

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Wnt glycoproteins regulate cellular homeostasis and development by binding to membrane Frizzled-LRP5/6 receptor complexes (1). Wnt signaling includes a canonical pathway and noncanonical pathways. Canonical signaling causes accumulation of cytosolic β-catenin followed by translocation to the nucleus, where β-catenin acts as a co-activator of T-cell factor (TCF) proteins to regulate gene expression (1,2). Noncanonical pathways lead to the activation of Rho, Rac, JNK and PKC, or to changes in the levels of Ca2+ (1,2).
Dickkopf-1 (DKK-1) is a member of the DKK protein family that includes DKK-1, DKK-2, DKK-3 and DKK-4 (3). DKK-1 encodes a secreted Wnt antagonist that binds to LRP5/6 and so induces its endocytosis, leading to the inhibition of the canonical pathway (1). DKK-1 itself is a target of the beta-catenin/TCF signaling pathway (2,4).
The DKK-1 expression is downregulated in human colon cancer and it is transcriptionally silenced by CpG island promoter hypermethylation in colon cancer cell lines; this suggests that DKK-1 may act as a tumor suppressor (2,5). Consistent with this idea, DKK-1 suppresses cell growth and induces apoptotic cell death by activating the JNK pathway in human mesothelioma cells (6), and DKK-1 suppresses tumorigenicity in cervical carcinoma HeLa cells (7). In contrast, recent studies have shown that DKK-1 is overexpressed in hepatoblastoma, Wilms' tumors and hepatocellular carcinoma (8,9). However, the role of increased DKK-1 in these tumors is not known. Here, we examined the DKK-1 expression in hepatocellular carcinoma (HCC) cell lines and we studied the effect of DKK-1 overexpression in HCC cell lines.
MATERIALS AND METHODS
1) Cell lines
Cell lines derived from hepatocellular carcinomas (HCCs) were purchased from the Korean Cell Line Bank (Seoul). Hep G2 and Hep 3B cells were grown in DMEM with 10% fetal bovine serum (FBS). The SNU 354, SNU 368, SNU 398, SNU 423, SNU 449 and SNU 475 cells (10) were cultured in RPMI 1640 with 10% FBS.
2) Construction of a dickkopf-1 (DKK-1) expression vector
DKK-1 cDNA was prepared by RT-PCR and then it was subcloned into the EcoRI and XhoI sites of the pcDNA3.1/Myc-His (+) B vector (Invitrogen, Carlsbad, CA) in the sense orientation, generating pDKK-1/Myc-His. The orientation and accuracy of the sequences were confirmed by sequencing. The primers for PCR were 5'-GGAATTCCCTGAGATGATGGCTCTGGGCG-3' and 5' -CCGCTCGAGCGGTGTCTCTGACAAGTGTG-3'. The EcoRI (GAATTC) and XhoI (CTCGAG) sites are underlined.
3) Construction of stable cell lines
The SNU 475 cells were grown to 50~80% confluence and then they were transfected with the control vector (pcDNA3.1/Myc-His (+) B vector) or with the pDKK-1/Myc-His vector with using Lipofectamine. After 2 days, the cell cultures were diluted 1 : 10 in selection medium (RPMI 10% medium with 400 µg/ml G418), and after 4 weeks, the individual and pooled clones were isolated and expanded.
4) RT-PCR analysis
cDNA was synthesized from 2 µg of the total RNA and small aliquots of the cDNA product were amplified with using the primers mentioned above. PCR amplification was performed after an initial denaturation for 5 min at 94℃, followed by 35 cycles for 30 s at 94℃, 45 s at 58℃ and 1 min at 72℃, and final elongation step was done at 72℃ for 10 min.
