

 PDF
PDF Citation
Citation Print
Print


Introduction
Acute promyelocytic leukemia (APL), which is known as acute myeloid leukemia-3, AML-3, or M3 according to the French-American-British (FAB) classification, is characterized by a predominance of malignant promyelocytes that carry a reciprocal translocation between the long arms of chromosomes 15 and 17: t(15;17)(q22;q11.2-q12) that results in the formation of a hybrid gene, PML–retinoic acid receptor α (RARα). The fusion protein PML-RARα has been shown to recruit methyltransferases on the CpG islands of the promoter region of the retinoic acid receptor RARα, resulting in the hypermethylation mediated silencing of RARα in APL cells [1]. In addition to the hypermethylation mediated silencing of the RARα gene, many tumor suppressor genes have been extensively reported in APL, particularly the p15 gene. p15Ink4b (p15) is a tumor suppressor gene located at 9p21, which encodes for p15 cyclin-dependent kinase inhibitor. The 9p21 chromosomal locus, which is referred to as the INK4/ARF locus, has been tightly linked to the formation of many types of tumors [2]. p15 belongs to the INK4 kinase family of cyclin-dependent kinase inhibitors (which consists of p15, p16, p19, and p21). These inhibitors negatively regulate the cell cycle through competitive inhibition of the cyclin-dependent kinases 4 and 6 involved in retinoblastoma (Rb)-dependent cell cycle regulation. p15Ink4b expression has been shown to increase specifically during myeloid differentiation in vivo in both human bone marrow and peripheral blood cells [3]. The role of p15Ink4b during myeloid differentiation was further supported by in vitro studies of the M1 leukemia cell line. When terminal differentiation is induced in these cells, upregulation of p15Ink4b expression is accompanied by inhibition of CDK4 kinase activity and a decrease in levels of phosphorylated Rb [4]. Furthermore, overexpression of p15Ink4b in M1 cells causes cell cycle arrest in the G1 phase, providing additional evidence that the protein is involved in maturation and cell cycle inhibition of late stage progenitors [5]. This function is further supported by studies in human CD34+ hematopoietic progenitor cells. When expression of p15Ink4b is triggered, higher levels of the protein are associated with transcriptional up regulation of genes known to induce myeloid differentiation. Studies conducted by Teofili et al. [6] suggested that p15 plays an important role in regulation of the proliferative activity of promyelocytes. Unlike other tumor suppressor genes, the p15 gene is preferentially hypermethylated at a 5’-CpG island, which has been shown to be associated with loss of transcription of this gene in leukemia cells [7,8].
In the present study, we attempted to evaluate the promoter hypermethylation of the p15 gene in newly diagnosed and relapsed APL patients treated with conventional chemotherapy protocols International Consortium on Acute Promyelocytic Leukemia-2006 (ICAPL-2006) and Arsenic Tri-Oxide (ATO) protocols. Moreover, mRNA expression of the p15 gene was analyzed using semi-quantitative reverse transcriptase polymerase chain reaction (RT-PCR) and correlated with p15 gene methylation and various clinico-pathological parameters.
Go to : 
Materials and Methods
A total of 37 APL cases were referred to the Department of Immunology and Molecular Medicine, Sher-I-Kashmir Institute of Medical Sciences (SKIMS), Srinagar (Jammu and Kashmir, India) from July 2013 to November 2015. We analyzed the course of all newly diagnosed and relapsed patients with APL treated consecutively on protocols (ICAPL-2006 and ATO) in the Department of Clinical Hematology and Medical Oncology at SKIMS. Patients were included in the study after approval from the ‘Institute Ethics Committee’ (IEC) of SKIMS and subjected to prospective evaluation of their response to conventional chemotherapy (ICAPL-2006 and ATO). All patients received baseline bone marrow analysis and were categorized against the morphologic criteria for the diagnosis of APL (APL AML-M3 or M3-variant) according to the FAB classification system [9]. About 5-6 mL of peripheral blood were collected from 37 newly diagnosed and relapsed patients into EDTA vials for methylation-specific polymerase chain reaction (MS-PCR) and RT-PCR after informed consent. The diagnosis was confirmed at our department by the presence of t(15:17) on peripheral blood cytogenetic studies, as well as detection of PML-RARα translocation by RT-PCR and quantitative PCR (q-PCR) Subsequently, blood samples were collected from healthy control subjects who did not harbor any hematological malignancies. The median age of the patients was 31 years (range, 6 to 92 years).
