

 PDF
PDF Citation
Citation Print
Print


Introduction
Thyroid cancer, the most common malignant tumor of the endocrine organs, has shown steadily increasing morbidity in recent decades [1]. Papillary thyroid carcinoma (PTC) is the main form of nonmedullary thyroid cancer, accounting for approximately 80% of all thyroid cancers [2]. In addition to genetic factors and environmental exposure, recent studies have found that epigenetic alteration may also play an important role in development of variety types of tumors including thyroid cancer. In this regard, involvement of long noncoding RNAs (lncRNAs), particularly BRAF-activated long noncoding RNA (BANCR), papillary thyroid cancer susceptibility candidate 3 (PTCSC3), and noncoding RNA associated with mitogen-activated protein kinase pathway and growth arrest (NAMA), in regulation of thyroid cancer cell survival and apoptosis has been reported [3-5].
LncRNAs utilize a variety of mechanisms in regulation of gene expression including chromatin modification, transcriptional, or post-transcriptional modulation. Through binding to nuclear protein such as polycomb enhancer of zeste homolog 2 (EZH2), a histone methyltransferase, lncRNAs modify histone and by this mechanism, lncRNAs regulate expression of target genes [6,7]. Cis- (stimulate) and trans-regulation (repress) are two main types of transcriptional regulation, under which lncRNAs can target genomically local and distant genes, respectively. Post-transcriptional regulation by lncRNAs includes mRNA splicing, editing, trafficking, translation, and degradation.
Thyroid stimulating hormone (TSH), also known as thyrotropin, is a pituitary hormone that stimulates the thyroid through binding to its receptor (TSHR). TSHR is found mainly on thyroid follicular cells and aberrant expression of TSHR may be involved in development of thyroid cancer as well as Grave’s disease [8-11].
While altered expression of lncRNAs, including BANCR, PTCSC3 and NAMA, has been reported in a variety of cell types of thyroid cancer [3-5], the role of these lncRNAs in modulating TSHR expression, has not been reported. Therefore, the current study was designed to investigate expression of BANCR, PTCSC3, and NAMA in thyroid cancer tissues, and the role of these lncRNAs in regulation of TSHR in a thyroid cancer cell line. Here, we report that three lncRNAs, BANCR, PTCSC3, and NAMA, were differentially expressed in PTC tissue and its adjacent normal tissue. BANCR-knockdown resulted in significantly suppressed proliferation of PTC-derived cell line, IHH-4. In addition, BANCR was enriched by EZH2 and interference of BANCR led to decreased chromatin recruitment of EZH2 and resulted in significant reduction in TSHR expression.
Materials and Methods
1. Cell culture
A human PTC-derived cell line, IHH-4, was used for in vitro cell culture experiments. IHH-4 cell line was cultured with RPMI1640 medium supplemented with 10% fetal calf serum at 37°C and 5% CO2 atmosphere. Passages 5 to 10 were used in the current study.
2. Transfection of cells with BANCR siRNA
Transfection was performed following the manufacturer’s instruction. Briefly, IHH-4 cells were grown to 75% confluency on 60-mm dishes in RPMI1640 supplemented with 10% fetal calf serum. Lipofectamine-2000 (24 μL; Life Technology, Grand Island, NY) was mixed with 400 μL of serum- and antibiotic-free Opti-MEM (Life Technology) for each dish transfected, and allowed to sit at room temperature for 5 minutes. Simultaneously, 48 μL of each siRNA (GE Dharmacon, Lafayette, CO) was mixed with 800 μL of Opti-MEM (for each dish) and likewise incubated. The two suspensions were then combined and incubated for an additional 20 minutes at room temperature. After washing cell layers gently with phosphate bufferd saline (PBS), 1.2 mL of the mixture with the addition of 0.8 mL Opti-MEM was added to each dish, followed by incubation overnight (18 hours) at 37°C and 5% CO2. Cells were then fed normal growth medium and used for experimentation as designed. Non-targeting control siRNA (GE Dharmacon) was used as a negative control.
