

 PDF
PDF Citation
Citation Print
Print


Introduction
For many human cancers, dysregulation of cellular metabolism is now recognized as a key event in tumor growth and development. Accordingly, research into various aspects of tumor metabolism is undergoing a period of sustained revival, with the hope that this will identify novel tractable pathways for cancer screening, diagnosis and the treatment of malignant disease [1]. Indeed, the development of antimetabolites, such as antifolates and pyrimidine analogues, and the application of Warburg's hypothesis in 18F-fludeoxyglucose positron emission tomography/computed tomography (18F-FDG PET/CT) as a diagnostic and therapeutic imaging tool, demonstrate the importance of studying cancer metabolism in the oncology clinic: continued dissection of the cancer metabolome is likely to further refine our management of patients with cancer. Testing for metabolic genetic drivers such as mutations in Kreb's cycle genes (i.e., fumarate hydratase [FH], succinate dehydrogenase [SDH], isocitrate dehydrogenase-1/2 [IDH 1/2]) is already available, and new PET tracers and antimetabolites are being devised that aim to exploit the differential biochemistry of sugars, lipids and amino acids in normal and malignant cells [2,3]. Here, the focus is on the amino acid L-arginine, which has garnered clinical interest as an exciting cancer target, driven by the emergence of several novel arginine-depleting agents.
Amino Acid Deprivation as an Anticancer Therapeutic Strategy
The rationale for targeting amino acids in cancer therapy was pioneered with the introduction of L-asparaginase in childhood leukemia over five decades ago [4]. In contrast to normal cells, leukemic cells are dependent on exogenous asparagine for growth (i.e., auxotrophic), due to lack of the biosynthetic enzyme asparagine synthetase [5]. As a monotherapy, deamidation of asparagine to aspartate and ammonia by L-asparaginase, led to responses of up to 50% in early studies of patients with chemorefractory leukemia [6]. However, it was not until the advent of multimodality chemotherapy regimens containing L-asparaginase, that the cure rate of acute lymphoblastic leukemia increased from 5% in the 1950's to the present 90%. The problem of hypersensitivity reactions and anti-drug neutralizing antibodies has been addressed to some extent by use of pegylation technology, switching to alternative L-asparaginase enzymes and individualized schedules with close monitoring of plasma asparagine levels [7]. Nevertheless, wider clinical investigation of L-asparaginase, has been hampered by the risk of severe dose-limiting side effects including hepatitis, pancreatitis, coagulopathy and neurotoxicity [8]. Bioengineering efforts are underway to modulate the secondary glutaminolytic activity that mediates, in part, the adverse effects but also the antitumor properties of L-asparaginase [9].
Based on the asparaginase paradigm, several argininecatabolizing enzymes have been developed, including mycoplasma-derived arginine deiminase and human recombinant arginase, for cancer therapy [10]. These enzymes are pegylated for enhanced pharmacokinetics and pharmacodynamics; however, further drug optimization will still be needed for arginine deprivation to gain traction in the clinic [11]. Although the data are preliminary, early signs suggest that arginine deprivation has the potential to make an impact across a broad range of human malignancies.
Arginine Cancer Metabolome: Argininosuccinate Synthetase 1 (ASS1) and Argininosuccinate Lyase (ASL) Enzymes
On an organismal level, diet and protein degradation are the main sources of plasma arginine, with only 5-15% (up to 30% in newborns) derived from de novo biosynthesis via the intestinal-renal axis. Indeed, under physiological conditions the arginine biosynthetic pathway is organized according to tissue function: for example, as the urea cycle in the liver for elimination of nitrogenous waste, and as the nitric oxide (NO)-citrulline cycle in endothelial cells to generate NO, with arginine acting as an intermediary molecule [12].
