

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
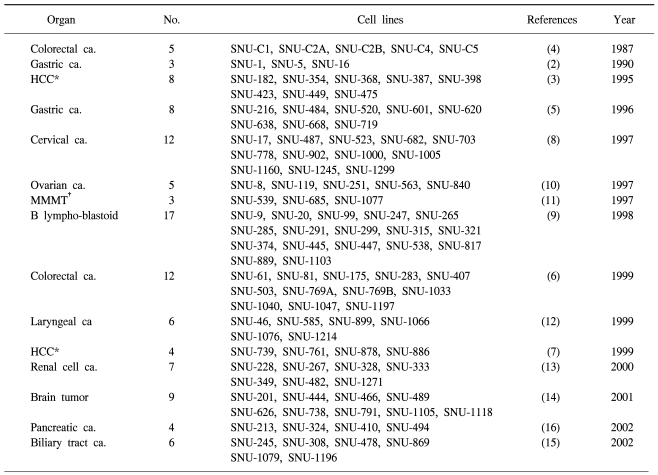
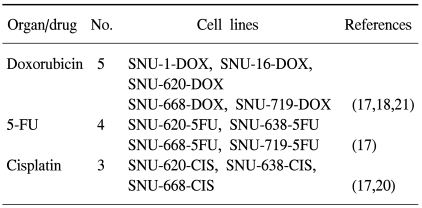
Cell lines are important because they provide a consistent renewable source of cell material for study. Cell line models should reflect the properties of the original cancers, such as, the maintenance of histopathology when transplanted into immunodeficient mice, genotypic and phenotypic characteristics, gene expression, and drug sensitivity (1). Moreover, advances in cell culture methods have made it possible to establish a variety of human cancer cell lines from surgical tissues, body fluids, and biopsy specimens. Because pure cells in culture can be used in many studies that cannot be conducted using tissue specimens, permanent cell lines established from human cancers offer great experimental flexibility (2~5). The Laboratory of Cell Biology at the Cancer Research Institute of Seoul National University (SNU) College of Medicine, has consistently worked to establish cell lines from human cancers for over 20 years. And currently, 180 tumor cell lines have been established from 2,200 cancer tissues. Of these 109 have been characterized and described in the literature. These include, 17 colorectal carcinoma (4,6), 12 hepatocellular carcinoma (3,7), 11 gastric carcinoma (2,5), 12 uterine cervical carcinoma (8), 17 B-lymphoblastoid (9), 5 ovarian carcinoma (10), 3 malignant mixed Müllerian tumor (11), 6 laryngeal squamous cell carcinoma (12), 7 renal cell carcinoma (13), 9 brain tumor (14), 6 biliary tract (15) and 4 pancreatic carcinoma (16) cell lines (Table 1). We have also established several anticancer drug resistant cell lines in the hope of identifying genes associated with anticancer drug sensitivity or resistance (17~21) (Table 2). And in 1987, we established the Korean Cell Line Bank (KCLB, http://cellbank.snu.ac.kr) at Cancer Research Institute in Seoul National University College of Medicine to distribute suitable cell lines to life science researchers. The characteristics of SNU cell lines have been reported in over 180 international journals by our laboratory and by other researchers since 1987 (references to these studies are included in the Reference section). The genetic, epigenetic and expression alterations of genes in SNU cell lines are summarized in Tables 3~6. In fact, several characteristics of SNU cell lines were reported in 2002 (22). In addition, we have contributed to three text books on the establishment and characterization of gastric, colorectal, and renal cancer cell lines (23~25). In this paper, we summarize the more recently discovered cellular and molecular characteristics of these cell lines and include the characteristics of recently developed cell lines. We also provide a summary on the establishment of cancer cell lines.