5) Immunoblotting
The total cell lysates were separated by 10% SDS-PAGE and then transferred to nitrocellulose membranes. The membranes were blocked with 6% milk in PBS for 1 h, and they were probed with mouse anti-β1-integrin antibody (BD Biosciences, Palo Alto, CA) and rabbit anti-β-catenin antibody (Cell Signaling, Beverly, MA). Horseradish peroxidase-conjugated sheep anti-mouse Ig (Amersham, UK) and donkey anti-rabbit Ig were used as the secondary antibodies at a 1 : 5,000 dilution, respectively. The bands were visualized using ECL Plus (Amersham). For the detection of DKK-1/Myc-His fusion protein in the culture supernatant, the conditioned media were collected and concentrated with using Centricon filters (10 kDa cut-off, Millipore, Bedford, MA); they were probed with mouse anti-c-myc-antibody (Roche, Germany).
6) Cell growth assay
The cells were plated at a density of 200,000 cells in 100 mm plates and they were incubated for 4 days. The viable cells were manually counted using a hemocytometer.
7) Analysis of the cell cycle distribution
The cells were cultured in serum-free medium for 4 days. They were collected and washed with PBS and then fixed in 70% ethanol at 4℃ overnight. The cell cycle distribution was analyzed by flow cytometry as was previously described (11).
8) Adhesion assays
The wells of 96-well plates were coated with 10 µg/ml Type I collagen or fibronectin overnight at 4℃, and they were blocked with 1.5% BSA for 1.5 h at 37℃. The cells were plated at a density of 20,000 cells/well and then incubated in serum-free medium for 24 h. They were then washed twice in PBS, fixed with 4% paraformaldehyde for 10 min, and stained with 0.1% crystal violet for 20 min. Thereafter, the stained cells were washed with PBS, air-dried and solubilized in 50 µl of 100 mM HCl; the optical density was measured at 630 nm.
9) Migration and invasion assays
30,000 cells in 100 µl of RPMI medium were added to the upper compartment of MiliCell chambers that had 8 µm pores, and the chambers were placed into 24-well culture dishes that contained 500 µl of 10% RPMI medium. After 2 h at 37℃, the cells on the upper side of the filter were removed with a cotton swab and those attached to the underside were stained with 0.1% crystal violet; following this, they were photographed and counted. For the invasion assays, growth factor-reduced Matrigel (BD Biosciences) was diluted (30 µg in 100 µl of H2O); this was added to the top chamber and allowed to gel for 1 h at 37℃, air-dried for 16 h, and the Matrigel barrier was reconstituted for 2 h at 37℃ prior to use. After 24 h at 37℃, cells were counted in the same manner as for the migration assay.
RESULTS
1) Expression of DKK-1 in the hepatocellular carcinoma cell lines
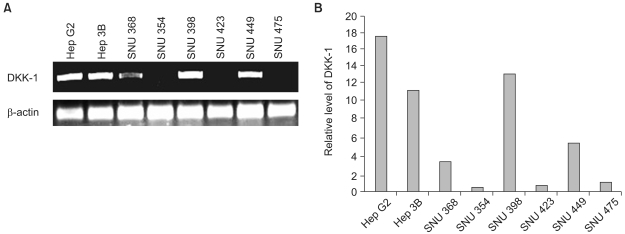
We investigated the DKK-1 expression in 8 human hepatocellular carcinoma cell lines by performing RT-PCR analysis. As shown in Fig. 1, we identified a DKK-1 expression in 63% (5/8) of the HCC cell lines. The Hep G2, Hep 3B, SNU 398 and SNU 449 cells expressed high levels of DKK-1. The SNU 368 cells expressed a moderate level of DKK-1, whereas the SNU 354, SNU 423 and SNU 475 cells expressed very low or no detectable levels of DKK-1.
