

 PDF
PDF Citation
Citation Print
Print


Introduction
Materials and Methods
1. Subjects
2. DNA isolation and genotyping
3. Statistical analysis
Results
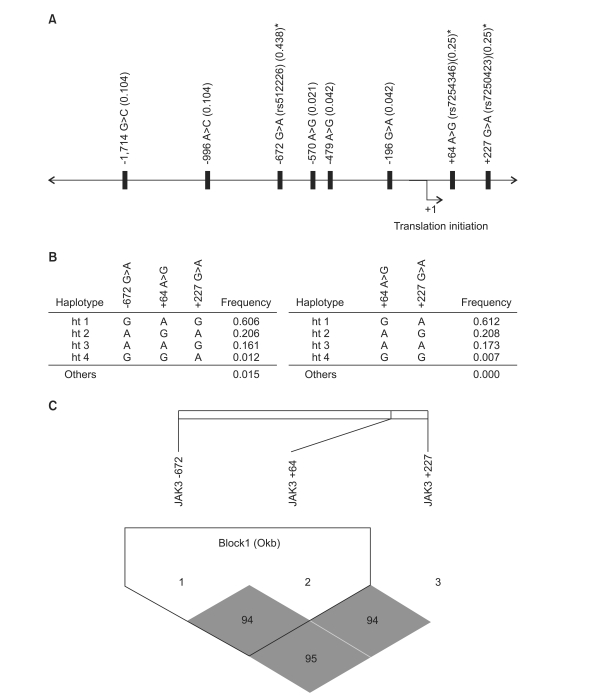 | Fig. 1Map of polymorphisms, haplotypes, and linkage disequilibrium (LD) coefficients in janus tyrosine kinase3 (JAK3) gene. (A) The location of 8 polymorphisms in the JAK3 gene on chromosome 19p13.1. Asterisks indicated polymorphisms that were genotyped in a larger population. The frequencies of polymorphisms were based on sequencing data (n=24). The first base of the translation site was denoted as nucleotide +1. (B) Haplotypes of the promoter region in JAK3 gene. (C) Linkage disequilibrium coefficient (|D'|) among JAK3 gene. Two polymorphisms, including +64 A>G and +227 G>A were used for construction of haplotype. The LD between the polymorphisms was quantified using the Haploview program ver. 3.2.
|
Table 1
![]()
Table 2
![]()
Table 3
| Loci | Genotype | Case | Control |
Dominant aOR (95% CI) |
Recessive aOR (95% CI) |
Codominant aOR (95% CI) |
|---|---|---|---|---|---|---|
| -672 G>A | GG | 148 (36.2) | 158 (38.5) | 1.17 (0.88-1.55) | 0.95 (0.64-1.41) | 1.07 (0.87-1.31) |
| (rs6512226) | GA | 201 (49.1) | 178 (43.4) | |||
| AA | 57 (13.9) | 58 (14.2) | ||||
| +64 A>G | AA | 244 (59.7) | 253 (61.7) | 1.18 (0.89-1.57) | 0.71 (0.37-1.35) | 1.07 (0.84-1.35) |
| (rs7254346) | AG | 146 (39.7) | 120 (29.3) | |||
| GG | 17 (4.2) | 23 (5.6) | ||||
| +227 G>A | GG | 240 (58.7) | 249 (60.7) | 1.17 (0.88-1.55) | 0.70 (0.36-1.36) | 1.07 (0.84-1.35) |
| (rs7250423) | GA | 153 (37.4) | 128 (31.2) | |||
| AA | 16 (3.9) | 22 (5.4) | ||||
| ht G-A-Ga) | -/- | 58 (14.2) | 58 (14.2) | 1.09 (0.73-1.61) | 0.85 (0.64-1.14) | 0.94 (0.77-1.16) |
| +/- | 204 (49.9) | 170 (41.5) | ||||
| +/+ | 143 (35) | 146 (35.6) | ||||
| ht A-G-Aa) | -/- | 248 (60.6) | 251 (61.2) | 1.29 (0.96-1.73) | 0.83 (0.42-1.65) | 1.17 (0.91-1.49) |
| +/- | 141 (34.5) | 105 (25.6) | ||||
| +/+ | 16 (3.9) | 18 (4.4) | ||||
| ht A-A-Ga) | -/- | 285 (69.7) | 267 (65.1) | 1.06 (0.78-1.45) | 0.85 (0.37-1.94) | 1.03 (0.79-1.34) |
| +/- | 109 (26.7) | 95 (23.2) | ||||
| +/+ | 11 (2.7) | 12 (2.9) | ||||
| ht G-Ab) | -/- | 58 (14.2) | 60 (14.6) | 1.11 (0.75-1.64) | 0.85 (0.64-1.13) | 0.95 (0.77-1.16) |
| +/- | 202 (49.4) | 170 (41.5) | ||||
| +/+ | 145 (35.5) | 151 (36.8) | ||||
| ht A-Gb) | -/- | 245 (59.9) | 249 (60.7) | 1.23 (0.92-1.64) | 0.75 (0.38-1.47) | 1.11 (0.87-1.42) |
| +/- | 144 (35.2) | 112 (27.3) | ||||
| +/+ | 16 (3.9) | 20 (4.9) | ||||
| ht A-Ab) | -/- | 278 (68) | 263 (64.2) | 0.13 (0.76-1.39) | 0.81 (0.37-1.77) | 1.00 (0.77-1.29) |
| +/- | 115 (28.1) | 104 (25.4) | ||||
| +/+ | 12 (2.9) | 14 (3.4) |
Values are presented as number (%). Logistic regression models were used to calculate the aORs, 95% CIs and the corresponding p-values of codominant (minor allele homozygotes vs. heterozygotes vs major allele homozygotes), dominant (minor allele homozygotes + heterozygotes vs. major allele homozygotes), and recessive (minor allele homozygotes vs. heterozygotes+major allele homozygotes) models whilst controlling for age and sex as covariates. aORs and 95% CI were calculated by logistic regression and adjusted for age and sex. JAK3, janus tyrosine kinase3; aOR, adjusted odds ratio; CI, confidence interval. a)Haplotype consisting of markers -642 G>A,+64 A>G and +227 G>A, b)Haplotype consisting of markers -642 G>A and +64 A>G.
