

 PDF
PDF Citation
Citation Print
Print


Neurologic Disease and Cancer: The Paraneoplastic Neurologic Degenerations
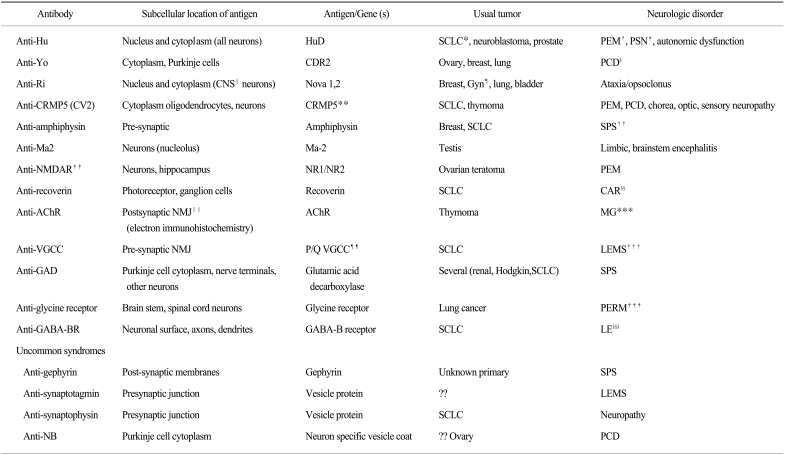
The paraneoplastic neurologic degenerations (PNDs) are a set of brain disorders that develop in the setting of occult cancer (Table 1). These disorders arise when common cancers express brain proteins, triggering an anti-tumor immune response and, at least in some cases, clinically effective suppression of the malignancy. PNDs come to clinical attention only when the tumor immune response breaches immune privilege of the nervous system, and begins to attack the neurons that are normally expressing the tumor antigen (which we have termed onconeural antigens). Our laboratory first demonstrated that genes encoding onconeural antigens can be cloned using high titer antisera from these patients (1-5), and currently over a dozen well-described PND antigens have been defined.
cDNAs encoding PND antigens have provided a wealth of information regarding the nature of the PND antigens (6-8) as well as their role in disease pathogenesis (9-11). For example, the initial cloning of a cerebellar antigen from a woman with presumed ovarian cancer led to the discovery of a long-suspected neuronspecific vesicle coat protein (4). Cloning of the cerebellar antigen (cdr2) associated more commonly with gynecologic cancer revealed the presence of antigen-specific T cells in these patients, establishing a cellmediated component to the disorders, as well as a means by which tumor antigen is likely cross-presented from apoptotic cells to dendritic cells to initiate the anti-tumor immune response (12,13). These studies have led to clinical cancer vaccine trials in prostate cancer patients that have shown some evidence of efficacy in Phase I safety studies (14). Moreover, cloning of the genes encoding T cell receptors from these patients offers a possible new therapeutic strategy, by passive transfer of these genes into T cells to induce tumor-cell targeting (15).
RNA Binding Proteins in Neurologic Disease and Cancer
Among the set of cloned PND antigens, perhaps none has generated as much biologic interest as the Nova and Elavl (Hu) family of neuronspecific RNA binding proteins. The Nova proteins (16,17) are ectopically expressed in lung or gynecologic cancers, and trigger neurologic symptoms (excess motor movements of the eyes and musculature) interpreted by neurologists as failure of inhibitory motor control. Studies of the Nova proteins have established what is arguably the best data available to understand a tissue-specific RNA regulatory protein in the brain; these findings will be discussed here. The Elavl proteins (originally termed the Hu proteins; (18,19) are still incompletely understood (6,20), but are believed to relate to post-transcriptional regulation of RNA in the brain. Many of the approaches used to understand the biology of Nova can be applied more generally-to the understanding of the biology of the Elavl proteins and other RNA binding proteins in the brain and cancer.