Numerous studies have confirmed that ASS1, considered a house-keeping gene in normal cells and a rate-limiting biosynthetic enzyme in hepatocytes and endothelial cells, is absent in many tumors [13,14]. Importantly, this leads to a critical dependence on exogenous arginine, with evidence suggesting that tumoral ASS1 deficiency may be both a prognostic biomarker and predictor of sensitivity to arginine deprivation therapy [15]. Methylation-dependent transcriptional silencing has been shown to drive tumoral ASS1 loss in some tumor types, although other mechanisms may also be involved, including repression of the ASS1 promoter by hypoxia-inducible factor-1alpha detected in melanoma cells [16,17]. Further study of the mechanisms of ASS1 loss and sensitivity to arginine depletion will be critical in overcoming resistance in the clinic.
The reason for down-regulation of tumoral ASS1 expression remains unclear and is an area of ongoing research. Recent studies have shown that ASS1 functions as a tumor suppressor in sarcoma and bladder cancer cell lines, reducing colony forming ability, proliferation and invasion of tumor cells and abrogating growth of tumor xenografts in mouse models [18-20]. Indeed, it appears that the reprogramming of tumor arginine metabolism by deactivating ASS1 favors a more aggressive phenotype fueled by exogenous arginine. Previously extracellular arginine has been shown to increase DNA synthesis in Burkitt lymphoma cells, and enhance incorporation of glutamine into nucleotides in colorectal cancer cells [21,22]. Work from our laboratory supports the hypothesis that exogenous arginine is sparing for glutamine in the synthesis of pyrimidines, specifically in ASS1-negative tumor cells. Thus, arginine depletion in ASS1-negative cells led to a marked increase in intracellular glutamine whilst thymidine levels declined secondary to inhibition of thymidylate synthase and dihydrofolate reductase, highlighting the interdependence between exogenous arginine and glutamine for nucleotide synthesis. In contrast, thymidine levels were unaffected by arginine withdrawal in ASS1-positive tumor cells, suggesting that glutamine is being diverted towards arginine synthesis, thereby limiting the capacity for proliferation and growth [20]. Evidence for crosstalk between glutamine and arginine has also been revealed in studies of melanoma cells exposed to ADI-PEG20 [23]. Moreover, metabolomic studies in sarcomas have identified arginine along with glutamine and serine, as essential amino acids with a signature of enhanced gemcitabine sensitivity in a subgroup of liposarcomas [24,25].
Notably, there is also evidence for an aberrant arginine metabolome in nasopharyngeal and bladder cancer cells with an inverse relationship between ASS1 and the expression of key serine pathway enzymes, including phosphoserine phosphatase (PSPH) and phosphoserine aminotransferase I (PSAT) [20,26]. Elevated levels of PSPH and PSAT have been identified in several tumors, and modulate one-carbon folate metabolism [27]. This is an area that will require detailed metabolomic studies to delineate the role of arginine given that there is increasing evidence for differential mechanisms involved in loss of ASS1 expression.
Less is known about the tumoral role of ASL, which is immediately downstream of ASS1 and catalyzes the conversion of argininosuccinate into arginine and fumarate. It is apparent from inborn errors of metabolism that ASL is critical in channeling arginine for NO production, with NO donors reversing the clinical picture of hypertension in children with ASL deficiency [28]. More recently, ASL has been explored in cancer, with upregulation noted in hepatocellular carcinoma (HCC) linked to increased aggressiveness mediated by NO and cyclin A2 signaling [29]. On the other hand, there is evidence that methylated ASL contributes to the arginine auxotrophy of glioblastoma multiforme, with loss of ASS1 and ASL conferring greater sensitivity to ADI-PEG20 [30]. In FH deficient renal cell cancer (RCC), where ASL is intact, the enzyme's activity is reversed producing high levels of argininosuccinate from arginine and fumarate. Here, arginosuccinate levels correlate with reversed ASL activity and may prove to be a biomarker in FH null RCC and other tumors [31,32]. Here too, the role of ASL in human cancer appears to be dependent on tumor type, and further work is needed to understand the implications of ASL expression in human cancer.