Go to : 
ESTABLISHMENT OF CANCER CELL LINES
This section provides a summary of the establishment of cancer cell lines, for more detail refer to references 23~25. Our laboratory has extensive experience of the use of serum-containing and fully-defined media for the establishment of permanent cell lines from human cancers. Most of the SNU cancer cell lines developed were cultured initially in ACL-4 medium supplemented with 5% heat-inactivated FBS (AR5). ACL-4 is a fully defined medium, which was formulated for the selective growth of human lung adenocarcinoma cells, and which proved to be useful during the establishment of colorectal cancer cell lines. ACL-4 is a complex medium, and consists of 12 additives including basal medium (Table 7).

Human cancer cell lines can be established from ascitic effusions, metastatic tissues (regional lymph nodes and distant metastatic sites), and from primary tumors. The establishment of cell lines from ascitic effusions has proven to be more efficient than their establishment from primary tumors, because the cancer cells are rich, free-floating, and primed for in vitro growth. However, effusions usually contain mesothelial cells and lymphocytes. The former can attach more easily to flasks than cancer cells and generally survive for more than one year, whereas lymphocytes can inhibit cancer cell growth. Moreover, the enzymatic digestion of solid primary tumor tissues may result in the poor recovery of viable cells and in the overgrowth of the contaminating faster growing stromal cells. The mechanical spillout method provides a simpler, faster, and less traumatic method of obtaining cells for culture. This method minimizes stromal cell contamination, because stromal cells are not easily detached from tissue matrix by mechanical means.
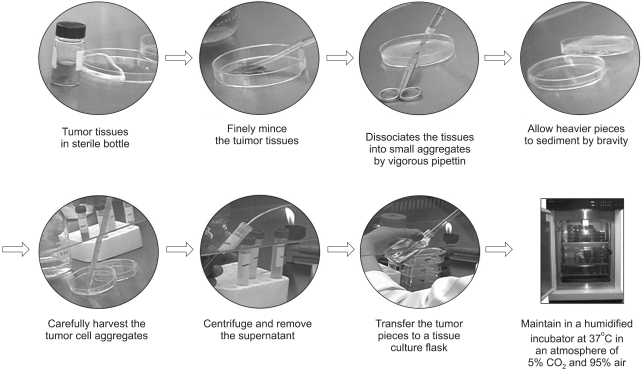
Solid tumors (primary tumors and tumors in lymph nodes) must be carefully and aseptically excised from pathologically proven human cancer samples and transferred to a cell culture laboratory in RPMI1640 medium. Whenever possible, invasive areas from the serosal surface should be selected for primary tumor cultures, to reduce the possibility of microbial contamination. The RPMI1640 medium used to transport the specimen should contain antibiotics, such as penicillin-streptomycin, to prevent bacterial contamination, and ascitic effusions should be collected in sterile glass bottles or syringes. Heparin (5 ug/ml) may be added to collected fluids to prevent clotting, if the effusions contain many erythrocytes. Cancer cells can grow either as floating aggregates, firmly, as loosely adherent colonies, or as both adherent and floating subpopulations. If cancer cells are adherent or grow as floating aggregates, the medium is changed weekly until a substantial outgrowth of cells is observed. Cancer cells are isolated when heavy tumor-cell growth is observed. The key features for successful culture are as follows: 1) nonenzymatic or minimal dissociation of tumor tissue; 2) seeding cultures as explants and at high cell densities; 3) the removal of contaminating fibroblasts, usually after they have aided culture initiation; 4) delaying passage until high cell densities have been achieved and 5) plating cells at high density.
Several methods can be employed to isolate pure cancer cells. After establishing cancer cell lines, initial passages are performed when heavy tumor-cell growth and large colonies are observed. Subsequent passages are performed 1 or 2 weekly. ACL-4 medium supplemented with heat inactivated 5% FBS, which is used in primary cell culture can be also used to propagate cancer cell lines. This medium must be used until the isolated cancer cells are considered a cell line. It is recommend that early passages, whenever passaged, are frozen in cryoprotective medium to prevent loss of the cell lines due to contamination or other laboratory accidents. After cells have been fully established, AR-5 medium can be replaced with RPMI 1640 medium supplemented with 10% heat inactivated FBS (Fig. 1). The growth characteristics of some cultured SNU cell lines are shown in Fig. 2.