3. Cell proliferation assay
Cell proliferation was assessed using a Cell Counting Kit-8 (CCK-8; Beyotime Institute of Biotechnology, Jiangsu, China). Briefly, 24 hours after transfection with control-siRNA or BANCR-siRNA, cells were trypsinized and plated in triplicate wells of a 96-well plate at a density of 2×103 cells/well. Cells were allowed to grow in RPMI1640 supplemented with 10% fetal calf serum for 5 days. The number of cells per well was determined by the absorbance at 450 nm at indicated time points, following the manufacturer’s instructions. Data were expressed as value of optical density at 450 nm wave length.
4. Clonogenic assay
Clonogenic assay was performed with a modification of a previously published method [12]. Briefly, after completion of siRNA transfection, the cells were plated into 60-mm tissue culture dishes in triplicate and at a cell density of 100 cells/dish. Cells were then allowed to grow in RPMI1640 supplemented with 10% fetal calf serum for 14 days. Cells were then fixed and stained with Diff-Quick. After air drying, colonies were counted under a microscope and cell number in each colony was at least 20 cells.
5. Cell cycle analysis by flow cytometry
After transfection with siRNA, cells were cultured in RPMI1640 supplemented with 10% fetal calf serum for an additional 48 days. Cells were then trypsinized and suspended with RPMI1640 containing 10% serum. Cells were then pelleted, suspended with cold ethanol/PBS at a ratio of 2.5:1 and fixed at 4°C for at least 30 minutes. Cells were then stained with propidium iodide solution for 30 minutes as described previously [13]. Cell cycle and DNA content were then analyzed by flow cytometry.
6. Quantitative real time reverse transcription polymerase chain reaction
Total RNA from cells or tissue was extracted using Trizol (Life Technology). After RNA quantification with Nanodrop 1000 (Thermo Scientific, Wilmington, DE), reverse transcription was performed using a commercially available kit (Applied Biosystem, Life Technology, Grand Island, NY) following the manufacturer’s instructions. Quantitative real time polymerase chain reaction (PCR) was then performed using commercially available pre-synthesized probe/primers and reagents, and the 7500 Real Time System (Applied Biosystems, Life Technology) following the manufacturer’s instructions. Each reaction was performed in triplicate, and the 2-ΔΔCt method was used to determine the relative gene expression levels. As an internal control, 18s RNA was used to normalize the data.
7. Immunoblotting
Proteins of total cell lysate or proteins from the tissue were extracted using RIPA lysis and extraction buffer. After brief sonication and centrifugation, protein concentration was determined using a Bio-Rad protein assay kit (Bio-Rad, Hercules, CA). A total of 10 μg protein was electrophoresed on a 10% sodium dedecyl sulfate polyacryamide gel and subsequently transferred to a polyvinylidene difluoride membrane (Bio-Rad). The membrane was blocked with 5% skimmed milk in Tris-buffered saline containing 0.1% Tween-20 (TBST) at room temperature for 1 hour. The membrane was then incubated with the desired primary antibodies (1:1,000 dilution) at 4°C overnight. The next day, the membrane was washed 3 times with TBST, followed by incubation with horseradish peroxidase-conjugated secondary anti-rabbit or anti-mouse antibody (1:4,000, Rockland, Limerick, PA) at room temperature for 1 hour. After washing 3 times with TBST, the membrane was developed using ECL Plus (EMD Millipore, Billerica, MA), and exposed to X-ray film. β-Actin was used as an internal loading control.
8. Chromatin immunoprecipitation assay
Chromatin immunoprecipitation (ChIP) assay was performed using an EZ ChIP Chromatin Immunoprecipitation Kit (Millipore, Bedford, MA) following the manufacturer’s instructions. Briefly, cross-linked chromatin was sonicated into 200-bp to 1,000-bp fragments. The chromatin was immunoprecipitated using anti-EZH2 antibody (Cell Signaling Technology, Beverly, MA). Normal mouse IgG was used as a negative control. Quantitative PCR was then performed using SYBR Green Mix (Takara Bio, Otsu, Japan).
9. RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) assay was performed using a Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore) following the manufacturer’s instructions. The anti-EZH2 antibody and IgG (as control) was used for RIP (Cell Signaling Technology). Co-precipitated RNAs were detected by quantitative reverse transcription polymerase chain reaction (RT-PCR). Total RNAs (input controls) and isotype controls were assayed simultaneously to demonstrate that the detected signals were the result of RNAs specifically binding to EZH2.