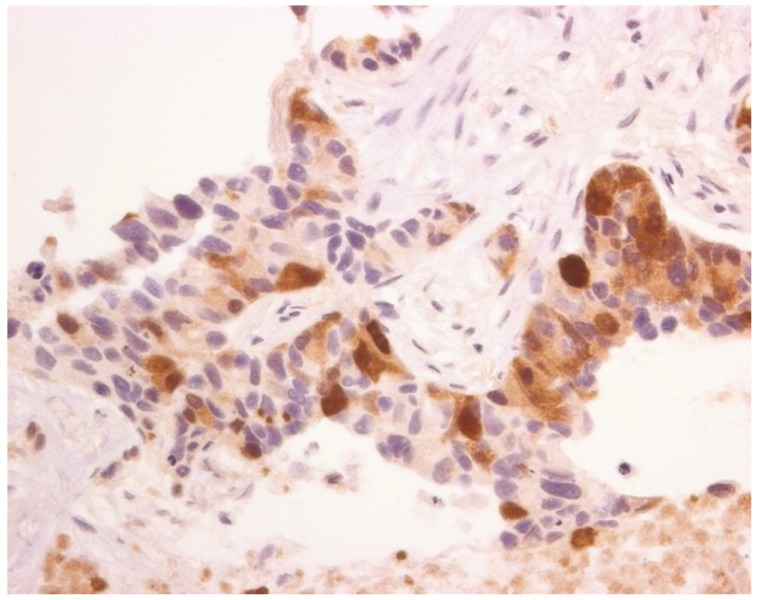
Lastly, tumoral heterogeneity of urea-cycle enzyme expression is likely to impact the development of arginine deprivation as a targeted therapy in human cancer. Concerning ASS1, whilst in some tumors, such as melanoma, lymphoma, and small cell lung cancer, there appears to be an 'all or none' level of expression, in the epithelial cancers, non-small cell lung carcinoma, mesothelioma and ovarian cancer, we have noted significant heterogeneity of expression (Fig. 1) [14,16,33-35]. As a result, moving forward into clinical trials we have opted for at least 50% ASS1 negativity within a tumor biopsy for patient selection, thus lowering the bar for patient inclusion, whereas previous trials in melanoma and small cell lung carcinoma (ClinicalTrials.gov Identifier: NCT01266018) have mandated at least 95% ASS1-negative cells.
Arginine Deprivation: Preclinical Data
Several decades of research have led to the notion that arginine promotes tumor growth: manipulation of dietary arginine in cancer cell line studies and in vivo transplantable tumor models; ex vivo analyses of cancer tissue arginase; and in vitro and in vivo pharmacological studies of native arginine-degrading enzymes [36-41]. More recent studies evaluating the efficacy of pegylated bioengineered enzymes have confirmed that arginine depletion is effective in various ASS1-deficient xenograft tumor models, including HCC, melanoma, pancreatic and small cell lung cancer and leukemia [34,42-44]. Whilst amino acid deprivation is known to induce nutritional starvation, the inhibitory effects of arginine depleting agents on tumor growth are not fully understood and remain an area of active investigation. In addition to the links between exogenous arginine and intracellular glutamine and pyrimidine synthesis described above-which impact directly on the cell cycle-ADI-PEG20 also downregulates mammalian target of rapamycin (mTOR) and modulates phosphoinositide 3-kinase (PI3K) via suppression of phosphatase and tensin homolog (PTEN) in ASS1-negative tumor cells [45-49]. It is noteworthy that arginine is one of two key amino acid regulators of mTOR-the other being leucine-and may explain, in part, the ability of arginine depletors to critically impair protein synthesis [50,51]. Apart from modulating tumor biomass via suppressing nucleotide and protein synthesis, arginine also affects tumor cell motility via a NO and focal adhesion kinase (FAK)-dependent pathway; importantly, FAK activation has been linked to deprivation of several other amino acids including glutamine and tyrosine [52-54]. Interestingly, arginine deiminase was identified in an anti-angiogenic screen and an anti-vascular effect has been confirmed both in HUVEC cells in vitro and in a xenograft melanoma study using fluorescence molecular tomography. Furthermore, arginine deiminase was antiangiogenic and potentiated the effect of radiation in a neuroblastoma mouse model [55-58]. Although some native arginine deiminases, found in micro-organisms such as Giardia lamblia, have functions beyond arginine depletion including citrullination of proteins (i.e., a peptidylarginine deiminase), this is not a feature of mycoplasmal-derived ADI-PEG20 used in clinical trials to date [59]. Clearly, arginine-lowering enzymes are likely to impact numerous other pathophysiological pathways and their further characterization will be crucial in optimizing arginine deprivation in the clinic [60-63].