2) The forced expression of DKK-1 reduces the β-catenin level in SNU 475 cells
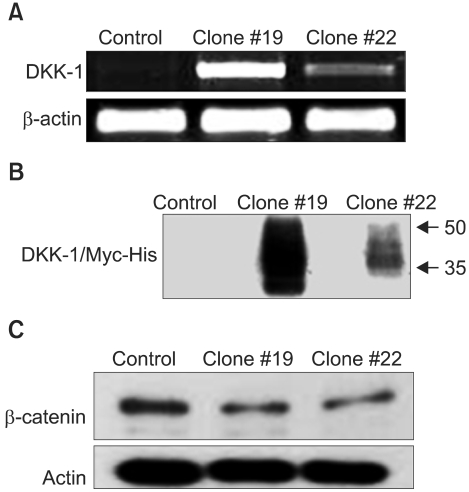
To assess the role of the overexpressed DKK-1 in the biology of hepatocellular carcinoma cells, we transfected SNU 475 cells with a DKK-1 expression vector (pDKK-1/Myc-His). The individual G418 resistant clones were isolated after a few weeks and the expression of DKK-1 was examined. We eventually selected two clones (Clone #19 and Clone #22) for further study. SNU 475 cells were also transfected with an empty vector (pcDNA3.1/Myc-His (+) B) and the G418 resistant clones were pooled and named as the 'control'. The Clone #19 cells expressed a higher level of DKK-1 mRNA than did the Clone #22 cells (Fig. 2A). Consistent with this, Clone #19 secreted higher levels of DKK-1/Myc-His fusion protein into the culture medium (Fig. 2B). A smeared signal around 35 KDa on the immunoblotting indicates that the secreted DKK-1 fusion protein is subjected to post-translational modification, as is the case for the wild-type protein (3,12). The SNU 475 cells expressing DKK-1 displayed a significantly reduced level of β-catenin, demonstrating the inhibition of the canonical Wnt pathway by DKK-1 fusion protein, and this resulted in β-catenin destabilization (Fig. 2C).
3) Forced expression of DKK-1 suppresses the growth of SNU 475 cells
Measurements of the effect of DKK-1 on cell growth revealed that cell growth was suppressed in the clones that expressed DKK-1 (Fig. 3A). The Clone #19 cells expressing a higher level of DKK-1 showed more suppression than did the Clone #22 cells. In addition, the clones expressing DKK-1 were more susceptible to apoptosis, as evidenced by the increased population of cells in the sub-G1 phase, which represented apoptotic cells (Fig. 3B).
4) The forced expression of DKK-1 reduces the adhesion capacity, but it stimulates migration
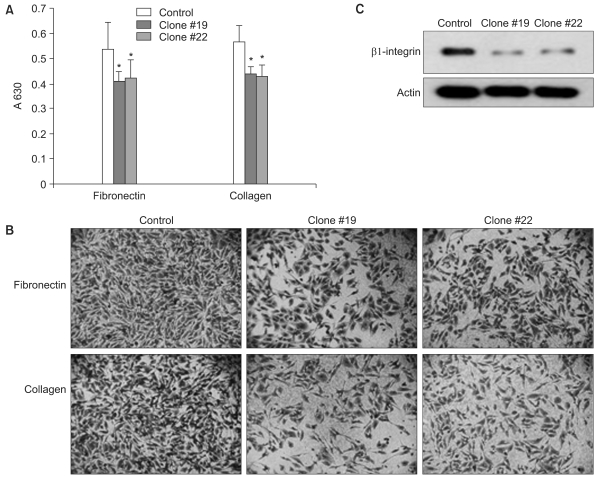
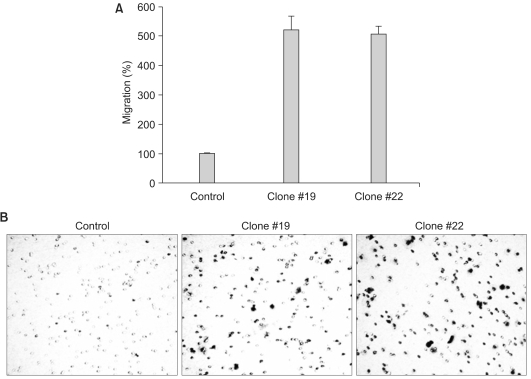
We examined whether the forced expression of DKK-1 alters cell-to-matrix adhesion. The SNU 475 cells expressing DKK-1 displayed significantly reduced adhesion to both collagen type I and fibronectin (Fig. 4A, B). In line with this finding, we found that β1-integrin, which is a transmembrane receptor that binds to fibronectin and collagen, is significantly decreased in the cells expressing DKK-1 (Fig. 4C). We also checked whether DKK-1 affects cell motility and invasiveness. The DKK-1 expressing cells (Clone #19 and #22) had a greater migratory capacity than did the control cells (Fig. 5). However, cellular invasiveness was not affected by DKK-1, as examined by the invasion assay (Fig. 6).