10. Tissue sample collection
With informed consent, a total of 40 pairs of specimens of human PTC and adjacent normal tissue were obtained from surgical procedures performed at Yantai Yuhuangding Hospital from September 2013 to November 2014 (Yantai, Shandong, China). The protocol of the current study was approved by the Protection of Human Ethics Committee, College of Medicine, Qingdao University (Qingdao, Shandong, China). PTC diagnosis was pathologically confirmed, tissues were snap frozen with liquid nitrogen, and stored at –80°C till RNA extraction.
11. Statistical analysis
Data were analyzed using PRISM 6 software (GraphPad, La Jolla, CA), and expressed as mean±standard deviation of the data from at least three separate experiments. Paired data were analyzed by Student’s t test, and group data were analyzed by one-way analysis of variance (ANOVA) followed by Tukey correction, as appropriate. p < 0.05 was considered significantly different in comparison.
Results
1. Differential expression of three lncRNA in PTC
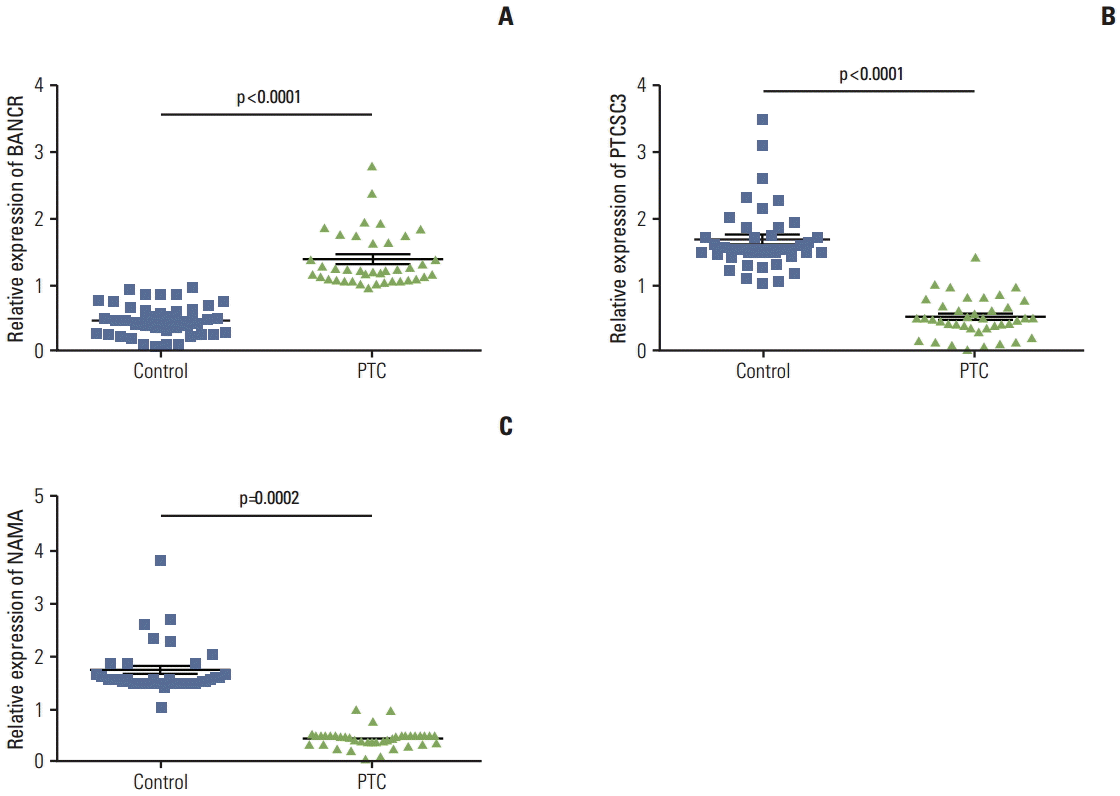
A total of 40 pairs of PTC tissue and its adjacent normal tissue were obtained from clinically ongoing surgical specimens with informed consent. Cellular types of the PTCs were PTC with poorly differentiated area (6 cases), follicular variant (15 cases), diffuse sclerosing variant (8 cases), and CP classical PTC (21 cases). To examine the role of lncRNAs in the pathogenesis of thyroid carcinoma, expression of three lncRNAs, BANCR, PTCSC3, and NAMA in these tumor tissues as well as their adjacent normal tissues was assessed by quantitative real-time RT-PCR. Interestingly, BANCR expression was significantly up-regulated in PTC compared to that in its adjacent normal tissue (p < 0.0001) (Fig. 1A). In contrast, expression of PTCSC3 and NAMA was significantly down-regulated in PTC compared to that in normal tissue (p < 0.0001 and p=0.0002, respectively) (Fig. 1B and C).