![]()
Table 4
| Subgroup | Haplotype |
Associated allele |
Case | Control |
Dominant aOR (95% CI) |
p-value | p-valuea) |
Recessive aOR (95% CI) |
p-value | p-valuea) |
Codominant aOR (95% CI) |
p-value | p-valuea) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Male | ht G-A | -/- | 36 (11.4) | 47 (11.4) | 1.52 (0.95-2.44) | 0.08 | NS | 0.87 (0.62-1.22) | 0.42 | NS | 1.04 (0.82-1.32) | 0.74 | NS |
| -/+ | 156 (38.1) | 118 (28.8) | |||||||||||
| +/+ | 110 (26.8) | 107 (26.1) | |||||||||||
| ht A-G | -/- | 181 (44.3) | 181 (44.1) | 1.31 (0.93-1.85) | 0.12 | NS | 0.71 (0.32-1.61) | 0.41 | NS | 1.16 (0.87-1.55) | 0.31 | NS | |
| -/+ | 110 (26.9) | 77 (18.8) | |||||||||||
| +/+ | 11 (2.7) | 14 (3.4) | |||||||||||
| ht A-A | -/- | 216 (52.8) | 183 (44.6) | 0.84 (0.59-1.20) | 0.2 | NS | 0.54 (0.21-1.39) | 0.2 | NS | 0.82 (0.61-1.12) | 0.21 | NS | |
| -/+ | 79 (19.3) | 77 (18.8) | |||||||||||
| +/+ | 7 (1.7) | 12 (2.9) | |||||||||||
| Female | ht G-A | -/- | 22 (5.4) | 13 (3.2) | 0.50 (0.23-1.07) | 0.06 | NS | 0.68 (0.38-1.21) | 0.19 | NS | 0.69 (0.46-1.02) | 0.06 | NS |
| -/+ | 46 (11.2) | 52 (12.7) | |||||||||||
| +/+ | 35 (8.6) | 44 (10.7) | |||||||||||
| ht A-G | -/- | 64 (15.6) | 68 (16.6) | 1.08 (0.61-1.90) | 0.8 | NS | 0.90 (0.26-3.17) | 0.87 | NS | 1.04 (0.65-1.65) | 0.88 | NS | |
| -/+ | 34 (8.3) | 35 (8.5) | |||||||||||
| +/+ | 5 (1.2) | 6 (1.5) | |||||||||||
| ht A-A | -/- | 62 (15.2) | 80 (19.5) | 1.88 (1.04-3.40) | 0.03 | 0.05 | 2.82 (0.52-15.22) | 0.23 | NS | 1.79 (1.07-3.01) | 0.03 | 0.05 | |
| -/+ | 36 (8.8) | 27 (6.6) | |||||||||||
| +/+ | 5 (1.2) | 2 (0.5) | |||||||||||
| Smoker | ht G-A | -/- | 29 (7.1) | 27 (6.6) | 1.48 (0.79-2.77) | 0.22 | NS | 1.01 (0.62-1.65) | 0.98 | NS | 1.12 (0.80-1.58) | 0.5 | NS |
| -/+ | 98 (23.9) | 64 (15.6) | |||||||||||
| +/+ | 65 (15.9) | 47 (11.5) | |||||||||||
| ht A-G | -/- | 113 (27.6) | 89 (21.7) | 1.11 (0.68-1.80) | 0.69 | NS | 0.69 (0.24-1.95) | 0.48 | NS | 1.01 (0.68-1.51) | 0.95 | NS | |
| -/+ | 70 (17.1) | 40 (9.8) | |||||||||||
| +/+ | 9 (2.2) | 9 (2.2) | |||||||||||
| ht A-A | -/- | 131 (32) | 86 (21) | 0.81 (0.49-1.32) | 0.4 | NS | 0.89 (0.25-3.07) | 0.88 | NS | 0.84 (0.55-1.29) | 0.43 | NS | |
| -/+ | 55 (13.4) | 46 (11.2) | |||||||||||
| +/+ | 6 (1.4) | 6 (1.5) | |||||||||||
| Non-smoker | ht G-A | -/- | 21 (5.1) | 8 (1.9) | 0.59 (0.24-1.43) | 0.11 | NS | 0.64 (0.35-1.18) | 0.15 | NS | 0.70 (0.45-1.08) | 0.1 | NS |
| -/+ | 57 (13.9) | 33 (8) | |||||||||||
| +/+ | 39 (9.5) | 31 (7.6) | |||||||||||
