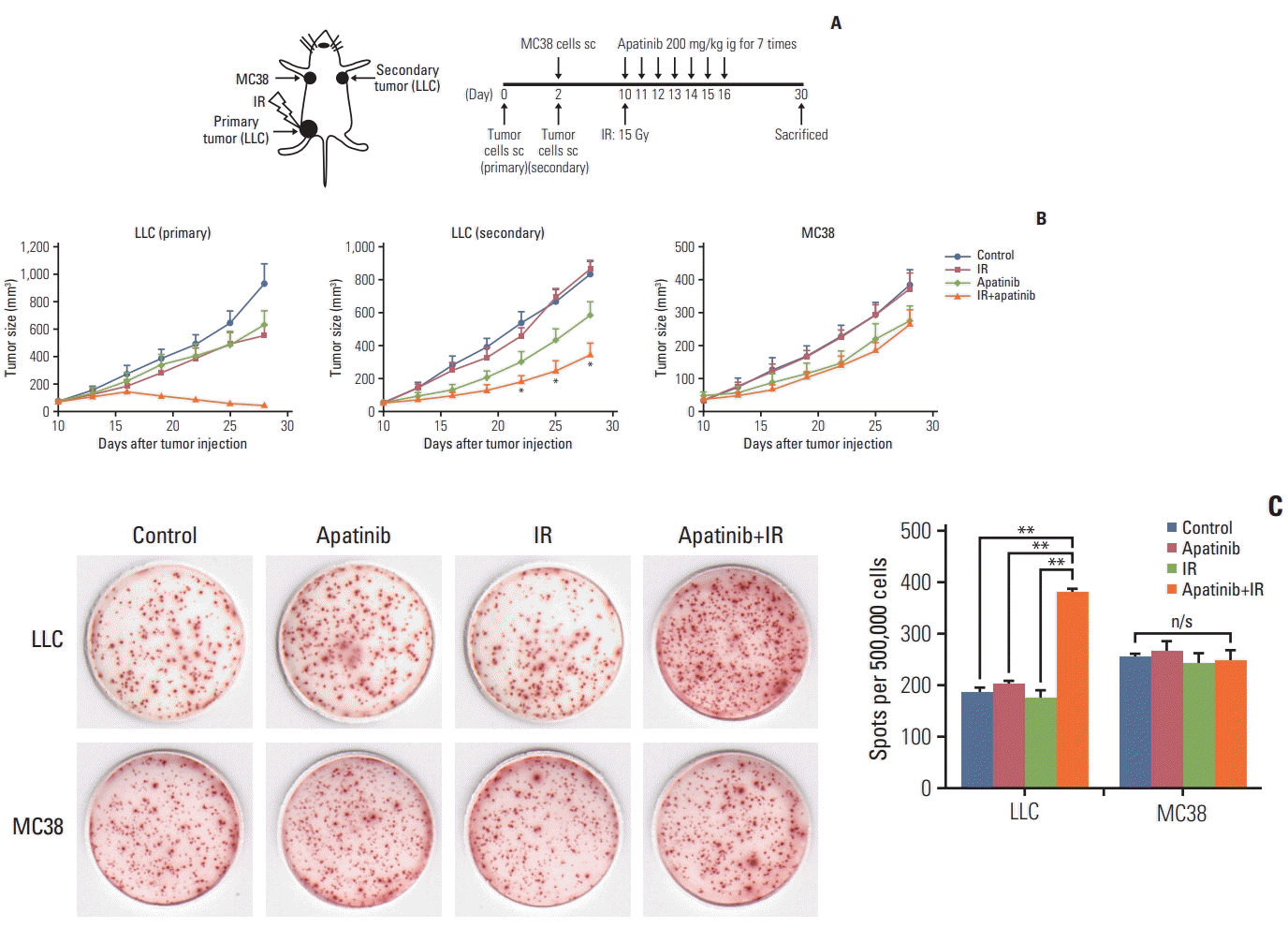
Fig. 5.