2. Effect of lncRNA suppression on TSHR expression in in vitro cell culture
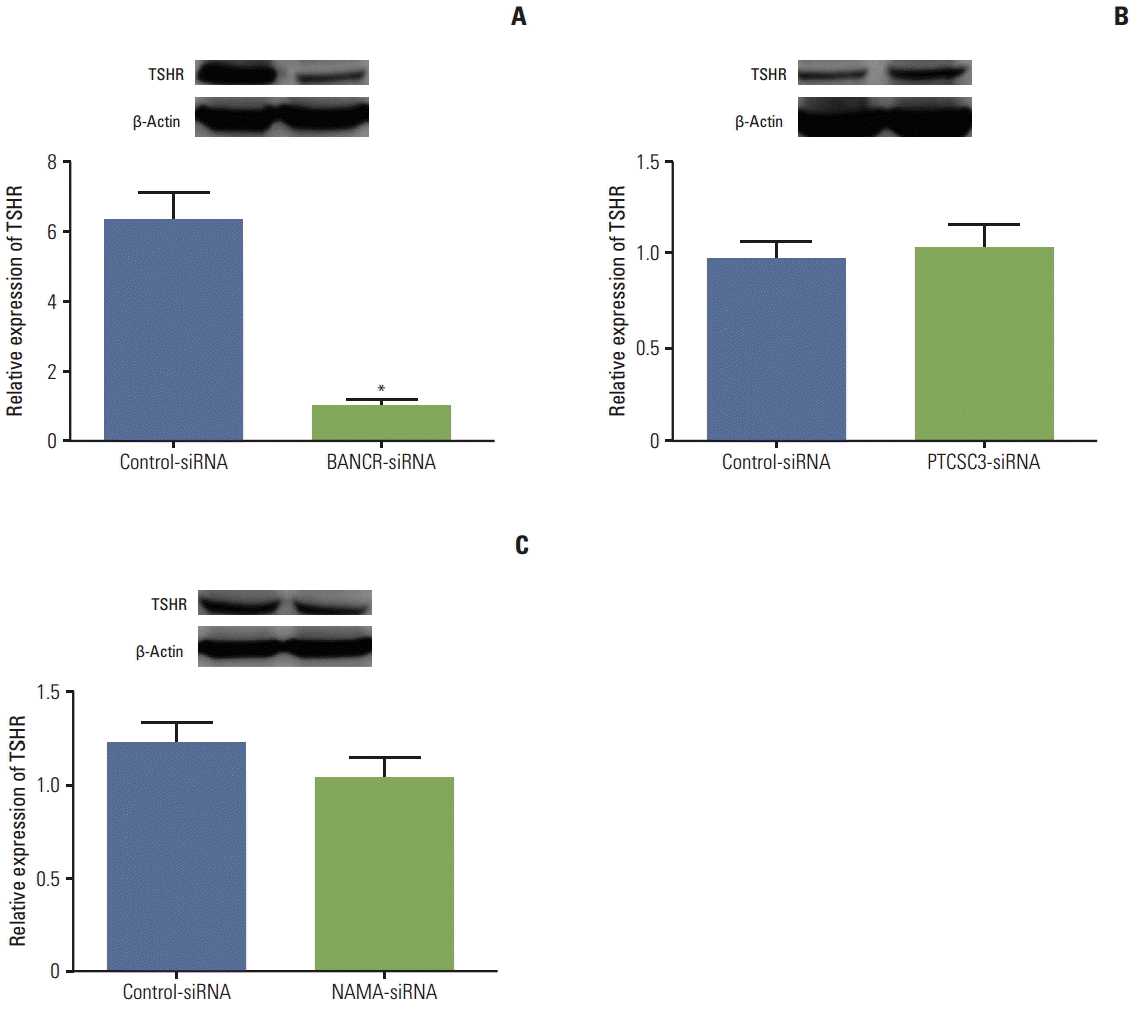
Since thyroid cell proliferation and function is mediated by the interaction of TSH and its receptor (TSHR), the effect of silencing lncRNA on TSHR expression was investigated using a cell line of PTC, IHH-4 cells. For suppression of BANCR, PTCSC3, and NAMA, siRNAs specifically targeting BANCR, PTCSC3, or NAMA were transiently transfected. Efficiency of lncRNA silence was determined by real-time RT-PCR and at least 60% suppression was demonstrated (data not shown). Silencing BANCR resulted in significant down-regulation of TSHR (p < 0.01) (Fig. 2A), while neither PTCSC3-siRNA, nor NAMA-siRNA had a significant effect on TSHR expression in IHH-4 cells (Fig. 2B and C, respectively).
