

 PDF
PDF Citation
Citation Print
Print


Introduction
Alkylation of DNA at the O6 position of guanine is known to be an important step in cancer mutations, because O6-methylguanine tends to pair with thymine during replication, resulting in guanine-cytosine (G : C) to adenine-thymine (A : T) transition mutation. O6-methylguanine-DNA methyltransferase (MGMT) protect cells from this mutation by transferring the alkyl group from the O6-guanine in DNA to an active cysteine within its own sequence (1). Hypermethylation of the promoter region of MGMT gene has been reported to be the most frequent mechanism of MGMT inactivation (2-4). The presence of aberrant hypermethylation has been known to be closely associated with loss of MGMT protein (5). Therefore, it has been suggested that MGMT protein loss via its inactivation mechanism plays an important role in primary human tumorigenesis associated with increased susceptibility to nitrosamines and alkylating agents. A few recent studies on non-small cell lung carcinomas (NSCLCs) have reported that its promoter methylation almost exclusively occurred in tumor cells (6,7). Also, MGMT inactivation through hypermethylation contributes to a large fraction of p53 mutation in NSCLC, including G : C→A : T transition (8,9), which is known to be one of the most important tumor suppressor genes in human.
Correlation between MGMT inactivation and various clinical characteristics has not been studied thoroughly in human malignant tumors including NSCLCs, but controversial data on patient survival (10-12) exists. A study on lung adenocarcinomas suggested that inactivation of MGMT is likely a later event occurring closer to the development of malignancy, based on a significant difference in its prevalence between stages I and II-IV, tumors and tumor sizes (13).
The present study is aimed at determining the prevalence of MGMT expression loss by immunohistochemistry in 112 human NSCLCs specimens and correlating it with various clinicopathologic parameters, including survival data to elucidate the role of MGMT expression in carcinogenesis and progression of NSCLCs.
Materials and Methods
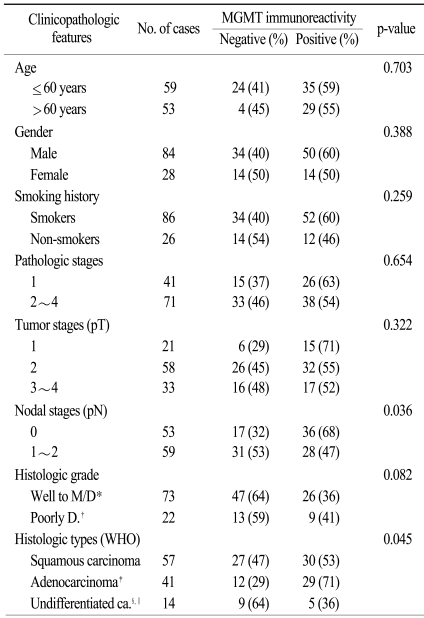
1. Patient characteristics (Table 1)
Tumor samples and clinical follow-up data were obtained from a consecutive collection of 112 stage I-IV NSCLC patients operated at the Dankook University Hospital (DKUH) between 1994 and 2002. The cohort included 84 men (75%) and 28 women (25%). The median age at diagnosis was 60.2 years (range 29~83) and the median survival was 46.1 months (range 15 days~157 months). Of the 112 patients, 86 (76.8%) had a history of smoking. Classifications of pathological staging are as follows: 26 cases of stage I, 16 cases of stage II, 20 cases of stage III and 2 cases of stage IV. The pathological T staging classifications were as follows: 21 cases of T1, 58 cases of T2, 29 cases of T3 and 4 cases of T4. The pathological N stages were as follows: pN0 in 53 cases, pN1 in 28 and pN2 in 31. Death from lung cancer was the terminal event for survival calculations.
2. Tumor samples
Tissue microarray (TMA)s were constructed using pairs of the most representative tumor and normal tissue cores, 2 mm in diameter, from the appropriate areas in formalin-fixed paraffin-embedded tissue blocks. These tissue cores were transferred and embedded into the recipient block that had 60 empty 2 mm-sized holes. Serial sections were cut from the TMA blocks at 4 µm thickness with a microtome and they were transferred to poly-L-lysine-coated slides.