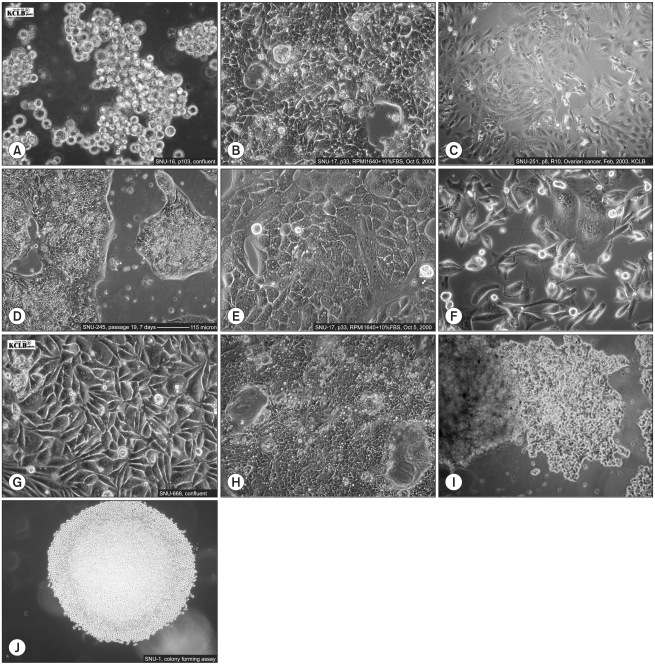 | Fig. 2Culture of SNU cell lines. (A) SNU-16 gastric cancer cell line (B) SNU-17 uterine cervical carcinoma cell line (C) SNU-251 ovarian carcinoma cell line (D) SNU-245 biliary tract cancer cell line (E) SNU-368 hepatocellular carcinoma cell line (F) SNU-482 renal cell carcinoma cell line (G) SNU-668 gastric carcinoma cell line (H) SNU-1033 colorectal carcinoma cell line (I) SNU-175 colorectal carcinoma cell line (J) SNU-1 gastric cancer cell line cultured in soft agar.
|
Go to :
CHARACTERISTICS OF SNU CANCER CELL LINES
1) SNU colorectal cancer cell lines
SNU colorectal carcinoma cell lines have become items in the research field (22,26~44). Seventeen cell lines (SNU-C1, SNU-C2A, SNU-C2B, SNU-C4, SNU-C5, SNU-61, SNU-81, SNU-175, SNU-283, SNU-407, SNU-503, SNU-769A, SNU-769B, SNU-1033, SNU-1040, SNU-1047, and SNU-1197) have now been established and characterized (4,6). These cell lines have been used for many research purposes, including, multidrug resistance (45) and anticancer drug screening (29,30,38,42,46~54), and in investigations into microsatellite instability (36,55), CEA expression (37), mutations in mismatch repair genes (6), the TGF-β signaling pathway (36), and many others (56~61).
The resistance of cultured cells to drugs, such as, the vinca alkaloids and the anthracyclins is frequently due to the expression of P-glycoprotein, which is encoded by the MDR1 gene in humans. P-glycoprotein functions as an energy-dependent drug efflux pump. In SNU gastric and colorectal cancer cell lines, using a slot blot assay, relatively low levels of MDR1 RNA were found to be present in the SNU-1, SNU-5, and SNU-16 gastric cancer cell lines, whereas high levels were found in the SNU-C4 colorectal carcinoma cell line (45).