3. BANCR-siRNA inhibited cell proliferation
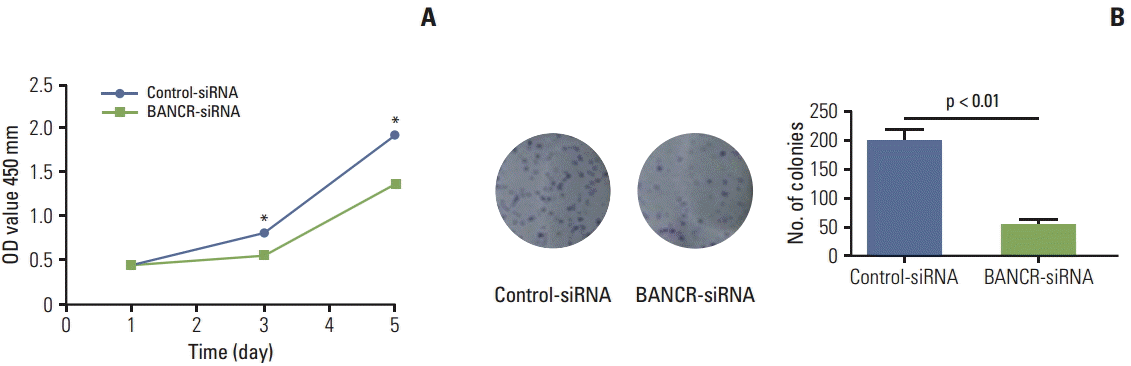
Following transfection of BANCR-siRNA, IHH-4 cells were allowed to grow in full medium for 5 days. Growth of cells transfected with BANCR-siRNA was significantly slower than that of cells with control-siRNA on day 3 and day 5 (p < 0.05 and p < 0.01, respectively) (Fig. 3A). A clonogenic assay was performed to confirm the effect of BANCR suppression on cell proliferation. Consistent with the result of cell proliferation assay, number of clones was significantly decreased in cells lacking BANCR (200.3±25.5 of controlsiRNA vs. 56.7±8.5 of BANCR-siRNA, p < 0.01) (Fig. 3B).
4. BANCR modulated the cell cycle via regulation of cyclin D1
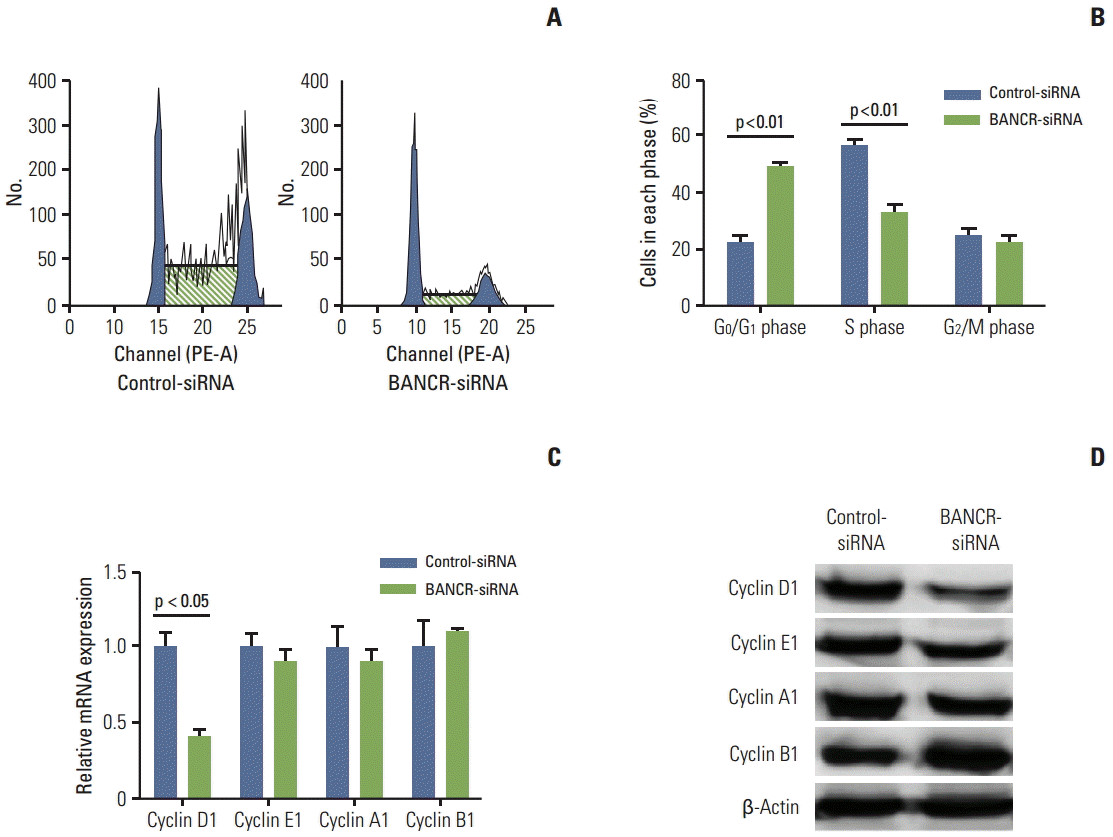
To further investigate the role of BANCR in regulating cell proliferation, cellular DNA content and cell cycle profiling was performed. As shown in Fig. 4A and B, G1 to S phase transition was significantly decreased. Gene expression and protein level of cyclin proteins including cyclin D1, cyclin E1, cyclin A1, and cyclin B1 were also determined following BANCR-siRNA transfection into IHH-4 cells. Of particular interest, cyclin D1 mRNA and protein was significantly decreased in cells