Drug resistance remains a significant challenge for effective usage of arginine depletors. Despite promising results observed in vitro and in vivo as a targeted therapy for arginine auxotrophic tumors, several mechanisms have been identified that mediate tumoral resistance to arginine depletors. The most plausible is re-expression of ASS1, which results in the re-cycling of citrulline back into arginine [64]. This has been described for melanoma cells via transcriptional activation of the ASS1 promoter by the oncogene c-myc [17]. More recently, c-myc has been shown to play a role in the metabolic reprogramming seen during the development of ADI-PEG20 resistance in arginine auxotrophic melanoma cell lines. Here, ADI-PEG20-resistant cells exhibited elevated expression of key enzymes in glutaminolysis, glutaminase (GLS1) and glutamine dehydrogenase, and increased glycolytic metabolic activity, resulting in preferential sensitivity to glutamine inhibitors and glycolytic inhibitors, respectively. c-Myc, not elevated ASS1 expression, was found to contribute to this metabolic rewiring [17,23].
Autophagy ('self-eating'), leading to the provision of a temporary but finite supply of arginine via the breakdown of intracellular organelles, is also triggered by arginine deprivation, circumventing arginine deprivation in several malignancies, including prostate cancer, lymphoma, glioblastoma multiforme, and melanoma [16,30,65,66]. An alternative mechanism may involve macropinocytosis whereby extracellular proteins are degraded to supply the tumor cell with amino acids, as shown recently for K-ras transformed cells deprived of exogenous glutamine [67]. Finally, the direct supply of arginine or its precursors by the tumor microenvironment is also an attractive hypothesis. A similar hypothesis was first proposed and validated for mesenchymal-derived stromal cells providing asparagine for L-asparaginase-resistant acute lymphoblastic leukemia cells and, latterly, adipocytes supplying glutamine, consistent with the dual catalytic activity of L-asparaginase [68,69]. Evidence suggests that tumor associated macrophages (TAMs), predominant cells of the innate immune system within the tumor microenvironment, are involved in complex chemical cross-talk with tumor cells. TAMs are highly versatile and this interplay has the potential to modulate TAM phenotype, which we propose may in turn lead to a form of metabolic cooperation between TAMs and the tumor cells, resulting in the provision of arginine or its precursors to the tumor, thereby by-passing the effect of arginine deprivation. We have recently reviewed the complexity of the tumor microenvironment in relation to arginine, and it is clear that host cells may influence tumorigenesis by being sources of arginase as well as arginine [70-73]. Irrespective of the resistance mechanism, the final common pathway ends with arginine, as this is essential to the survival of the ASS1-negative tumor cell that is unable to supply sufficient amounts of arginine via upregulation of ASS1.
It is likely that further tumoral mechanisms involved in the efficacy and resistance of arginine depletors will be identified and this will be critical in exploiting arginine deprivation with rationally selected drugs for cancer therapy. Ultimately, this may improve the treatment efficacy of many cancers that depend on arginine for survival. To date several preclinical combinatorial studies have shown additive and/or synergistic effects of arginine depletors with taxanes, 5-fluorouracil, pemetrexed cytarabine, PI3K inhibitors, arginine mimetics (e.g., canavanine) and modulators of autophagy using chloroquine and TRAIL [20,44,45,74-77]. Additional interactions that merit further investigation include the potentiation observed between arginine deprivation and radiotherapy in 3D spheroid models of arginine auxotrophic cell lines [78]. Indeed, recent work indicates that ADI-PEG20 is synergistic in vitro and in vivo with radiotherapy in pancreatic cancer cells and may have utility in combination with gemcitabine, a known radiosensitizer and potentiator of ADI-PEG20 in pancreatic malignancy [79,80].