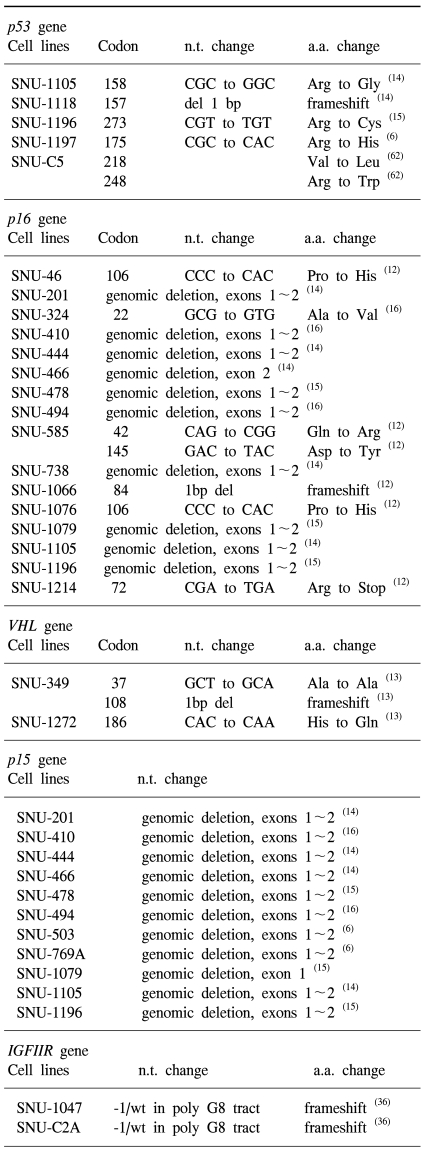
Rand et al. (62) found two missense mutations of the p53 gene in the SNU-C5 colorectal carcinoma cell line, and identified two mutations located at codons 218 and 248 of separate alleles. SNU-C5 cells exhibited complete loss of normal p53 function, as evidenced by the overexpression of p53 and its by the failure to induce p53, p21, and mdm-2 in response to DNA damaging agents.
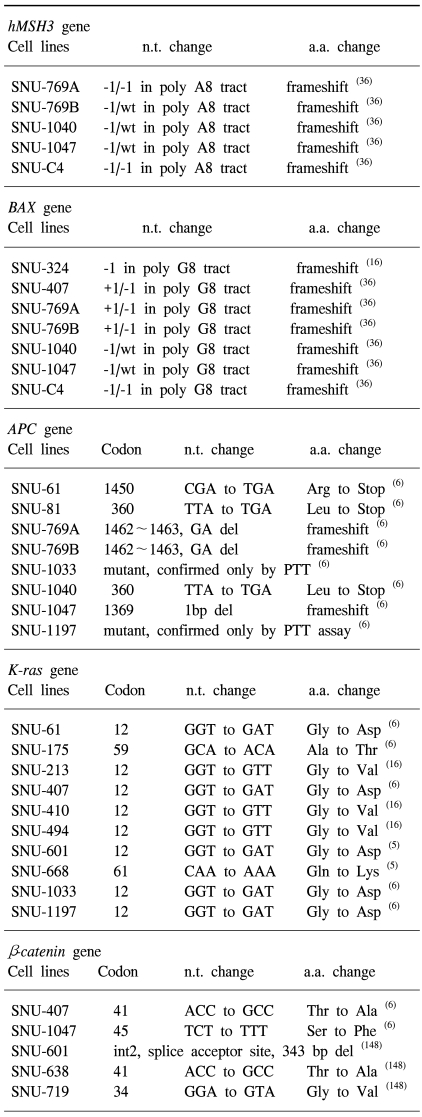
In 1999, we reported upon 12 new colorectal carcinoma cell lines derived from 6 primary tumors and 6 metastatic sites in 11 Korean colorectal-carcinoma patients, their morphologies in vivo and in vitro were described as were mutations of K-ras2, p15, p16, p53, APC, β-catenin, hMLH1, and hMSH2 genes in vitro (6). All 12 lines were found to express surface carcinoembryonic antigen and to secrete it into the supernatant fluid. Morphological correlations between the original tumors and cultured cells suggested that mucinous adenocarcinoma correlated with floating aggregates in culture, and that the degree of desmoplasia in the original tumor correlated with attached growth in culture. Five of the 12 cell lines showed mutations in the K-ras2 gene, and 6 lines showed mutations in the p53 gene. The p15 gene was deleted in 2 cell lines, and the p16 gene was hypermethylated in 3 cell lines. Moreover, mutations of the mismatch-repair genes (hMLH1 and hMSH2) were found in 4 cell lines, and the APC and β-catenin genes were mutated in 9 and 2 lines, respectively.