1. DNA extraction
Blood samples from each patient were diluted with equal volumes of RPMI-1640 cell culture media and subjected to density gradient centrifugation using Ficol Histopaque as density gradient material. Isolated white blood cells were equally distributed into two tubes for DNA and RNA extraction.
High molecular weight DNA was extracted from Ficol Histopaque isolated cells from patient samples and healthy control samples using the phenol-chloroform extraction protocol. The quality of extracted DNA was checked on 1% agarose gel and the concentration of the DNA obtained was measured in a spectrophotometer at 260 nm.
2. DNA methylation assay
MS-PCR was carried out according to the method described by Chim et al. [10]. Extracted DNA was subjected to bisulphite conversion using an EZ DNA Methylation kit from Zymo Research (Irvine, CA). MS-PCR analysis of P15 gene was carried out for its promoter using primer sets F: 5’-GCGTTCGTATTTTGCGGTT-3’ and R: 5’-CGTACAATAACCGAACGACCGA-3’ for methylated allele and F: 5’-TGTGATGTGTTTGTATTTTGTGGTT-3’ and R: 5’-CCATACAATAACCAAACAACCAA-3’ for unmethylated allele. Approximately 2.0 μL of the bisulphite converted DNA (normal and leukemic) from each sample was used for MS-PCR. Universal Methylated Human DNA (Zymo Research) was used as a positive control for methylated alleles, while DNA from normal healthy subjects was used as a control for unmethylated alleles. Water was used as a negative PCR control in both reactions. The following thermal conditions were for used for MS-PCR: 95°C for 5 minutes, followed by 35 cycles of 95°C for 40 seconds, 56°C for 35 seconds and 72°C for 45 seconds, and then final extension for 7 minutes at 72°C. Primer sequences of p15 promoter for the unmethylated reaction amplified a 154 bp product, while the methylated reaction amplified a 148 bp product (Fig. 1). PCR products were analyzed by 2% agarose gel electrophoresis as described above.
 | Fig. 1.Representative pic of methylation-specific polymerase chain reaction showing different patterns of p15 gene methylation in APL patients. Lane 1, molecular weight markers (MW, 50 bp); lanes 2 and 3, unmethylated p15 gene (U, 154 bp); lanes 4 and 5, partial methylated p15 showing both 154 and 148 bp bands (U/M); lanes 6 and 7, methylated p15 gene (M, 148 bp); lane 8, positive control for methylated DNA (PC, 148 bp); lane 9, negative control without template DNA (NC).
|
3. RNA extraction
Total RNA from Ficol Histopaque separated white blood cells from patients and healthy control subjects was extracted using Trizol (Amresco, LLC, Solon, OH). The extracted RNA was checked for purity and integrity by diethyl pyrocarbonate–treated gel electrophoresis. The concentration of RNA was measured spectrophotometrically and the ratio of the optical density at 260 and 280 nm was checked. A ratio of 1.80 to 2.00 was accepted as good quality RNA.
4. cDNA Synthesis
A Maxima cDNA synthesis kit (Thermo Scientific, Waltham, MA) containing enzyme mix (M-MuLV RT enzyme and Ribolock RNAse inhibitor), 5× reaction mix (reaction buffer, dNTPs, oligo(dt)18, random hexamer primers, and nuclease-free water) was used to perform the reverse transcriptase reaction.
Two micrograms of total cellular RNA were reverse transcribed to cDNA by incubation for 10 minutes at 25°C, 15 minutes at 50°C, and 5 minutes at 85°C in a total volume of 20 μL that contained 4 μL of 5× reaction mix, 2 μL of maxima enzyme mix and 12 μL of nuclease free water.