3. Immunohistochemical staining and semiquantitative analysis
The microarrayed tissue sections were deparaffinized with standard xylene and rehydrated through graded alcohols to water. The sections were microwaved in 10 mM citrated buffer at 90℃ for 10 min, then treated with 3% H2O2-PBS solution to reduce endogenous peroxidase activity. They were incubated with normal bovine serum to reduce non-specific antibody binding, and subsequently subjected to primary antibody reactions. The standard avidin-biotin-peroxidase complex method was used for immunohistochemical examination with monoclonal antibodies against MGMT (MT23.2, Novus Biologicals, CO) and p53 (SP5, Thermo Scientific, CA). Each primary antibody was reacted with the sections at room temperature for one hour at 1 : 40,000 and 1 : 100 dilution, respectively. A negative control was incubated without the primary antibodies. Positive controls for MGMT protein were matched samples of normal bronchial mucosa and peripheral lung parenchyma. Detection of immunoreactive staining was carried out by the avidin-biotin-peroxidase complex method with the LSAB kit (DAKO, Glostrup, Denmark). The sections were subjected to a color reaction with 3,3-diaminobenzidine tetrahydrochloride that contained 3% H2O2 in Tris buffer. They were lightly counterstained with Meyer's hematoxylin.
The immunoexpressions for MGMT and p53 proteins were analyzed semiquantitatively, with the qualitative basis of the distinct nuclear staining regardless of the associated cytoplasmic staining, which was faint and considered non-specific. Cases were considered to demonstrate loss of MGMT expression when less than 10% of the tumor cells is immunostained, with the background of normal internal or matched positive controls stained in over 10% of area (12). Cases were interpreted to overexpress p53, when more than 10% of tumor cells showed a distinct nuclear immunoreactivity as well as rare to absent immunopositivity in the normal cells.
4. Statistical analysis
Comparison of each immunoreactivity between two- or three-categorical, clinicopathologic variables and the relationships between MGMT and p53 were analyzed by the chi-square test (SPSS 15.0). The survival period was defined as the time elapsed from the date of surgery to the date of death, or the date of the last follow-up. The survival data of 112 NSCLC patients was analyzed by the Kaplan-Meier method and then compared by the log-rank test. A p-value of less than 0.05 was defined as statistically significant.
Results
1. Histopathological tumor characteristics and MGMT immunoexpression patterns in tumor and normal tissues
The histological subtypes of the 112 tumor specimens were classified according to the WHO criteria (2004): 57 squamous carcinomas (51%), 38 adenocarcinomas (ADCs) (34%) including 3 bronchioloalveolar carcinomas (BACs), 3 adenosquamous carcinomas (ASCs) (2.5%), 7 large cell carcinomas (LCCs) (6.25%), 7 pleomorphic carcinomas (PLCs) (6.25%). For the statistical analyses, the 3 ASCs were grouped into the ADC category, and 7 LCCs and 7 PLCs were combined to form an undifferentiated carcinoma group (Table 1).
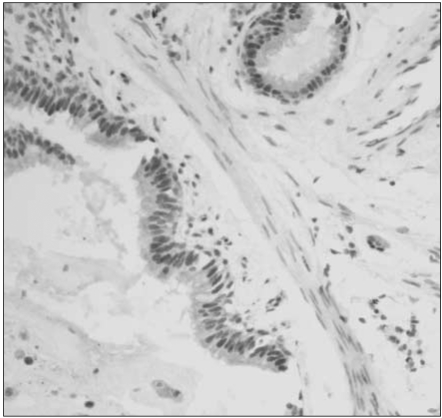
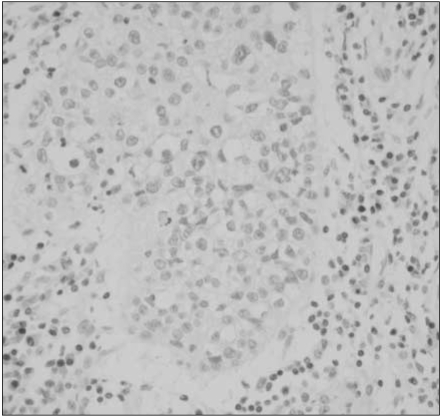
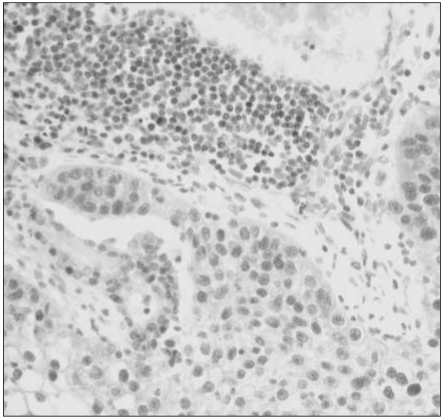
For MGMT protein, normal bronchial epithelia and alveolar pneumocytes were stained distinctly in the nuclei, with the occasional immunoreactivity in inflammatory or stromal cell nuclei (Fig. 1). Overall, 48 of 112 specimens (43%) showed MGMT immunostaining in less than 10% of the tumor cells, which suggests significant loss of MGMT expression in NSCLC. Their nuclear immunoreactivity for MGMT protein was found to be markedly lost in the tumor cells, in contrast to the normal stromal cells with retained MGMT-immunoreactivity (Fig. 2). However, a considerable portion (57%) of 112 specimens exhibited retained MGMT expression similar to that of the normal pulmonary tissue components (Fig. 3).