We have investigated further the effect of combining ADI-PEG20 with the antifolate drug pemetrexed based on the novel finding that ADI-PEG20 suppresses the pemetrexed target enzymes, thymidylate synthase (TS) and dihydrofolate reductase (DHFR). It is noteworthy that high TS expression, in particular, has been linked to poor clinical outcome and resistance to antifolates in several cancers including the bladder, lung, mesothelium, and colon [81-85]. Moreover, we showed that loss of ASS1 was correlated with high levels of TS and DHFR and predicted for a poor outcome in patients with transitional carcinoma of the bladder. Subsequently, we validated that arginine deprivation with ADI-PEG20 potentiated the effect of pemetrexed with enhanced apoptosis as assessed by cleaved poly (ADP-ribose) polymerase and annexin V staining. Furthermore, decreased 18F-fluorothymidine (FLT) uptake by PET may be utilized as a predictive imaging biomarker in selecting tumors susceptible to this drug combination (see below).
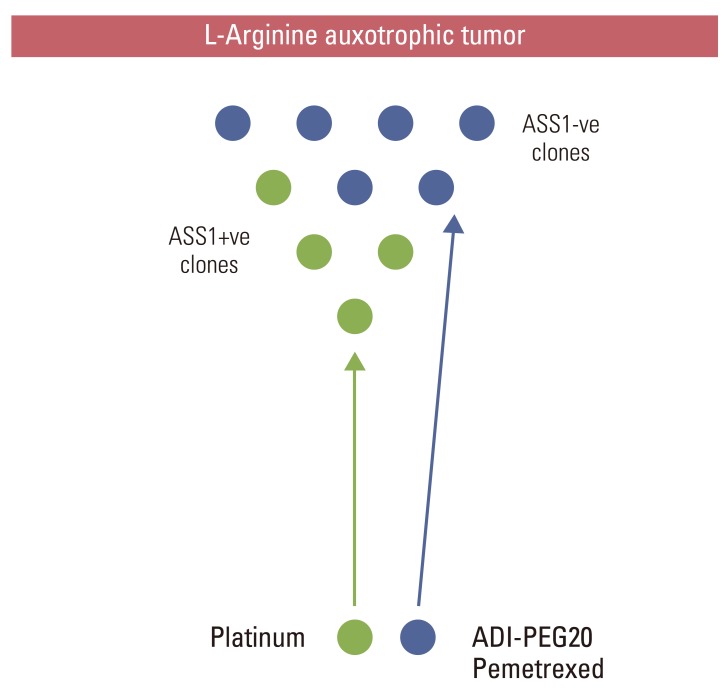
Specific mention should be made here regarding platinum compounds and their relationship to ASS1 and arginine auxotrophy. Helleman et al. [86] first identified loss of ASS1 as one of 9 genes that predicted independently for cisplatin resistance in a microarray retrospective study of patients with ovarian cancer [87]. Subsequently ASS1 gene overexpression and knock-down in ovarian cancer cells confirmed that while ASS1 loss conferred platinum resistance, there was collateral sensitivity to arginine deprivation [88]. In contrast, studies linking ASS1 loss to enhanced platinum salt sensitivity, namely oxaliplatin and human arginase in HCC cell lines, and cisplatin and ADI-PEG20 in melanoma cells, may exploit a prosurvival pathway that depends on NO synthesis derived from exogenous arginine [89,90]. Consistent with this hypothesis, targeted inhibition of inducible NO synthase synergizes with cisplatin chemotherapy in the treatment of arginine-auxotrophic melanoma cells [91,92].
