

 PDF
PDF Citation
Citation Print
Print


Introduction
Gastric cancer is one of the most common cancers and a leading cause of cancer-related death around the world. In high-risk areas such as Korea, Japan and China, the age-standardized incidence rate of gastric cancer is greater than 20 per 100,000, even though its incidence has gradually decreased (1). Gastric carcinogenesis is a multistep process, with interactions between both host factors and environmental factors, and the accumulation of numerous genetic and epigenetic alterations. Abnormalities occur, for example, in growth factors/receptors, angiogenic factors, cell cycle regulators and DNA mismatch repair genes (2).
Osteopontin (OPN) was first identified as a non-collagenous bone matrix protein, and it was subsequently shown to be a cytokine that regulates cell trafficking in the immune system (3). In addition, OPN signaling represents a key regulatory circuit in tumorigenesis, tumor invasion and metastasis in breast, lung, prostate, ovary and colon cancers (4,5). OPN can bind to various cell surface receptors, including the vitronectin receptor (ανβ3 integrin) and CD44 (6), and OPN can induce changes in the gene expression of tumor cells, including the induction of proteolytic enzymes and the activation of growth factor kinases, which may lead to increased cell motility and invasion (7). CD44 is a cell surface glycoprotein that can be expressed as a standard receptor (CD44s) and as multiple splice isoforms (CD44v), and the expressions of these can be altered during tumor growth and progression. CD44 binds hyaluronate in the extracellular matrix and it increases the metastatic potential of tumor cells by promoting migration (8).
Nuclear factor κB (NFκB) is a family of transcription factors that are involved in inflammation, the immune response, cell proliferation and apoptosis (9). Activation of NFκB has been detected in lymphomas, melanomas and breast cancers. NFκB proteins induce anti-apoptotic genes that protect cancer cells from apoptosis, and NFκB proteins regulate the expression and activation of matrix metalloproteinases, which play major roles in extracellular matrix degradation and they facilitate cell motility, tumor growth and metastasis (10). NFκB is known to be involved in the rapid onset of OPN transcription (11) and OPN induces the NFκB-mediated signaling pathways by binding to its receptor.
OPN binds to the cell surface receptor CD44 and it also activates the NFκB pathway. The NFκB site is known to be located in the OPN promoter site. CD44 combined with hyaluronan promotes the activation of the NFκB pathway (12). Yet the precise interactive roles of these molecules are not well defined, and the research on their expression in gastric carcinoma is quite limited. This study was conducted to evaluate the expressions of OPN, CD44 and NFκB in gastric adenocarcinoma and to explore their relationships with various clinicopathological factors.
Go to : 
Materials and Methods
1. The patients and tumor samples
We studied 211 patients with primary human gastric carcinoma and who were treated between 1992 and 1998 at Sanggye Paik Hospital in Seoul, Korea. The ethical use of the human tissue for research was approved by our Institutional Review Board. Of the patients, 138 were men and 73 were women. Their mean age was 57.3±12.4 years (range: 28~82). All the tumors were adenocarcinomas by the histologic type, including eight cases of mucinous carcinoma and 21 cases of signet ring cell carcinoma. Most of the tumors were moderately differentiated (118 patients) and the body and antrum were the most frequently involved sites. The ulceroinfiltrative type was most frequent (96 cases). The pathologic T stage was primarily pT3 (87 patients), with 152 cases of advanced gastric cancer (AGC) and 59 cases of early gastric cancer (EGC). Early gastric cancer was defined as a lesion confined to the mucosa and submucosa, regardless of the presence of perigastric lymph node metastases. None of the patients had undergone preoperative chemotherapy or radiotherapy. The median follow-up period was 137.0 months (range: 41.6~172.1). The tumor specimens were fixed in 10% buffered formalin; they were routinely processed and then embedded in paraffin wax.