2. Correlation of MGMT expression with clinicopathologic prognostic factors (Table 1)
The loss of MGMT expression was significantly associated with nodal metastasis because it was found to be more frequent in patients with pN1-2 (53%) disease, than in patients with pN0 (32%) disease (p=0.036). MGMT expression was lost more frequently in histologic subtypes such as undifferentiated and squamous carcinomas than in adenocarcinomas (p=0.045). It also tended to be significantly higher in the histologic grade 1-2 than in the grade 3 (p=0.082) diseases. However, age, sex, smoking history, pathologic stage and tumor size were not correlated with MGMT expression loss.
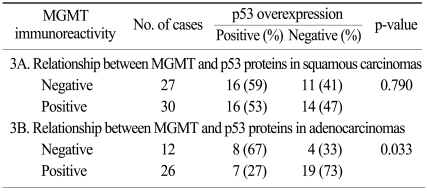
3. Association of MGMT expression with p53 overexpression (Table 2 and Table 3)
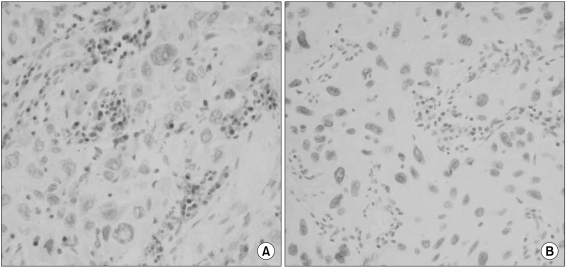
Overall, p53 overexpression was found in 55 (49.1%) of the 112 specimens. As for correlation with MGMT expression, 48 specimens with loss of MGMT expression exhibited p53 overexpression in 56% (Fig. 4), whereas 64 cases with retained MGMT proteins demonstrated 44% of p53 overexpression. The difference in p53 overexpression between them was not statistically significant in overall NSCLCs (p=0.252). However, adenocarcinomas with loss of MGMT expression showed significantly higher proportion of p53 overexpression than those with retained MGMT expression (p=0.033). Squamous carcinomas didn't reveal any significant correlation between loss of MGMT expression and p53 overexpression (p=0.790).
4. Effect of MGMT loss on patient survival (Data not shown)
Kaplan-Meier survival estimates in 112 cases did not reveal statistically significant difference in patient survival between MGMT-negative and -positive cases throughout the follow-up period (range: 15 days~157 months) (p=1.189). In addition, when survival data was analyzed by combining MGMT and p53 immunostainings into 4 groups, such as MGMT-/p53+, MGMT+/p53-, MGMT+/p53+, and MGMT-/p53-, there was no significant difference between them (p=0.63).
Discussion
MGMT is a DNA repair gene critical for the removal of mutagenic adducts from the O6 position of guanine. MGMT protein expression is lost frequently in tight association with hypermethylation of the promoter region (2,8,13) and aberrant DNA methylation is one of the most consistent epigenetic changes in human cancers (14-16). Therefore, loss of MGMT expression through MGMT inactivation by its promoter methylation has been suggested to play a key role in primary human carcinogenesis, including non-small cell lung carcinomas (5).