| ht A-G | -/- | 73 (17.8) | 44 (10.7) | 0.96 (0.52-1.78) | 0.49 | NS | 1.81 (0.43-9.51) | 0.49 | NS | 1.04 (0.62-1.74) | 0.89 | NS | |
| -/+ | 38 (9.3) | 26 (6.3) | |||||||||||
| +/+ | 6 (1.4) | 2 (0.5) | |||||||||||
| ht A-A | -/- | 75 (18.3) | 57 (13.9) | 2.19 (1.09-4.40) | 0.02 | 0.05 | 3.13 (0.35-27.56) | 0.31 | NS | 2.04 (1.09-3.81) | 0.03 | 0.05 | |
| -/+ | 37 (9) | 14 (3.4) | |||||||||||
| +/+ | 5 (1.2) | 1 (0.2) | |||||||||||
| Drinker | ht G-A | -/- | 38 (9.2) | 28 (6.8) | 1.56 (0.90-2.72) | 0.11 | NS | 0.84 (0.55-1.28) | 0.41 | NS | 1.04 (0.77-1.40) | 0.8 | NS |
| -/+ | 145 (35.4) | 60 (14.6) | |||||||||||
| +/+ | 105 (25.7) | 58 (14.1) | |||||||||||
| ht A-G | -/- | 170 (41.6) | 102 (24.9) | 1.58 (1.02-2.44) | 0.06 | NS | 0.88 (0.32-2.42) | 0.81 | NS | 1.36 (0.94-1.98) | 0.1 | NS | |
| -/+ | 107 (26.2) | 37 (9) | |||||||||||
| +/+ | 11 (2.7) | 7 (1.7) | |||||||||||
| ht A-A | -/- | 207 (50.6) | 93 (22.7) | 0.72 (0.46-1.09) | 0.08 | NS | 0.59 (0.21-1.62) | 0.3 | NS | 0.73 (0.51-1.06) | 0.09 | NS | |
| -/+ | 72 (17.6) | 45 (11) | |||||||||||
| +/+ | 9 (2.2) | 8 (2) | |||||||||||
| Non-drinker | ht G-A | -/- | 20 (4.9) | 19 (4.6) | 0.67 (0.33-1.37) | 0.27 | NS | 0.84 (0.49-1.43) | 0.52 | NS | 0.82 (0.57-1.20) | 0.31 | NS |
| -/+ | 50 (12.2) | 83 (20.2) | |||||||||||
| +/+ | 37 (9) | 60 (14.6) | |||||||||||
| ht A-G | -/- | 69 (16.9) | 96 (23.4) | 0.81 (0.48-1.37) | 0.42 | NS | 0.83 (0.26-2.67) | 0.75 | NS | 0.84 (0.55-1.30) | 0.43 | NS | |
| -/+ | 33 (8) | 57 (13.9) | |||||||||||
| +/+ | 5 (1.2) | 9 (2.2) | |||||||||||
| ht A-A | -/- | 64 (15.6) | 119 (29) | 1.84 (1.07-3.16) | 0.03 | 0.05 | 1.54 (0.25-9.59) | 0.64 | NS | 1.71 (1.04-2.92) | 0.03 | NS | |
| -/+ | 40 (9.8) | 41 (10) | |||||||||||
| +/+ | 3 (0.7) | 2 (0.5) |
Values are presented as number (%). Logistic regression models were used to calculate the aORs, 95% CIs and the corresponding p-values of codominant (minor allele homozygotes vs. heterozygotes vs. major allele homozygotes), dominant (minor allele homozygotes+heterozygotes vs. major allele homozygotes), and recessive (minor allele homozygotes vs. heterozygotes+major allele homozygotes) models whilst controlling for age and sex as covariates. aORs and 95% CI were calculated by logistic regression and adjusted for age and sex. Bold data indicated p-values <0.05. JAK3, janus tyrosine kinase3; aOR, adjusted odds ratio; CI, confidence interval; NS, not significant. a)p-values were calculated for multiple testing using Bonferroni's inequality method.
![]()
 XML Download
XML Download