5. RT-PCR analysis
Multiplex PCR was carried out in a 25 μL reaction mixture using reverse and forward primers sets for p15 and β-actin. The primers used for p15 were F: 5’-TGGGGG CGGCAGCGATGAG-3’ and R: 5-́AGGTGGGTGGGGGTGGGAAAT-3’, while for β-actin a primer set of F: 5’-TGACGGGGTCACCCACACTGT-3’ and R: 5’-CTAGAAGCATTTGCGGTGGAC-3’ was used. Briefly, 2 μL of cDNA was amplified in a 25 μL reaction containing (Taq polymerase, Taq buffer, MgCl2, and dNTPs). cDNA synthesized from the RNA of healthy control subjects were used as a positive control, water was used as a negative control and β-actin served as an internal control. Reaction times consisted of an initial denaturation of 94°C for 5 minutes, annealing at 54°C for 30 seconds, and elongation at 72°C for 7 minutes, followed by 35 additional cycles (30 seconds at 94°C, 30 seconds at 54°C, and 30 seconds at 72°C). Next, 6-8 μL of the PCR product was size-fractionated by electrophoresis in 2% agarose gel stained with ethidium bromide and visualized under a UV trans-illuminator (Flourchem, HD2-Cell Biosciences, Santa Clara, CA) at 365 nm. Primer sequences of p15 mRNA amplified a 450 bp product; whereas a product of 680 bp was amplified with the primer set for β-actin mRNA.
Go to :
Results
The present study included a total of 37 APL patients comprising 22 males (59.5%) and 15 females (40.5%) ranging in age from 6 years to 91 years. Overall, 25 patients (67.6%) were < 30 years of age and 12 (32.4%) were ≥ 30 years of age (Table 1). Based on risk stratification, patients were classified as low risk, high risk or intermediate risk according to the total leukocyte count and platelet counts at presentation. Nine of the 37 patients (24.3%) presented with leukocytosis (> 1×1010/L), whereas the remaining 28 patients (75.7%) had leucopenia (≤ 1×1010/L). High platelet counts (≥ 4×1010/L) were detected in seven patients (19.0%), while 30 patients (81.0%) presented with low platelet counts (< 4×1010/L). Bone marrow examination revealed six patients (16.2%) with less than 60% promyelocytes and 30 (83.8%) with a promyelocyte count greater than 60% (Table 1).
Table 1.
Clinicopathological parameters of 37 APL patients at diagnosis according to the p15 methylation state
![]()
Among 22 male patients, nine (41.0%) were found to have hypermethylation and 13 (59.1%) were unmethylated, while among 15 female patients, seven (46.7%) had p15 gene methylation and eight (53.3%) were unmethylated patients. Moreover, p15 methylation status differed significantly between males and females (p=0.021).
Overall, 16 patients were found to have methylated p15 genes in which seven patients were methylated partially and nine had complete methylation based on the presence of either two or a single band, respectively, upon MS-PCR analysis. The remaining 21 patients were observed to be negative for p15 gene methylation, as depicted by the absence of a methylated band upon MS-PCR analysis (Fig. 1). Moreover, the p15 gene methylation status in APL patients was found to be significantly associated (p=0.031). The discrimination of unmethylated and methylated DNA was explored by MS-PCR in healthy control subjects in which amplification of unmethylated DNA was only seen as depicted by the presence of a specific DNA band with unmethylated primers.
Interestingly, all nine of the 37 patients (24.3%) who presented with leukocytosis at their baseline (> 1×1010/L) had complete p15 gene methylation, while among the remaining 28 patients with leucopenia (≤ 1×1010/L), seven (18.9%) had partial methylation of the p15 gene at baseline. This difference between groups was significantly associated (p < 0.05). The other clinicopathological parameters and their correlation with p15 gene methylation status are given in Table 1.