Using these newly developed colorectal carcinoma cell lines, we assayed mutations in the mononucleotide repeat tract in the coding region of the TGF-β RII gene. Four lines were found to contain deletion or insertion mutations in the 10 base pair adenine repeat tracts. These 4 lines also showed microsatellite instability (36).
A RT-PCR analysis of FGFR3 from human colorectal carcinomas revealed novel mutant transcripts caused by aberrant splicing and by the activation of cryptic splice sequences. Two aberrantly spliced transcripts were detected at high frequency in 50% of 36 primary tumors and in 60% of 10 human colorectal cancer cell lines. Most transcripts used normal splice sites and either skipped or included exons 8 and 9. The predicted translation products are expected to exhibit frameshifts and a premature termination codon in exon 10 (63).
Promoter hypermethylation of the RUNX3 gene was investigated in 16 SNU colorectal cancer cell lines. RUNX3 is not expressed in SNU-61, SNU-81, SNU-175, SNU-769A, SNU-769B, SNU-1047, or SNU-C5 cell lines. Using primers for methylated DNA amplification on bisulfite modified DNA, DNA fragments were amplified from SNU-81, SNU-1047, and SNU-C5 cell lines. In all these cell lines, the 24 individual CpG sites (located between -270 and -121, relative to the transcription initiation site of exon 1 of RUNX3) were found to be fully methylated. In four SNU cell lines (SNU-61, SNU-175, SNU-769A, and SNU-769B cell lines), which did not express RUNX3 mRNA, bisulfite PCR products were not amplified by methylated DNA-specific PCR. RUNX3 mRNA was re-expressed in SNU-81, SNU-1047, and SNU-C5 cell lines after treating them with the demethylating agent, 5-aza-2'-deoxycytidine (35).
2) SNU gastric carcinoma cell lines
In general, gastric carcinoma cell lines have proven to be more difficult to establish in long-term culture than esophageal or colorectal-carcinoma cell lines (25). A total of 11 gastric carcinoma cell lines (SNU-1, SNU-5, SNU-16, SNU-216, SNU-484, SNU-520, SNU-601, SNU-620, SNU-638, SNU-668, and SNU-719 cell lines) have been established and reported (2,5). SNU gastric carcinoma cell lines are among the most popular in use today. These cell lines have been used in various research fields including multidrug resistance (43,45), anticancer drug screening (47,51,64~81), the establishment and characterization of anticancer drug resistant gastric cancer cell lines (17,18,20,21,82), TGF-β signaling pathway (68,83~91), gene amplification (92~98), telomerase assays (21,99), methylation studies (100~107), cytogenetic studies (108~110), the mutational and expression analysis of FGF receptors (111,112), responsiveness of gastric cancer cells after Helicobacter pylori infection (113~117), studies on mismatch repair genes (118,119), mechanism of apoptosis (71,72,120~124), invasion assays of gastric cancer cells (125~130), mutational analysis of genes (131~135), cDNA and oligonucleotide microarrays (17,131,136~138), proteomic analysis in gastric cancer cell lines (20), and in other studies (139~144).
SNU-16 line has been used as source of amplified DNA sequences (92~96). This line was amplified for the c-myc protooncogene and found to contain four homogeneously staining regions (HSR). c-myc was found to be amplified 50-fold more than in other cell lines (2) and FGFR2 (KSAM) was amplified in extrachromosomal double minute chromosomes (DMS) (92). Fibroblast growth factor (FGF) is fundamentally involved in embryonic development, angiogenesis, and tumorigenesis, and Shin et al. (112) found fibroblast growth factor receptors -2, -3 and 4 were amplified and FGFR2 and 4 were overexpressed in the SNU-16 gastric cancer cell line. They also found that acidic FGF, keratinocyte growth factor (KGF) treatment enhanced the invasive potential of SNU-16 cells versus a control cell line.