transfected with BANCR-siRNA compared to that of control-siRNA transfected cells (p < 0.05) (Fig. 4C and D). In contrast, mRNAs of cyclin A1, E1, or B1 were not altered in cells after silencing BANCR (Fig. 4C), although proteins of cyclin A1 and E1 were slightly but not significantly reduced in cells lacking BANCR (Fig. 4D).
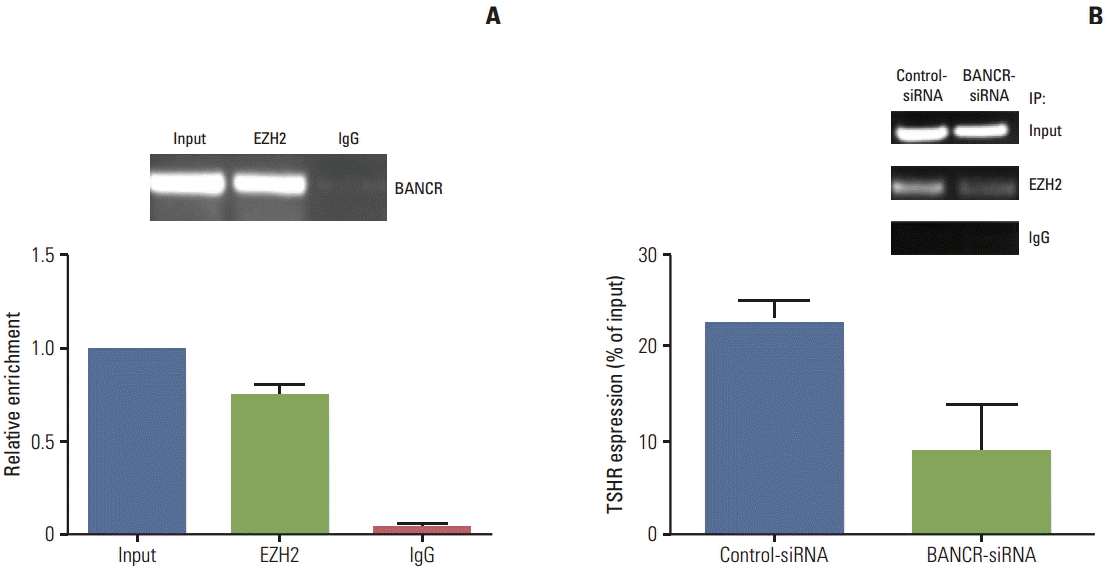
5. BANCR is associated with EZH2 and might regulate cancer cell growth
EZH2 is known to inhibit genes responsible for suppressing tumor growth, and lncRNAs modulate tumor cell growth through binding to EZH2 in variety of cancer cells [14,15], therefore, association of BANCR and EZH2 was further investigated in the current study. First, RIP assay was performed using anti-EZH2 and IgG as a negative control. Then, quantitative RT-PCR was performed for quantification of the retrieved RNA. As shown in Fig. 5A, BANCR was highly enriched in the EZH2 group compared to the non-immune IgG group. Next, ChIP assay was performed to further determine whether BANCR knockdown altered expression of TSHR. As shown in Fig. 5B, expression of TSHR was significantly decreased in BANCR-knockdown cells compared to control cells, suggesting that BANCR may enhance TSHR expression through binding to EZH2.