This observation was further supported by the semi-quantitative expression of the p15 gene observed upon RT-PCR analysis of samples from the same series of 37 APL patients. We found that no p15 gene mRNA was expressed in nine patients who presented with leukocytosis coupled with complete p15 methylation (M group). In comparison, p15 mRNA was expressed in all patients in whom p15 alleles were either partially methylated (U/M group) or were unmethylated (U group). Similarly healthy control subjects showed a consistent pattern of unmethylation in the p15 gene and their mRNA was found to be intact Table 2. Thus, p15 gene silencing was found exclusively in patients with high leukocyte counts in addition to complete methylation of both p15 alleles (p=0.031).
All 37 patients were treated with all-trans retinoic acid in addition to conventional chemotherapy (ICAPL-2006). During the course of the study, eight patients expired. Among these, six patients (four from group M and two from group U) died during the induction phase of treatment, while two patients (from M group) relapsed and expired during the maintenance stage of chemotherapy.
Overall, 31 patients were followed during the course of our study. All patients achieved bone marrow remission by the end of the consolidation phase of chemotherapy as confirmed by their negative q-PCR and RT-PCR status for PML-RARα. However, during the maintenance stage of chemotherapy, six patients (four from M group and one each from the U/M and U groups) relapsed. High leukocyte count was observed in all six relapsed cases who were subsequently subjected to treatment with arsenic trioxide. Upon follow-up evaluation of the p15 methylation status and p15 mRNA expression during maintenance, we found that all six relapsed patients had complete p15 methylation, while two patients with complete methylation status belonged to the U and U/M groups at baseline. These findings indicate the poor prognostic implication of acquired complete methylation. The loss of p15 mRNA expression was subsequently seen in all six of the relapsed cases. The only remaining patient from M group and patients from U and U/M groups who achieved remission had no p15 methylation, and p15 mRNA was found intact in these patients. Of the six relapse patients, two complete methylation expired because of cerebral hemorrhage.