Back to the Basics - RBP Functional Studies
Understanding RBP function requires understanding the protein's interaction with RNA. Nova has emerged as one of the best understood mammalian RNA binding proteins in part because there is a good understanding of its RNA substrates. After recognizing that Nova harbors three KH-type RNA binding proteins (21), in vitro RNA selection experiments (22,23) were applied to reveal (17,24) that Nova recognizes a core 4 nucleotide repeat sequence-(UCAU)3. Mutagenesis and X-Ray crystallographic studies identified the CA dinucleotide as a critical invariant component of Nova-RNA binding (25,26), with specificity restricted to bounding pyrimidines, hence the more general consensus of YCAY repeats as the core binding motif.
Genetic Systems and RBP Function
These studies prompted a search for brain transcripts harboring such repeat elements. This relatively pedestrian approach yielded three targets, which were characterized by mutagenesis, boundary mapping, and functional analyses in some detail. The first target transcript harboring a Nova-regulated YCAY cluster was the inhibitory glycine receptor α2 (GlyRα2). Interestingly, this cluster was located within an intronic sequence upstream of an alternatively spliced exon (E3A) of GlyRα2. Analysis of alternative splicing of minigenes transfected into tissue culture cells demonstrated that Nova acted specifically on the YCAY cluster to mediate an increase in inclusion of E3A (24). Subsequently, analogous studies demonstrated that Nova was able to bind intronic YCAY elements to mediate an increase in the γ2L exon of the GABAA transcript (27), and to bind exonic and intronic YCAY elements to autoinhibit splicing of Nova1 exon 4 (28). This led to the hypothesis that Nova might regulate alternative splicing of these and other transcripts in neurons.
This hypothesis was tested in vivo by generating Nova null mice. GlyRγ2 E3A and GABAA γ2L splicing showed consistent two and three-fold decreases in utilization, respectively, in Nova1 KO mice (25), while Nova E4 exclusion was decreased -0.75-fold in Nova1 heterozygous mice (28). These studies established Nova as the first bona-fide mammalian tissue specific splicing factor whose actions were validated in live animals, and set the stage for more global studies of Nova function. These were further stimulated by recognizing the correlation between Nova targeting in PND patients, who show defects in inhibitory motor control, and the observation that 2/2 Nova-regulated transcripts encoded inhibitory neurotransmitter receptors (29).
Bioinformatics, Genetics and Biochemistry: Beginnings of a Holistic Approach to RBP Function
Initial studies aimed at discovering the general nature of Nova action on alternative splicing were done using a new (at the time) Affymetrix exon junction array. Applying WT or Nova KO RNA to these arrays, whose probesets were designed to detect alternative splice junctions, revealed a wealth of new target transcripts that could be independently validated as Nova targets in vivo. Most surprisingly, the results revealed that the vast majority of Nova-regulated transcripts encoded synaptic proteins. More precisely, the steady-state level of these regulated RNAs was unaltered in Nova null mouse brain, but their quality manifest by different ratios of alternative exons-was altered (30).
A limitation of this work came from careful parsing of the term Nova-regulated transcripts. These transcripts showed Nova-dependent changes in alternative splicing, but such changes might be a direct action of Nova-for example, acting on YCAY elements, or an indirect action-for example through regulation of a different splicing factor. Hence two series of studies were used to follow-up the finding from the "correlative" Affymetrix exon junction array. A heuristic bioinformatic algorithm was developed to search for YCAY elements within this set of Nova targets, revealing an enrichment of elements within them (although the algorithm was limited, with -50% false positive rate, in its ability to predict targets de novo) (31).
Surprisingly, a position-dependent action of Nova on these transcripts was also discovered, such that Nova binding elements upstream or within alternative exons correlated with inhibition of exon inclusion, while Nova binding downstream correlated with enhancement of exon inclusion (31). An effort to further strengthen these correlations was made, both by undertaking detailed in vitro splicing assays to provide a mechanistic basis for the positional effects, and analysis of splicing intermediates by RT-PCR, providing evidence for asymmetric actions on splicing intermediates that correlated with the position of the YCAY elements (31).