Discussion
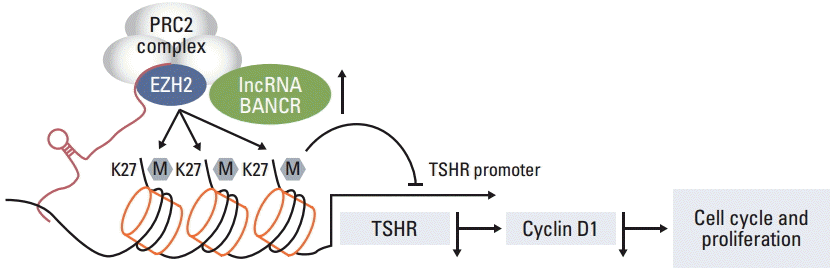
Accumulating evidence suggests that lncRNAs may play an important role in development of various types of cancer. PTC is the most common type of thyroid cancer. In the current study, we first examined expression of three lncRNAs in the PTC and its adjacent normal tissue. It was found that BANCR was significantly up-regulated in PTC tissue compared to normal tissue, and in contrast, expression of PTCSC3 and NAMA was significantly decreased in tumor tissue. Using in vitro cell culture and molecular strategy, we further demonstrated that BANCR regulated expression of TSHR, while neither PTCSC3 nor NAMA had an effect on TSHR expression. BANCR controlled growth and cell cycle of IHH-4 cells (a cell line of PTC) through regulation of cyclin D1 protein expression. In addition, physical binding of BANCR to EZH2 was demonstrated by RIP assay, and by this mechanism, BANCR may regulate TSHR as well as subsequent target genes including cyclin D1 (Fig. 6).
PTC is the main form of nonmedullary thyroid cancer, accounting for approximately 80% of all thyroid cancers. In contrast to other cancers, the incidence of PTC has increased in recent decades, but this may due to improved diagnostics [2]. Although the etiology of PTC is not well defined, it is clearly influenced by genetic factors and environmental exposures. Radiation exposure of the thyroid during childhood is the most clearly defined environmental factor. However, genetic predisposition also plays an important role as evidenced by case control studies [16]. In addition, recent accumulating evidence indicates that epigenetic alteration may also play an important role in the development of various kinds of tumors including thyroid cancers. In this context, involvement of lncRNAs, particularly BANCR, PTCSC, and NAMA, in regulation of thyroid cancer cell survival and apoptosis has been reported [3-5].
Noncoding RNAs are a group of RNAs that do not code for protein synthesis. They are categorized according to two classes based on transcript size: “long noncoding RNAs (lncRNAs)” are greater than 200 nucleotides in length, and “small noncoding RNAs” are less than 200 nucleotides in length including microRNAs [17,18]. Unlike miRNAs, which have been studied extensively in the last decade, lncRNAs are more diverse, not highly conserved, and have been regarded as transcriptional “noise” and not biologically functional for a long period of time [19,20]. Recently, however, accumulating evidence has revealed the significance of lncRNAs in regulation of multiple biological functions at all levels of chromosome modification, mRNA transcription, and post-transcriptional protein synthesis [21-23]. Notably, several lncRNAs including BANCR, PTCSC3, and NAMA have been reported to play an important role in regulating thyroid cancer formation and progression [3-5]. Consistent with previous reports, we found that expression of BANCR was significantly up-regulated in PTC compared to that in normal tissue, while expression of PTCSC3 and NAMA was dramatically down-regulated in PTC compared to its adjacent normal tissue, suggesting that thyroid carcinogenesis is positively regulated by BANCR and negatively regulated by PTCSC3 and NAMA.