2. Tissue microarray block construction and the immunohistochemistry
We constructed the tissue microarray blocks as previously described (13). Sections (4µm) of the microarray blocks were cut and immunohistochemical staining was performed. The following antibodies were used: anti-OPN monoclonal antibody (NovoCastra, UK), anti-CD44 (Zymed, San Francisco, CA), anti-CD44v6 (Lab Vision, Fremont, CA) and NFκB/p65 (Rel A) (Lab Vision). For each antibody, the deparaffinized tissue sections were placed in 10mM citrate buffer (pH 6.0) and heated at 100℃ for 20 min for antigen retrieval. After incubation with each antibody at room temperature for 1 h, the slides were subsequently incubated with a biotinylated secondary horse anti-mouse IgG antibody for 30 min and the reaction was detected with avidin-conjugated horseradish peroxidase (DAKO, Carpenteria, CA). The color was then developed using 3,3'diaminobenzidine (DAB; ScyTek, Logan, UT). Mayer's hematoxylin was then used as a counterstain. Gallbladder and tonsil were used as positive controls, and staining without the primary antibodies was used as a negative control. We performed the staining in two sets of microarray blocks and any discrepancy of the immunostaining results was evaluated by staining with using the conventional paraffin tumor sections.
Granular cytoplasmic staining was considered positive for OPN. Though NFκB was located both in the cytoplasm and in the nucleus according to the activation state, we used the commercial antibody and the cytoplasmic staining was considered positive for NFκB as per the instructions. The expression levels were evaluated according to the proportion and intensity of the granulation. The intensity of the staining was scored using the following scale: 0, no staining of the tumor cells; 1+, mild staining: 2+, moderate staining; 3+, marked staining. The area of staining was evaluated and recorded as a percentage: 0, less than 10%; 1+, 10% to 25%; 2+, 25% to 50%; 3+, 51% to 75%; 4+, more than 75%. The combined score was recorded, and a score of more than 3 was regarded as positive. The presence of more than 10% of distinct cell membrane staining was considered positive for CD44 and CD44v6.
3. mRNA quantification using real-time reverse transcription polymerase chain reaction (RT-PCR)
Tumor and the normal mucosal tissue remote from the tumor were collected and frozen in liquid nitrogen. We selected 10 cases with relatively well preserved samples. The total RNA was isolated from the fresh frozen tissue using the TRIzol Reagent (Canadian Life Technologies, Burlington, Canada) according to the manufacturer's protocol. After purification, the RNA concentration was determined by the absorbance at 260 nm by routine spectrophotometric analysis. Reverse transcription of the RNA was performed with an RNA PCR Kit (TaKaRa, Japan) in a final volume of 10µL containing 1µL of 10× reverse transcription buffer (250 mM Tris-HCl, 40 mM MgCl2, 150 mM KCl, 5 mM dithiothreitol, pH 8.5), 1 mM of each deoxynucleotide triphosphate, 1 U/µL of RNase inhibitors, 2.5 µM random 9-mers, 0.25 U/µL of reverse transcriptase and 1µL of total RNA. The samples were incubated at 30℃ for 10 min and then at 42℃ for 30 min, and the reverse transcriptase was inactivated by heating the samples at 99℃ for 5 min.
Real-time quantitative PCR was performed using the SYBR Green PCR MasterMix (Applied Biosystems, Foster City, CA) in an iCycler IQ Multi-Color Real Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA). The amplification reactions (20µL final reaction volume) contained 200 nM of each primer, 3 mM MgCl2, 300µL deoxy-NTPs, 2× SYBR Green PCR Buffer (SYBR Green PCR Master Mix; Qiagen, Valencia, CA), 0.1 U/µL AmpliTaq Gold DNA polymerase, 0.01 U/µL Amp Erase, ribonuclease-free water and 1µL of the cDNA samples. The amplification was monitored by measuring the increase in fluorescence caused by SYBR Green binding to double-stranded DNA. The fluorescence threshold values were measured, and the fold changes were calculated by the formula where ΔCt is the difference between the amplification fluorescence thresholds of the mRNA of interest and GAPDH (14). The primer sequences were as follows: OPN forward, 5'-AGC GAG GAG TTG AAT GGT G-3'; OPN reverse, 5'-GAC TGC TTG TGG CTG TGG-3'; GAPDH forward, 5'-ACA TCA TCC CTG CCT CTA CTG-3'; and GAPDH reverse, 5'-CCC AGG ATG CCC TTG AGG-3'. Thermocycling was carried out for 40 cycles, with denaturation at 95℃ for 20 s, annealing at 60℃ for 30 s and extension at 72℃ for 45 s.