Lastly, there is increasing interest in the development of novel types of human arginase, in particular a pegylated cobalt-modified (i.e., substituted for native manganese) enzyme (HuArg I [Co]-PEG5K) that has shown good efficacy in depleting arginine [93,94]. Animal toxicity testing indicates that citrulline supplementation is required since arginase, unlike ADI, produces ornithine which cannot be recycled readily to citrulline [95]. Several studies confirm that HuArg I [Co]-PEG5K displays tumor cytotoxicity in vitro and is active in xenograft tumor models [96,97].
Arginine Deprivation: Preclinical Data
The initial phase I/II studies of ADI-PEG20 were performed in patients with liver cancer and melanoma, tumors with a high frequency of ASS1 loss (Table 1) [33,98-105]. Following the first intramuscular injection of 160 IU/m2 of ADI-PEG20, plasma arginine levels decrease by 24 hours and remain low for at least seven days. ADI-PEG20 therapy also triggers a reciprocal increase in plasma citrulline and a decline in plasma nitrite and nitrate levels due to reduced NO synthesis. Response rates of between 25-47% were observed in these small studies with good safety and tolerability [98,99]. The common adverse reactions have been self-limiting grade 1-2 injection site reactions, less frequently diffuse skin rashes and arthralgia, and rarely neutropenia, anaphylactoid reactions and serum sickness. The latest ADI-PEG20 clinical trials have recorded stable disease as the best response, with evidence for a rebound in arginine plasma levels due to drug neutralising antibodies that limit the duration of arginine depletion to a median of 50 days [100,101]. However, as pharmacodymanic measurements were made eight days apart it is possible that more frequent blood testing may have detected oscillations in plasma arginine and this needs further study. Notably, Yang et al. [102] performed a randomized phase 2 study of two different doses of ADI-PEG20 (320 IU/m2 vs. 160 IU/m2) in HCC showing that disease control rates were similar in both groups. A trend towards better survival was observed in patients with >4 weeks (10.0 months) compared with <4 weeks (5.8 months) of arginine deprivation. Based on these encouraging results, a large phase 3 trial (ClinicalTrials.gov Identifier: NCT01 287585) is currently accruing in HCC using the 160 IU/m2 dose with results expected in 2015.
The role of ASS1 as a predictor of response to ADI-PEG20 has been explored only recently in clinical trials. Feun et al. [33] assessed the role of ASS1 expression in a trial of ADI-PEG20 in patients with metastatic melanoma. Responses were confined to patients with ASS1-negative tumors, and were seen only with the higher ADI-PEG20 dose of 320 IU/m2. An improvement in the progression-free survival (PFS) was detected in patients with ASS1-negative compared to ASS1-positive tumors (p=0.025) however no overall survival (OS) difference was observed in this small study. In turn, we have conducted the first prospective multicenter randomized study of ADI-PEG20 in patients pre-selected for ASS1 deficiency in cancer by focusing on the asbestos-related cancer mesothelioma (arginine deiminase and mesothelioma or ADAM). ADAM met its primary endpoint of an improvement in the PFS in the best supportive care (BSC) plus ADI-PEG20 group compared to the BSC alone group of almost 6 weeks (p=0.02) [103]. Our data support differential methylation as the basis for the loss of ASS1 observed in mesothelioma, consistent with our earlier cell line studies.
Thus far, human trials of arginase have been confined to a small phase I study in patients with HCC using peg-rhArgI (BCT, Hong Kong, China), a human pegylated enzyme derived from an Escherichia coli expression system [104]. In contrast to ADI-PEG20, peg-rhArgI was given intravenously (I.V.) and reduced plasma arginine to undetectable levels throughout the treatment period with no evidence for neutralizing antibodies. Interestingly, the PFS and OS for the peg-rhArgI are consistent with that observed in trials of ADI-PEG20 in heavily pre-treated patients with HCC and suggest that the ability to switch on tumoral resistance mechanisms may be critical in determining efficacy to arginine depletors, rather than the duration of arginine depletion per se.