In the present study, 48 out of 112 NSCLCs cases exhibited low or absent MGMT expression (43%), suggesting that loss of MGMT proteins might be important in pulmonary carcinogenesis. To the present, altered or reduced MGMT expression has been reported to be up to 78% in a recent study (11). The much higher percentage of absent MGMT expression compared to ours is assumed to be caused by differences in patient populations and scoring system or criteria for the evaluation of immunostaining. The association betweenMGMT and clinicopathologic prognostic factors is still controversial. In one study, MGMT promoter hypermethylation was significantly more common in adenocarcinomas and poorly differentiated tumors (8). However, the present study showed that loss of MGMT expression was significantly less common in adenocarcinomas than in squamous or undifferentiated carcinomas. Adenocarcinomas had a significantly lower MGMT expression (29%) than the other histologic types, such as squamous or undifferentiated carcinomas (47% and 64%, respectively) (p=0.045). These results suggest that MGMT loss is less involved during tumorigenesis of adenocarcinomas than other histologic types. Tumors with nodal metastasis (N1-2) in the present study showed more frequent loss of MGMT expression than those without nodal metastasis (N0). The association of MGMT loss with nodal metastasis has also been reported in other human cancers, such as oral, bile duct, and gastric cancers (17,18). Previous reports underscored its potential as a candidate of predictive marker for nodal metastasis. The present study also suggests that MGMT loss may play an important role in the nodal progression of NSCLC. Most studies including ours have reported that there is no significant difference between tumor stages (8,10,11) and MGMT loss. The prevalence of MGMT methylation was similar in stage I and III tumors in these studies, suggesting that MGMT loss precedes the development of invasive cancer, although another study reported a significant difference in the prevalence of MGMT inactivation between stages I and II-IV tumors (13), indicating that MGMT inactivation is likely a later event which occurs in the development of malignancy.
The data on the patient survival reported to date has also been controversial. A few studies including ours showed no significant relationship between overall patient survival and MGMT expression (11,13), whereas a recent literature reported that MGMT promoter hypermethylation is associated with inferior survival, suggesting that it might be an important biomarker for biologically aggressive disease in NSCLC (10). Our survival data were also analyzed by combined the results of MGMT loss and p53 overexpression, but there was no significant difference between the combined groups (MGMT-/p53+, MGMT+/p53-, MGMT+/+, MGMT-/-) (data not shown). This result suggests that MTMT loss and p53 overexpression do not interact on the patient survival. In the present study, the seemingly contradictory results on patient survival and lymph node metastasis between MGMT loss and its retained expression could be explained by the fact that the patient survival is generally determined by a combination of several clinicopathologic factors including lymph node metastasis.
According to recent literatures, MGMT inactivation by hypermethylation has been reported to be associated with a shift from the G : C→A : T mutation in the p53 gene (9,19), suggesting that MGMT promoter methylation may increase the occurrence of p53 mutation in lung cancer. When the present study investigated the association between MGMT inactivation and p53 overexpression based on the previous observations above, there was no significant relationship between them (p=0.252). These discrepancies appear to be originated from either a difference of the detecting targets, i.e. gene or protein, or inconsistent interpretation of p53 overexpression compared to the relatively consistent results on p53 mutation. However, there was a significant association between reduced MGMT expression and p53 overexpression in adenocarcinomas, whereas squamous carcinomas did not demonstrate similar outcomes in the present study. According to a published literature (8), G to A mutations occur more frequently in lung adenocarcinomas than other types of p53 mutation and they are the consequence of MGMT inactivation. G to A transitions are also generally known to occur in 18~24% of p53 mutations. Therefore, significant association between the loss of MGMT expression and p53 overexpression in adenocarcinomas in the present study could be explained partly by the existing data. In addition, the present results suggest that MGMT loss associated with p53 overexpression plays a role in the development of NSCLCs, especially adenocarcinomas.
Conclusion
Loss of MGMT expression was a relatively common event in human NSCLCs and significantly associated with nodal metastasis and, especially in adenocarcinoma, p53 overexpression. The outcomes of this study suggest that loss of MGMT expression in association with p53 overexpression may be important in pulmonary carcinogenesis and in the disease progression of NSCLCs.

 XML Download
XML Download