Combination radiotherapy and apatinib therapy evoked tumor antigen–specific immune responses. (A) Mice were injected subcutaneously with 5×105 Lewis lung carcinoma (LLC) cells in the left hindlimb (primary tumor; irradiated) on day 0, and in the right flank (secondary tumor; nonirradiated) on day 2. The third-party tumor cells (MC38) were inoculated subcutaneously with 5×105 cells in the left flank (nonirradiated) on day 2. A patinib 200 mg/kg was administered intragastrically to mice from day 10 to 16 for 7 times an d primary LLC was administered as a single 15-Gy dose. (B) Tumor growth delay of primary LLC (left), secondary LLC (middle) and secondary MC38 tumor (right). IR, irradiation; ig, intragastric administration; sc, subcutaneous administration. (C) The number of interferon-γ–producing lymphocytes response to LLC or MC38 cells was detected with ELISPOT assay. *p < 0.05, **p < 0.01, n/s, no significant. (B, C) Representative data are shown from two experiments conducted with five mice per group
![]()
5. Apatinib and radiotherapy treated tumor-bearing mice were resistant to tumor rechallenge and presented improved long-term survival
To facilitate a broader application of this novel simultaneous strategy, we next investigated whether this combination therapy could prevent metastasis and further increase survival. LLC cells were injected in the left hindlimb as primary tumor. After all the treatments were complete, mice were rechallenged with LLC cells on the opposite flank. Both tumor growth and survival time were monitored (Fig. 6A). As shown in Fig. 5B, for rechallenged tumors, SABR or apatinib therapy alone showed negligible inhibition of tumor growth compared with that of control group. However, when SABR was combined with apatinib, tumor growth was much slower than that of in other groups (Fig. 6B). As for survival analysis, survival could be more effectively improved with the combined treatment of radiation therapy and apatinib, and 40% of mice were still alive until the end of the experiment (Fig. 6C). Together, these results suggest that the two-pronged regimen could not only prevent tumor metastasis but also prolong the survival time in tumor-bearing mice.
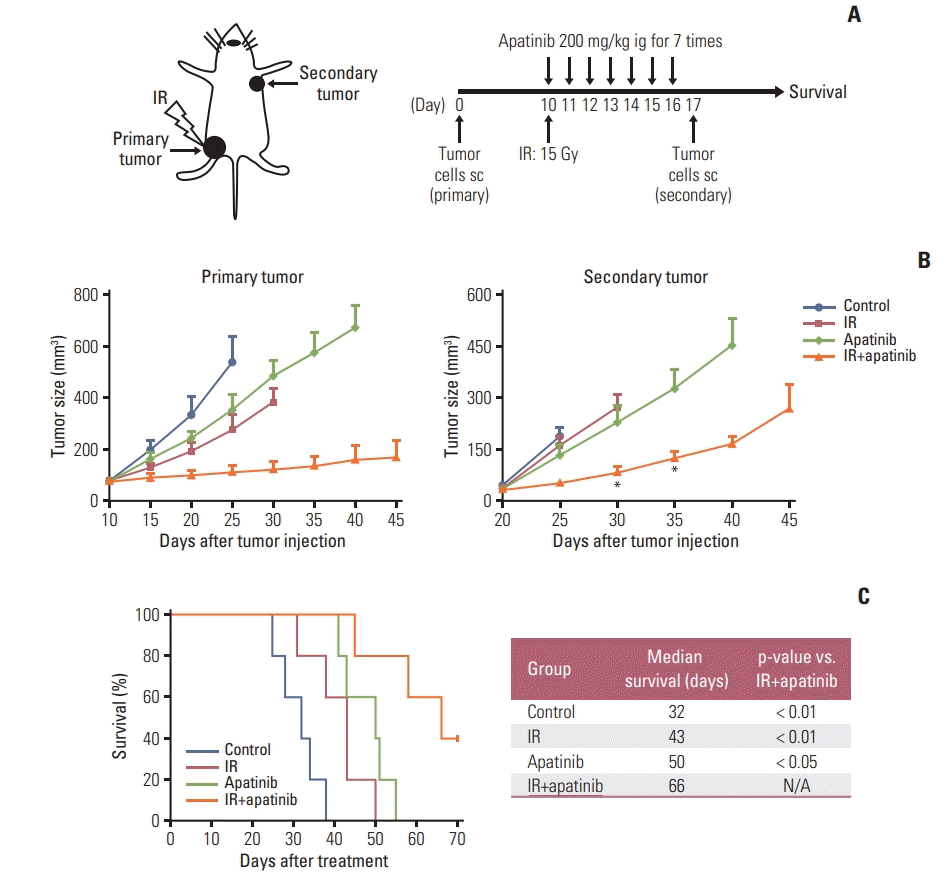
Fig. 6.
Tumor-bearing mice with combined stereotactic ablative radiotherapy and apatinib treatment were not only resistant to the tumor rechallenge but also improving long-term survival. (A) Lewis lung carcinoma (LLC) cells (5×105) were injected in the left hindlimb as primary tumor, after all the treatments finished, mice were rechallenged with LLC cells (5×105) on the opposite flank. (B) Tumor growth delay of primary tumor (left) and secondary rechallenge tumor (right). *p < 0.05, **p < 0.01. (C) Kaplan-Meier survival curves and life table for LLC bearing mice. (B, C) Representative data are shown from two experiments conducted with five mice per group. IR, irradiation; ig, intragastric administration; sc, subcutaneous administration; N/A, not available.