Park et al. (87) found several genetic changes in TGF-β RII in SNU gastric carcinoma cell lines resistant to the growth inhibitory effect of TGF-β. This was the first report that TGF-β resistance to cancer cells is associated with genetic changes in the TGF-β RII gene in human gastric carcinoma cell lines. The cell lines SNU-1, SNU-5, and SNU-719 resistant to TGF-β showed no expression or overexpression of the TGF-β RII gene.
Myeroff et al. (86) also reported that mutations in the 10 base pair polyadenine repeat tract (BAT-RII) of the TGF-β RII gene are common in gastric cancer, and demonstrated microsatellite instability using the SNU gastric carcinoma cell lines Concerning the SNU gastric carcinoma cell lines, mutations have been found in BAT-RII of the TGF-β RII gene in SNU-1 and SNU-668. SNU-638 gastric carcinoma cell line showed microsatellite instability (MSI) and frameshift mutations in BAT-RII of the TGF-β RII gene (83). The wild type TGF-β RII transfected SNU-638 cell line responded to TGF-β and tumorigenicity was reduced and delayed when it was transplanted into nude mice. These results suggest a strong association between the expression of wild type TGF-β RII and the degree of malignancy of human gastric cancer cells.
It is known that MutL proteins form the MutL-alpha complex, hPMS2 and hMLH1. Using the SNU-1 cell line, which lacks hMLH1, Leung et al. (145) proved the existence of other MutL protein, namely, the MutL-beta complex, hPMS1 and hMLH1. Moreover, in the SNU-1 cell line, these workers found that lower levels of hMLH1 were associated with markedly lower levels of hPMS2 and hPMS1 proteins, but that the RNA levels of hPMS1 and hPMS2 were normal. They found that hPMS2 and hPMS1 are present in the cells bound to hMLH1, but that hPMS1 and hPMS2 do not associate with each other.
SNU-gastric carcinoma cell lines have also been used in a Helicobacter pylori studies (113-117). After infection with H. pylori, SNU-5 expressed several cytokines, including, interleukins, GM-CSF, MCP-1, and TNF-alpha. H. pylori infection of SNU-5 up-regulated the mRNA and protein levels of COX-2 and stimulated the release of PGE2.
The overexpression of cyclooxygenase-2 has been reported in gastric cancer. However, Yamamoto et al. (146) found that SNU-1 (a gastric carcinoma cell line which is MSI and hMLH1 defective) lacks COX-2 expression, and that after treatment with aspirin or sulindac the MSI phenotype was reduced.
Grady et al. (101) found that several hereditary gastric cancers with germline mutation in the E-cadherin gene have no LOH or somatic mutation. These cancers with germline mutation do not express the E-cadherin gene and the promoter region of the E-cadherin gene proved to be methylated. Therefore, it was concluded that E-cadherin gene promoter methylation acts as a second genetic hit in hereditary gastric cancer. In this study SNU-1 was used as control cell line for E-cadherin promoter methylation. The E-cadherin promoter in SNU-1 proved to be methylated, and after treatment with the demethylating agent 5-aza-2'-deoxycytidine, E-cadherin expression was restored.
To investigate the genetic changes that occurred during the establishment of the 4 SNU gastric cancer cell lines, Bae et al. (147) examined microsatellite instability, loss of heterozygosity, and p53 mutation. They found that MSI, LOH, and p53 gene mutations were sustained during the establishment of the gastric cancer cell lines, and they suggested that minor genetic differences between original tumor tissues and the cancer cell lines could be attributed to tumor heterogeneity, because portions of the original tumor tissues showed similar variations.
Fukuda et al. (109) examined 25 gastric cancer cell lines, including SNU cell lines, to detect aberrations in the DNA copy number used to identify chromosomal gains and losses, and investigated amplifications by comparative Genomic hybridization (CGH). Amplification within 11p13, which bears the CD44 gene, was found in HSC series cell lines of Japanese origin. However, no such amplification was found in SNU gastric cancer cell lines, in which amplification within 8q was frequently observed.