4. Statistical analysis
The statistical associations between the protein expression levels of OPN, CD44 and NFκB and the clinicopathological factors were determined using chi-square tests, Fisher's exact tests and Pearson's tests. Overall survival was calculated by the Kaplan-Meier method, and the differences between the survival rates were compared using the log-rank test. Cox's proportional hazards model was used to identify the prognostic factors that influenced survival. P values <0.05 were deemed statistically significant. All the statistical analyses were performed using SAS software (SAS Institute Inc., Cary, NC)
Go to :
Results
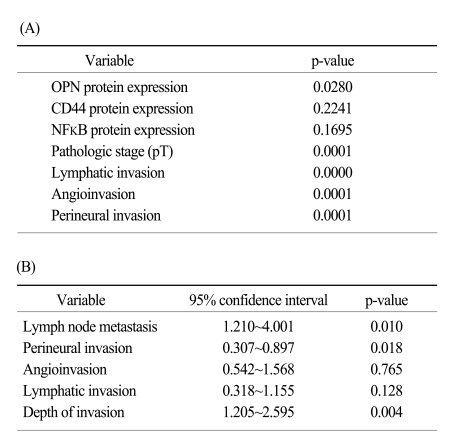
Between 205 and 210 of the tumor samples could be interpreted after immunohistochemical staining; the failed analyses were due to missing cores or too few tumor cells in the cores. The protein expression of OPN, CD44 and NFκB was noted in 61.7% (129/209 cases), 11.4% (24/210 cases) and 26.6% (55/207 cases), respectively, of the adenocarcinoma tissue samples (Table 1, Fig. 1). A OPN protein expression was negatively correlated with angioinvasion (p<0.05), but it was not correlated with any of the examined clinicopathological factors, including age, gender, location, stage, lymph node metastasis, lymphatic invasion or perineural invasion. The CD44 and NFκB expressions were not associated with any clinicopathological factors. No difference in protein expression was observed according to the gross type, the microscopic subtype or differentiation. The OPN, CD44 and NFκB protein expressions were not correlated with each other (Table 2), and a CD44v6 protein expression was not observed in any case (0/171 cases). On comparing the EGC and AGC, no differences of the OPN, CD44 and NFκB expressions were noted (Table 3). Real-time RT-PCR for OPN was performed in 10 cases, and the expression of OPN mRNA was found to be slightly higher in the adenocarcinoma samples than in the paired normal mucosa (23.09±30.41 versus 16.18±13.41, respectively); however, this difference was not statistically significant.
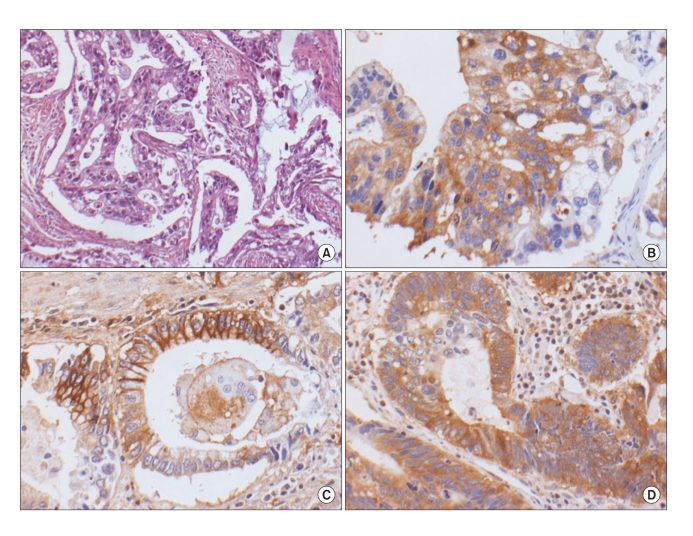 | Fig. 1The expression of OPN, CD44 and NFκB proteins in gastric adenocarcinoma tissue [A, H&E (×100); B, OPN (×200); C, CD44 (×200); D, NFκB (×200)].