HITS-CLIP and the Development of a Comprehensive Approach to RBP Function
The heavy use of the word "correlation" in the previous section emphasis what was missing from the microarray and bioinformatic studies-evidence of direct Nova-RNA interaction. Although such evidence was possible on a case-by-case basis, made most persuasively with Nova's action on the GABAA transcript (27,31), it was not possible to undertake such studies, nor develop an entirely compelling description of Nova action, on what begain to approach 100 putative Nova-regulated transcripts. Hence a new approach was needed, and developed in the form of cross-linkimmunoprecipitation (CLIP) (32). CLIP takes advantage of the finding that UV-B irradiation induces covalent complexes between protein-nucleic acids when contact distances are within -1 Å (33). By applying UV-irradiation to acutely dissected mouse brains, Nova-RNA complexes in situ were stabilized, allowing rigorous purification. After purification, removal of the protein with proteinase K, CLIP also established that RNA could be efficiently sequenced, using RNA linker ligation and reverse transcription-PCR. Analysis of 34 transcripts identified independently among the first 340 CLIP targets sequenced revealed that they were enriched in YCAY elements, that 71% encoded neuronal synaptic proteins, and that they included Nova-regulated alternative exons (32). Hence CLIP was established as a genome-wide unbiased means of identifying functional RNA-protein interaction sites.
To broaden the observations made with CLIP, high throughput sequencing (HITS) methods were applied, termed HITS-CLIP, to the study of Nova-RNA interactions in 2008 (34). In these experiments, millions of Novacrosslinked RNA fragments of -50-100 nt were sequenced, and very conservatively winnowed into a pool of 168,632 unique tags mapping across the mouse genome. These validated predicted Nova-regulated alternative exons that had been correlated with Nova by exon junction array and bioinformatic strategies. They also identified new clusters of Nova tags surrounding alternative exons that were in fact predictive of de novo regulated exons, and they confirmed the position-dependent bioinformatic map. Moreover, these studies also identified many previously unknown Nova binding sites. For example, clusters of Nova-RNA tags within 3' UTRs were found in some cases to surround alternative poly (A) sites, and this led to biochemical validation and the discovery that Nova was able to regulate alternative polyadenylation choice in the brain.
These studies were thus able to close the loop on a new approach to genomewide, unbiased discovery of RBP-RNA function. Application of basic biochemical principles to identify high affinity targets, descriptive analysis of RNA variants under conditions of a defined variable, such as a genetic null, and HITS-CLIP to discriminate direct from indirect target transcripts, offers a powerful approach to discovery (35).
Future directions for this approach are several. The wealth of data coming from HITS-CLIP suggests that new bioinformatic approaches will be important in synthesizing information. Indeed, recent studies have demonstrated the use of Bayesian networks to combine data from such datasets as exon junction arrays, HITS-CLIP, and motif analysis to both validate and produce new discovery regarding RBP-RNA interaction (36). The approaches worked out for Nova provide general solutions. A number of other RBPs have now been studied by HITS-CLIP (33,37-41), in efforts that, when combined, may provide the groundwork for development of a true combinatorial map of RNA regulation. Finally, HITS-CLIP analysis has been extended to the study of ternary interactions-between small non-coding RNAs, mRNA and regulatory proteins. This was first established for the mammalian Ago protein, demonstrating that Ago-mRNA crosslinked tags cluster directly over microRNA binding seeds, such that these ternary HITS-CLIP maps could decode sites of miRNA-mRNA interactions on a robust, genome-wide basis (42). Subsequently these studies have been reproduced in C elegans (43) and mammalian tissue culture cells (36), and have begun with other small RNAs (piRNAs) (44). These studies offer the exciting possibilities of both developing better understandings and means of capitalizing upon small RNA regulation of gene expression, and overlaying such maps with those developed with HITS-CLIP analysis of more traditional RBPRNA interactions.
 XML Download
XML Download