Multivariate analysis was performed to determine if complete p15 methylation is an independent poor prognosis factor in APL patients. We observed that the group M patients had an inferior survival (hazard ratio, 8.60; 95% confidence interval [CI], 1.7 to 43.0; p=0.009) compounded with an increased risk of relapse (hazard ratio, 11.5; 95% CI, 1.9 to 135.4; p=0.050) relative to group U. In contrast, comparison of group M to group U/M revealed no significant difference in survival or risk of relapse. Therefore, the detection of complete p15 methylation status was found to be an independent poor prognosis factor for group M relative to group U that was not influenced by any other tested variables (e.g., age, sex, WBC count, platelet counts, treatment, and PML/RARα breakpoint type).
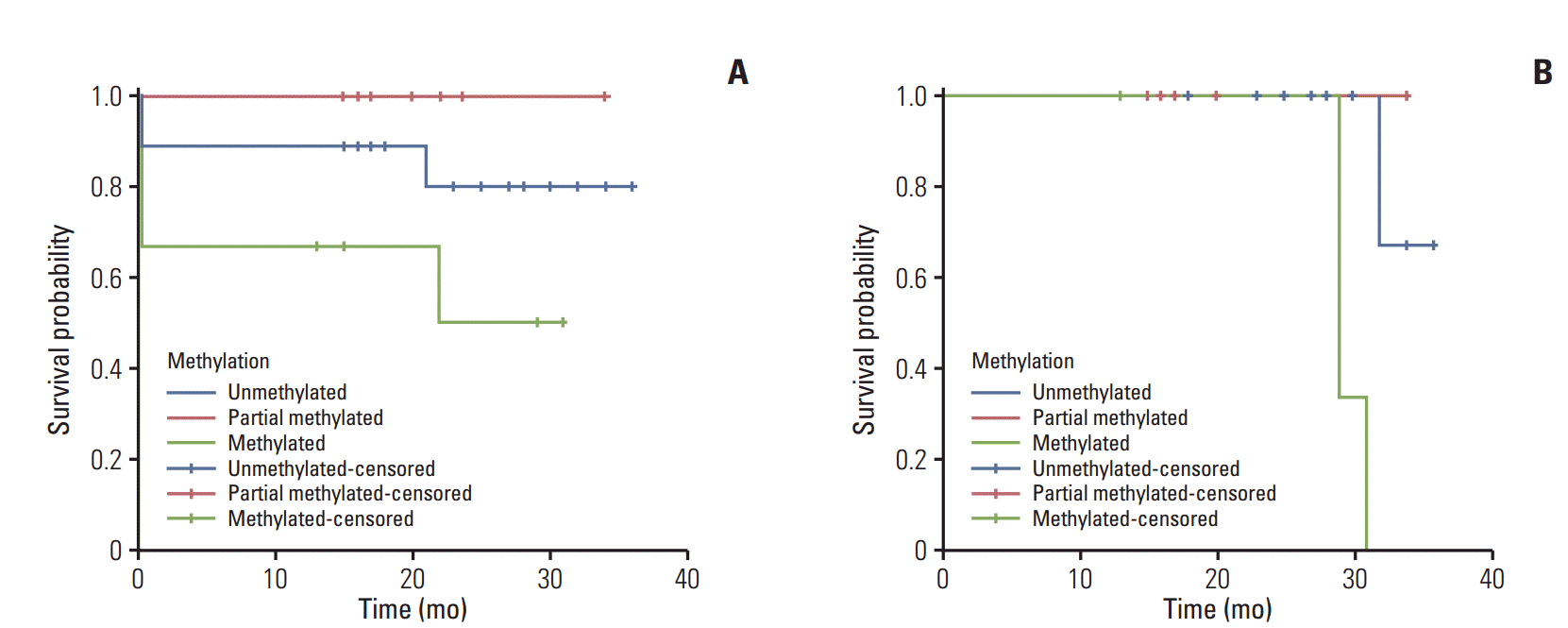
Clinical outcome according to the pattern of p15 methylation and its mRNA expression was analyzed in APL patients in terms of their overall survival (OS) and disease-free survival (DFS). Patients were followed for a mean duration of 19.1 months ranging from 0 to 34 months and Kaplan-Meier survival analysis was performed to assess the OS of all 37 patients and the DFS of 31 patients who achieved clinical remission. Comparison of survival between patient groups was based on the log-rank test.
The OS for groups M, U/M, and U patients was 38%, 100%, and 90%, respectively. Patients from group M showed the lowest OS, while those in group U/M showed a higher OS, but this difference was not significant (p=0.295) (Fig. 2A). We observed a significant difference in DFS (log-rank test, p=0.022), which was estimated to be 22%, 83%, and 93% in patients from groups M, M/U, and U, respectively (Fig. 2B). Moreover, patients from group M showed the lowest DFS (16.5 months).
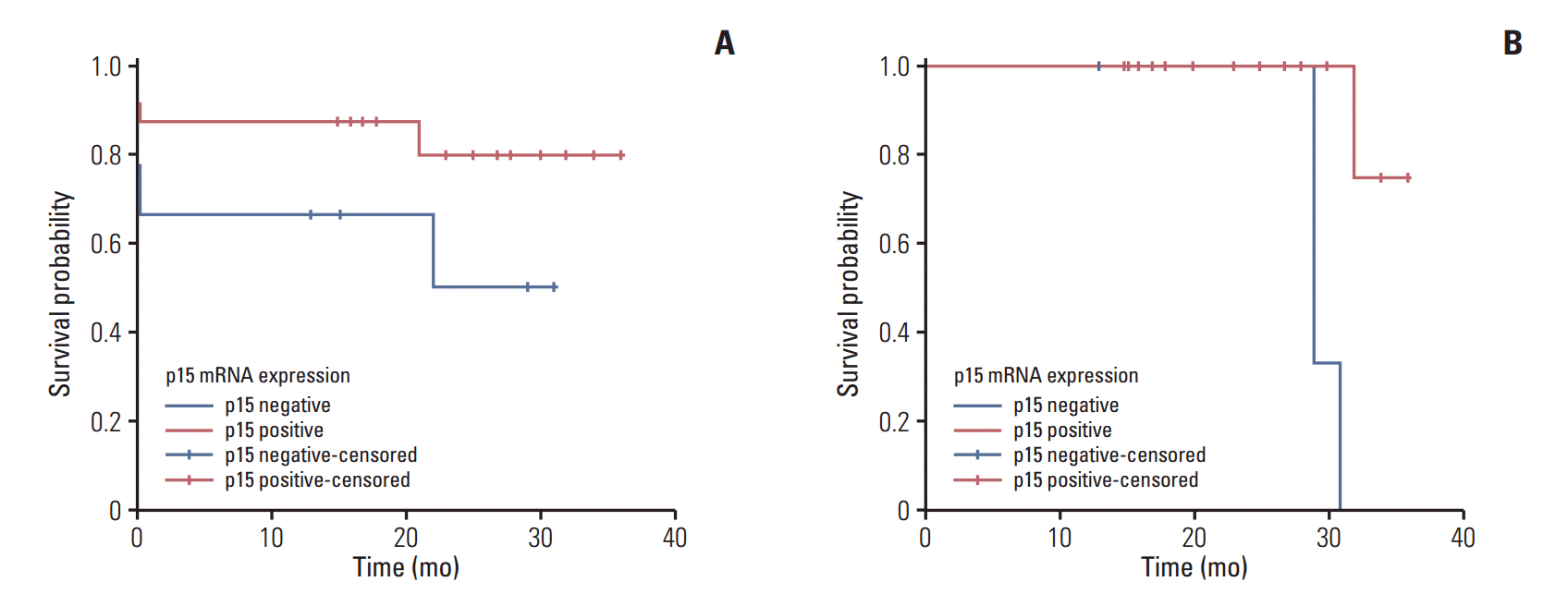
An OS of 38% and 91% was observed in p15-negative and p15-positive patients, respectively. Patients who did not express p15 mRNA showed a lower mean OS of 19.2 months as compared to 30.3 months in patients who expressed p15 mRNA. The difference in OS between two groups was significant based on the log-rank value (p=0.031) (Fig. 3A). The DFS for p15-negative patients was lower than that of p15-positive patients (22% and 89%, respectively; log-rank test p=0.004), with a mean DFS time of 29.6 months for p15-negative and 34.0 months for p15-positive patients (Fig. 3B).