In the current study, biological function of BANCR in thyroid cancer was further investigated utilizing an in vitro IHH-4 cell culture. IHH-4, a cell line derived from PTC, has been used by other investigators in study of thyroid cancer biology [3]. Consistent with findings in human PTC tissue, IHH-4 cells also highly express BANCR (data not shown). In this study, silencing BANCR with siRNA in IHH-4 cells resulted in remarkable suppression of cell proliferation, and cell cycle arrest at G0/G1 phase. In addition, significant down-regulation of both mRNA and protein of cyclin D1 was observed in IHH-4 cells lacking BANCR. Paradoxically, mRNA of cyclin A1, E1, and B1 was not altered in cells transfected with BANCR-siRNA, although proteins of cyclin A1 and E1 were slightly but not significantly reduced in cells lacking BANCR, suggesting that BANCR regulated cell growth and cell cycle through modulation of cyclin D1.
TSH exerts its effect on thyroid cell proliferation through binding to its receptor, TSHR. TSHR is mainly found on thyroid follicular cells. According to some reports, pathogenesis of thyroid cancer and Grave’s disease may be attributed to the aberrant expression of TSHR [8-11]. While we were unable to examine expression of TSHR in PTC tissues due to lack of sources, which is a pitfall of the current study, we demonstrated that BANCR-knockdown in IHH-4 cells resulted in a significant reduction in the TSHR protein. Interestingly, however, TSHR protein synthesis was not significantly altered by either PTCSC3-siRNA or NAMA-siRNA, although TSHR protein was slightly decreased in cells transfected with NAMA-siRNA, indicating that BANCR may contribute to development of PTC through regulation of TSHR, but not PTCSC3 or NAMA.
Tumorigenesis and progression of cancer result not only from genetic factors and environmental exposure, but also from epigenetic alteration. Epigenetic modifications include DNA methylation, histone modifications, and modulation by miRNAs or lncRNAs. Histone modification has been reported to play a role in the tumorigenesis of thyroid cancer, and EZH2, a histone methyltransferase, is a well-known histone modifier. In this regard, researchers have demonstrated a strong association of overexpression of EZH2 with progression and invasion of several types of cancers including thyroid cancer [7,14,15]. In addition, studies have shown that EZH2 was up-regulated by aberrant expression of MYC transcription factor and miRNAs such as miR-26a [24,25]. Interaction of BANCR and EZH2 in the tumorigenesis of thyroid cancer, however, has not been reported. The results of the current study demonstrated that BANCR was enriched by EZH2, and that silencing BANCR by siRNA resulted in decreased chromatin recruitment of EZH2 as well as significant reduction in TSHR expression as evidenced by ChIP assay with anti-EZH2 antibody, suggesting that BANCR could stimulate TSHR expression by transcriptional up-regulation of the TSHR promoter through EZH2 enrichment. The current study, however, did not determine whether histone methylation in the promoter region of cyclin D1 was affected by the suppression of BANCR and reduced chromatin recruitment of EZH2, which is a limitation of the current study and remains to be defined in future study.
Taken together, the three lncRNAs tested in the current study were differentially expressed in PTC tissue and a PTC-derived cell line (IHH-4). Specifically, BANCR was significantly up-regulated in PTC, and silencing BANCR with siRNA resulted in dramatically reduced expression of TSHR as well as suppression of cell proliferation via a mechanism of cell cycle arrest. In addition, BANCR was enriched by EZH2 and BANCR was required for EZH2 binding to TSHR promoter because interference of BANCR abrogated chromatin recruitment of EZH2 and resulted in significant reduction of TSHR expression. These findings suggested regulation of BANCR as a therapeutic goal of PTC.
Conclusion
The current study demonstrated that BANCR was significantly up-regulated in PTC, and silencing BANCR with siRNA resulted in dramatically reduced expression of TSHR as well as suppression of cell proliferation via a mechanism of cell cycle arrest. These findings indicated that regulation of BANCR be a therapeutic strategy for PTC.
 XML Download
XML Download