|
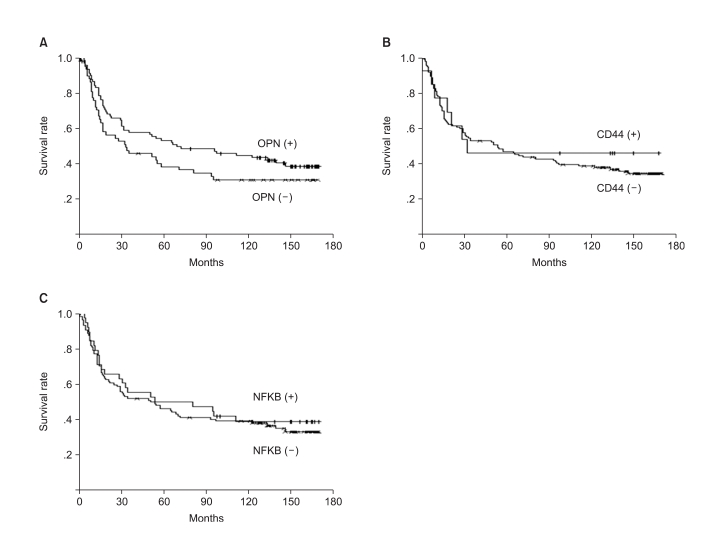
The OPN protein expression was negatively correlated with patient survival, but the CD44 and NFκB expressions were not correlated with survival (Fig. 2). Tumor size, depth of invasion, lymphatic invasion, angioinvasion and perineural invasion were the prognostic factors on the log-ranks test, but on Cox's regression analysis, only the depth of invasion, the lymph node status and perineural invasion were independent prognostic factors (Table 4).
Go to :
Discussion
OPN was recently suggested to be a marker of cancer progression (15). The level of the OPN expression is known to be positively correlated with the tumor stage, metastasis and patient survival (16). Patients with elevated plasma OPN levels have significantly poorer prognoses, and the plasma OPN level has high sensitivity to detect the development of cancer (17). Another study revealed higher OPN immunoreactivity in gastric carcinoma, but there was no significant correlation with the stage of gastric carcinoma (18). CNS tumor cells expressing a high level of OPN failed to generate aggressive tumor in vivo, resulting in significantly prolonged survival and in some animals, no sign of tumor formation was detected (19). Researchers have reported a low or negative OPN expression in breast, skin and salivary gland tumors, and there was a significant negative correlation of stage with the OPN expression in mouth cancer (18). We found slightly increased OPN protein and mRNA expressions in gastric adenocarcinomas, but the group with a positive OPN expression had a little longer survival. There have been several hypotheses proposed as to why no significant difference of the OPN expression was detected between survival and OPN expression scores. One was the different expression and value of OPN according to the primary site of the cancer (20). The second was the OPN gene expression was elevated in the majority of ductal carcinomas in situ in the breast and in the invasive front of oral squamous cell carcinoma, which supports OPN having a role in the early stages of cancer progression and it being an early invasion marker rather than a prognostic marker (7). The third was the concentration-dependent nature of OPN's effect on migration. Once a threshold expression was exceeded, OPN negatively impacted the malignant character of tumors. Our data was limited to an immunohistochemical study and a small number of real-time PCR samples; as a result, we could not determine the exact role of OPN, but we suggest that the molecular pathways of tumor progression in gastric adenocarcinoma may be different from those associated with OPN overexpression (18).
OPN-CD44 interaction can support survival functions to protect cells that are under environment stress, and this protection may support the transformation process during the initial tumor formation and expansion, while also facilitating metastatic dissemination (21). The studies focused on the OPN expression in colorectal cancer have shown differences; in one study, OPN enhanced migration through binding to CD44 (5), while in another study, the suppression of OPN was significantly correlated with metastasis (22). NFκB is widely known to be involved in cellular proliferation and cancer invasion and it is a target for cancer therapy. The cells transfected with breast cancer metastasis suppressor 1 (BRMS1) expressed significantly less OPN, and this was due to either the reduction of p65 trans-activation or the decreased binding of NFκB to the OPN/NFκB site (12). Hyaluronan binding to CD44 up-regulates the p300 expression and the acetyltransferase activity, and these promote acetylation of beta-catenin and the transcriptional up-regulation of NFκB (23). Despite the close relationship among OPN, CD44 and NFκB, we couldn't find a significant correlation between them. During the cell cycle, the different roles and the degrees of participation of OPN, CD44 and NFκB in gastric carcinogenesis and tumor invasion may be one of the reasons for the lack of obvious correlation between these protein factors. Other interactions between these factors and other membrane receptors may occur during cellular transformation and tumorigenesis (24).
Go to :
Conclusion
The protein expressions of OPN, CD44 and NFκB were noted in the gastric adenocarcinoma tissue. OPN may be involved in gastric carcinogenesis, but the role of CD44 and NFκB may be minor in gastric adenocarcinoma. Further studies that will examine genetic changes will be needed to confirm the relationships among OPN, CD44 and NFκB in human gastric adenocarcinoma.
Go to :
 XML Download
XML Download