Go to :
Discussion
Promoter hypermethylation of p15Ink4b causes its silencing almost exclusively in cancers of the hematopoietic system, and is observed in acute leukemias of myeloid (AML) and lymphoid origins [11]. Aberrant hypermethylation occurs at the CpG islands of the gene, which extend throughout the promoter region, exon 1, and part of intron 1 [12]. Various studies have reported hypermethylation of p15 with a high frequency in all FAB subtypes of AMLs [13]. However, few studies have been conducted to specifically investigate p15 methylation in APL. In our study, which is first from India, we investigated the pattern of p15 promoter hypermethylation in APL patients and its subsequent expression by RT-PCR. The results revealed overall hypermethylation of the p15 gene in 45% of APL patients, which is marginally in agreement with the results of a previous study conducted by Teofili et al. [14], who reported that 52% of cases involved hypermethylation. Our results differ from another study by Chim et al. [15], who observed hypermethylation in 79% of cases. Aberrant methylation of p15Ink4b has been reported in up to 80% of patients with primary and secondary AML [16,17]. Although these studies vary slightly in terms of the frequency of p15 hypermethylation, they all indicate that this gene is actively involved in the leukemogenesis of APL. Moreover, aberrant. p15Ink4b methylation levels have been associated with a generally poor prognosis in many forms of the disease. In contrast to Teofili et al. [14], who found no clinical variables showing any significant association with the pattern of methylation, our study noted a significant association of p15 hypermethylation with the gender of the APL patients (p=0.021) and leukocyte count at baseline (p=0.031). Interestingly, all nine patients (100%) who presented with leukocytosis at baseline (> 1×1010/L) were found to harbor p15 gene methylation (p=0.031). These findings differ from those reported by Teofili et al. [14], who found p15 gene methylation in 58% of patients. Therefore, this study points towards a strong link between leukocytosis, which is generally considered a poor prognostic indicator in APL [18,19], and the pattern of p15 methylation in APL cases.