DISCUSSION
In this study, we observed that DKK-1 is expressed in 63% (5/8) of the HCC cell lines and the overexpression of DKK-1 caused reduced cell adhesion as well as increased cell migration, which are the hallmarks of tumor promotion. Our findings are in conflict with recent reports (2,5) in which the DKK-1 expression is downregulated in human colon cancer and it is transcriptionally silenced by CpG island promoter hypermethylation in colon cancer cell lines, which suggests that DKK-1 may act as a tumor suppressor. However, our finding is in line with the overexpression of DKK-1 in hepatoblastoma, Wilms' tumors and hepatocellular carcinoma (8,9).
Because DKK-1 is one of the target genes of the canonical Wnt/β-catenin signaling pathway (4), the overexpression of DKK-1 in some HCC cell lines may represent a negative feedback mechanism of uncontrolled Wnt signaling, as has been suggested in the case of hepatoblastoma and Wilms' tumors (8). In this regard, it is worthy to note that activatingβ-catenin mutations have been found in 8 of 31 HCCs (13), and some subgroups of HCCs are strongly related to β-catenin mutations (14). Also, it has been reported that aberrant nuclear accumulation of β-catenin is observed in ~30~40% of HCCs (15~17) and the Wnt receptor frizzled-7 is overexpressed in human HCC (18), leading to activation of the Wnt pathway.
We also observed that the forced expression of DKK-1 suppresses the growth of SNU 475 cells, and the cells expressing DKK-1 were more susceptible to apoptosis, as evidenced by the increased population of cells in the sub-G1 phase. These results are in line with the finding that DKK-1 overexpression in brain tumor cells and HeLa cervical carcinoma cells increases their sensitivity to apoptosis following alkylation damage of their DNA and UV irradiation, respectively (7,19).
The decreased adhesion to fibronectin and collagen by DKK-1 expressing cells is thought to be due, at least in part, to the down-regulation of β1-integrin. This is in line with the finding that HeLa cells expressing DKK-1 had a ~30% reduction in plating efficiency (7). It is also intriguing that DKK-1 increases cellular migration. We guess it is possible that the increased migration of cells expressing DKK-1 may be due to the reduced level of β-catenin by inhibition of the canonical Wnt pathway via DKK-1 (Fig. 2C). In this regard, it is interesting to note that β-catenin not only plays a role as a coactivator of TCF proteins, but it also binds directly to the intracellular domain of E-cadherin that mediates adhesion between cells (20). On the other hand, DKK-1 did not increase cellular invasiveness. This result suggests that DKK-1 does not affect synthesis of proteases such as matrix metalloprotease-2 (MMP-2) that acts on collagen type IV, a major component of the Matrigel matrix used in this study. We have also found, by performing RT-PCR analysis, that the level of MMP-2 did not change in Clones #19 and #22 (data not shown).
CONCLUSIONS
In this study, we found that DKK-1 is expressed in some HCC cell lines and the forced expression of DKK-1 caused growth inhibition. On the other hand, the overexpression of DKK-1 resulted in reduced cell adhesion as well as increased cell migration, which are the hallmarks of tumor promotion. Collectively, our data suggest that overexpression of DKK-1 affects the biology of HCC cells, although further in vivo studies are needed to confirm the in vitro data presented here.

 XML Download
XML Download