![]()
Discussion
Our proof-of-principle study suggested that, apatinib, a novel VEGFR2 TKI, may modulate the tumor microenvironment, not only by reversing the PD-L1 expression increased by SABR but also by changing other constituents. Especially at a high dose, apatinib worked in a synergistic manner with SABR, and induced an abscopal effect. In addition, we demonstrated that SABR-induced antitumor immunity was tumor antigen–specific, and the combined SABR and apatinib therapy could resistant to tumor rechallenge, thereby extending survival time.
Many studies have shown that VEGFA/VEGFR2 is not only a pivotal growth factor that promotes angiogenesis but also directly supports cancer cell growth through an autocrine- positive feedback loop [21]. Apatinib (YN968D1) is a novel and highly selective competitor for the ATP binding site of VEGFR2 tyrosine kinase, and it blocks downstream signaling transduction [12]. Thus, apart from the anti-angiogenetic effect, apatinib may directly inhibit tumor cells or affect other cells expressing VEGFR as a cytotoxic agent [12,22].
PD-L1 is expressed on many types of cells, including T cells, B cells, monocytes, and tumor cells, which suppresses T-cell function and results in immune escape by binding to PD-1 [23]. STAT3, a point of convergence for numerous tyrosine kinases (including VEGFR, platelet-derived growth factor receptor, epidermal growth factor receptor, and Src) in diverse cancer types including NSCLC [22,24]. STAT3 could bind to the PD-L1 promoter to regulate its expression transcriptionally [25]. It has been previously demonstrated that there was a significant correlation between PD-L1 and VEGFR2 expression in tumor cells and PD-L1 expression could be inhibited with apatinib by targeting STAT3 [18,19]. These results were consistent with our findings (Fig. 2, S2 and S3 Figs.).
Recent studies have reported that VEGFR2 is also selectively expressed on Tregs and can directly induce proliferation [26]. Drugs with anti-angiogenic properties can also counteract the proliferation of tumor-induced immunosuppressive cells. Sunitinib, another TKI-targeted VEGFR family, has been found to be able to decrease the number of Tregs in a dose-dependent fashion in colon-tumor-bearing mice [27]. It demonstrated that VEGF could directly suppress T-cell proliferation and activation via binding to VEGFR2 [28]. In this study, we found that the overall percentage of CD4+ and CD8+ T cells in the spleen could be significantly increased with apatinib, and the infiltration of T cells into the tumor could also be improved (Fig. 4A and B). This better infiltration could also be associated with the capacity of anti-angiogenic molecules to normalize tumor vasculature and preferentially promote infiltration of immune effector cells into the tumor parenchyma [29].
Increased evidence and our study indicated that SABR-induced a local inflammatory response that could enhance the infiltration of T lymphocyte and simultaneously induce PD-L1 expression in the tumor microenvironment (Fig. 3), which markedly weakens SABR-induced antitumor immunity. Based on the comprehensive immuno-modulatory capacity, anti-angiogenic molecules may be sufficient to enhance the systemic antitumor immunity mediated by SABR, especially at the sites remote from irradiation. This is the first report on apatinib—a novel anti-angiogenic drug—combined with SABR that elicited a curative abscopal effect. In addition, we also found that the antitumor abscopal effect induced by our dual therapy was tumor antigen–specific (Fig. 5C). This observation is consistent with the hypothesis that radiotherapy has the potential to trigger immunogenic tumor-cell death and induce danger signals that effectively exposes tumor-specific antigen, contributing to the phenomenon referred to as “in situ” vaccination [2]. Moreover, our study suggested herein that this novel dual therapy could protect tumor-bearing mice from tumor rechallenge. In this way, survival time could be prolonged (Fig. 6).
The current radiotherapy dose and best fractionation have been empirically validated. It was suggested that optimal antitumor immunity could be induced with 8 Gy×3 fractionated doses instead of single-dose irradiation in the preclinical model [30]. However, this is a sub-curative dose, we chose a single 15 Gy dose in our experiment and combined it with 200 mg/kg apatinib therapy, without increasing additional toxicity (Fig. 2). Park et al. [11] previously applied 15 Gy×1 irradiation regimen in animal models, without showing apparent toxicity and the effects on immune activation were superior to a 3 Gy×5 fractionated strategy.
In summary, our results suggested that apatinib could be synergistically applied with SABR to enhance the systemic antitumor efficacy through modifying tumor microenvironment. Moreover, our findings could broaden the scope of current strategy to reverse the immunosuppressive tumor environment and provide insights into rational design of clinical trials.
 XML Download
XML Download