Apart from complete methylation (M group) and absence of methylation (U group), 19% of patients showed partial methylation of p15 (U/M group). A similar pattern was observed by Teofili et al. [14], although they reported a partial methylation pattern in more patients (41.0%) than in our study. Partial methylation of the p15 gene is expected to cause incomplete transcription of in which only the unmethylated part contributes to the expression or mRNA formation. Therefore, p15 mRNA expression was evaluated by semi quantitative RT-PCR in all patients to explore the pattern of methylation and silencing of the p15 gene.
All the cases from group M patients, who reported complete methylation (9/9), showed a lack of p15 mRNA expression. Additionally, complete expression was observed in patients having fully unmethylated DNA. In the group U/M patients, who had partially methylated DNA, a differential pattern of p15 mRNA expression was observed as shown by the differential intensity of RT-PCR bands for mRNA. All seven patients with partial methylated DNA showed low intensity mRNA bands against those patients with no methylation and/or the control.
The loss of p15 expression in group M confirms the findings of the previous study by Teofili et al. [14]. All patients who were fully methylated showed loss of expression, which differs from the results of a study by Preisler et al. [20], who found that the presence of p15 methylation in leukemic cells is not always associated with the lack of p15 expression and that p15 expression is not always observed in the absence of methylation. p15 loss of expression was observed in nine patients with leukocytosis who had already been found with complete methylation (p=0.031), which is in stark contrast with the results of a previous study [14] in which leukocytosis did not show a significant relationship with methylation or expression of p15.
The prognostic and clinical significance of p15 promoter hypermethylation and subsequent gene silencing caused by it was evaluated in terms of the three year OS and DFS durations by the Kaplan-Meier method. Comparisons of the OS and DFS between patient groups were based on the log-rank test. There was no significant difference between the OS of M, U/M, and U groups patients (38%, 100%, and 90%, respectively; log-rank test: p=0.295), with patients from group M showing the lowest survival of 19.2 months (Fig. 2A). Significant DFS (log-rank test p=0.022) (Fig. 2B) was associated with different patterns of p15 gene hypermethylation in patients who achieved clinical remission. Group M patients showed a lower DFS (22%) than groups M/U and U patients (83% and 93%, respectively). Similar results were reported by Teofili et al. [14], who found a significant 5 years DFS rate of 29%, 64%, and 79% for M, M/U, and U patients respectively, with OS values of 69%, 71%, and 85%, respectively. These findings are concordant with the OS values of 38% and 90% observed for groups M and U, respectively, in the present study. With regard to the U/M group, our OS was 100% when compared to the corresponding OS of 71% of Teofili et al. [14]. These findings can be attributed to the smaller sample size and lesser follow-up duration of our study. Further, our finding of decreased overall and DFS in APL patients with complete methylation is similar to the results reported by Chim et al. [15], who reported a high incidence of p15 methylation in patients with APL and demonstrated that the 5-year DFS of patients with abnormal methylation of p15 was significantly inferior to that of patients without p15 methylation.
A significant 3-year OS of 38% and 91% (log-rank p=0.031) (Fig. 3A) and DFS of 22% and 89%, respectively (log-rank p=0.004) was observed in p15-negative and p15-positive patients, respectively (Fig. 3B). These results are in agreement with those reported by Teofili et al. [14], who reported significant 5-year estimated DFS values (24% and 66% for p15-negative and -positive patients, respectively). However, they found no significance difference in the OS of p15-negative and positive patients (OS, 74% and 85%, respectively). Our findings are also in agreement with many previous studies of patients with all AML FAB subtypes, in which individuals without p15Ink4b hypermethylation at diagnosis had increased complete remission rates that also correlated with increased survival times [21,22]. While our study indicates p15 as a valid potential prognostic marker in APL, these findings need to be validated in larger series of samples.
We concluded that hypermethylation of p15 gene coupled with its loss of mRNA expression causes susceptibility to patients with APL which leads to their decreased survival. Thus, p15 promoter hypermethylation is a prospective prognostic indicator and a reliable clinical aid in assessment of patients with APL.
Go to :
 XML Download
XML Download