Imaging Arginine Depletion in Human Cancer
Several groups have assessed potential imaging biomarkers of tumor response to arginine deprivation. Ott et al. [101] assessed the effect of arginine deprivation with ADI-PEG20 on metastatic melanoma with both early and late imaging timepoints "using PET." Partial metabolic responses (PMRs) were detected at the early time point in ~25% of patients (day 4), but were not sustained by the late time point (56 days), likely due to a combination of early resistance (e.g., re-expression of ASS1 and induction of autophagy) and a rebound in plasma arginine levels. We also performed early time point imaging (day 15) in the ADAM trial, and while by modified Response Evaluation Criteria in Solid Tumors (RECIST) we confirmed stable disease as the best response, almost 50% of patients had evidence of a PMR by imaging with 18F-FDG PET/CT [103,105].
One of the drawbacks in imaging some arginine auxotrophs such as melanoma, is the apparent increase in glucose uptake following ADI-PEG20 therapy secondary to PTEN loss and PI3K-induced glucose transporter type 1 (GLUT-1) expression [47]. Thus, we and others have explored FLT as an alternative imaging PET tracer and although this does not appear predictive in ASS1-negative melanoma, our studies confirmed robust suppression of FLT uptake following ADI-PEG20 treatment of ASS1-methylated tumors [20,106]. Moreover, the uptake of FLT correlated directly with thymidine-kinase 1 (TK1) protein levels consistent with the mode of action of ADI-PEG20 in modulating protein turnover. A trial is planned to explore the role of FLT in patients with ASS1-deficient thoracic cancers as part of a phase I/II study discussed in more detail below.
Future Directions: Combination Therapies
Targeting arginine is moving towards combinatorial studies based on the emerging preclinical data of additive and synergistic drug interactions in the treatment of arginine auxotrophic cancers. Several ADI-PEG20 combination trials are ongoing or planned and follow the developmental template for asparaginase several decades ago (summarized in Table 2). Studies of the doublet of ADI-PEG20 and cisplatin and ADI-PEG20 and docetaxel to date have shown no untoward toxicity other than that ascribed to either agent alone (Bomalaski J, personal communication).
Building on the ADAM trial, we have hypothesized that ADI-PEG20 with the standard doublet of cisplatin and pemetrexed may provide a novel way of targeting the heterogeneity and chemoresistance of mesothelioma. As previously mentioned, several epithelial tumors, including mesothelioma and non-small cell lung cancer, harbor cells with varying degrees of dependence on exogenous arginine as inferred by the differential expression of ASS1. Since ASS1 positivity is a biomarker of cisplatin sensitivity whereas ASS1 loss is a predictor of cisplatin resistance but collateral sensitivity to ADI-PEG20-pemetrexed, the triplet regimen may be superior to standard chemotherapy alone (Fig. 2). Once the maximum tolerated dose has been defined from the phase 1 triplet combination study, early FLT-PET/CT will be assessed as a potential biomarker of response in patients with thoracic malignancy. Patients will be imaged at baseline and at 24hrs following a single dose of ADI-PEG20 with additional FLTPET/CT scans after the first cycle of chemotherapy and upon completion of all treatment.
From the preclinical studies referred to above, the role of combining arginine deprivation with autophagy inhibitors, glutamine and glycolytic inhibitors, modulators of the tumor microenvironment and radiotherapy, are just a few of the exciting questions to explore in the context of future clinical trials.
Conclusions
Arginine deprivation has the potential to improve the management of several hard-to-treat cancers by exploiting differential expression and regulation of key urea cycle enzymes, ASS1 and ASL. Clinical trials that employ biomarkers for patient selection and treatment response are needed to optimize arginine deprivation therapy in cancer. The field is evolving towards combination strategies and it is hoped that arginine-targeted therapies will soon become part of the standard cancer armamentarium.
 XML Download
XML Download