β-catenin is a key molecule in Wnt/wingless signal transduction and in the E-cadherin pathway. Altered expression and mutations of the β-catenin gene have been identified in a number of human malignancies. Woo et al. (148) identified mutations of the β-catenin gene in three (SNU-601, SNU-638, and SNU-719) of the 11 SNU gastric cancer cell lines. Interestingly, β-catenin protein in these three cell lines was located in the cell nucleus, which suggests that β-catenin protein in these three cell lines is complexed with Tcf/lef and translocated to the nucleus from the cytoplasm.
Oh et al. (149) recently found that a naturally derived SNU-719 gastric cancer cell line is infected with type I Epstein-Barr Virus (EBV). EBNA1 and LMP2A were expressed, while LMP1 and EBNA2 are not expressed in this line. EBV infection was also shown in the original carcinoma tissue of this cell line. These findings suggest that SNU-719 cell line may serve as a valuable model system to clarify the precise role of EBV in gastric carcinogenesis.
Many chemotherapeutic agents have been used to treat gastric cancer patients, but the emergence of drug resistance has prevented successful treatment in many cases. The two major forms of drug resistance are intrinsic resistance, in which previously untreated tumor cells are inherently insensitive to the chemotherapeutic agent, and acquired resistance, in which treated tumor cells become insensitive after drug exposure. Our laboratory has developed anticancer drug resistant cell lines from SNU-620, SNU638, SNU-668, and SNU-719 gastric cancer cell lines, namely, four 5-FU-resistant gastric cancer cell lines (SNU-620-5FU, SNU-638-5FU, SNU-668-5FU, and SNU-719-5FU), 5 doxorubicin-resistant cell lines (SNU-1-DOX, SNU-16-DOX, SNU-620-DOX, SNU-668-DOX, and SNU-719-DOX), and 3 cisplatin-resistant cell lines (SNU-620-CIS, SNU-638-CIS, and SNU-668-CIS) (Table 3).
Kang et al. (17) performed global gene expression analysis in these acquired drug-resistant gastric cancer cell lines using an Affymetrix HG-U133A microarray after exposing them to the commonly used drugs 5-fluorouracil, doxorubicin, or cisplatin. They identified over 250 genes differentially expressed in these 5-fluorouracil-, cisplatin-, or doxorubicin-resistant gastric cancer cell lines. Expression analysis also identified eight multidrug resistance candidate genes that were associated with resistance to two or more of the chemotherapeutics tested. This investigation provided comprehensive gene information upon acquired resistance to anticancer drugs in gastric cancer cells and a basis for additional functional studies.
Comparative proteomics involving 2-dimensional gel electrophoresis (2-DE) and matrix-associated laser desorption ionization-mass spectroscopy (MALDI-MS) was performed by Yoo et al. on protein extracts from SNU-620 and SNU-638 gastric cancer cell lines and drug-resistant derivative lines to screen for drug resistance-related proteins (20). Pyruvate kinase M2 (PK-M2) was identified as a protein with lower expression in cisplatin-resistant cells than in parental cells. They also found that PK-M2 activity in 11 individual human gastric carcinoma cell lines was positively correlated with cisplatin sensitivity, and concluded that PK-M2 protein and activity levels are lower in cisplatin-resistant human gastric carcinoma cell lines than in their corresponding parental cell lines.
Han et al. (150) observed that Smad3 was expressed at low to undetectable levels in SNU-1 and SNU-484 cell lines, and that it was detected in SNU-5, SNU-16, SNU-601, SNU-620, SNU-636, SNU-668 and SNU-719 cell lines. They found that the introduction of Smad3 into the SNU-484 cell line restored TGF-β responsiveness, the induction of p21 and p15 gene expression, and the growth inhibition response to TGF-β. They also observed that Smad3-expressing cells showed markedly lower and delayed tumorigenicity in vivo.
The aberrant methylation of normally unmethylated CpG islands located in the 5' promoter region of genes is associated with transcriptional inactivation, which is as effective as inactivation by gene mutation or deletion. The aberrant methylation status of some genes was examined in SNU gastric cancer cell lines. Song et al. (106) found that the p16 gene was not expressed mRNA and that CpG islands of the promoter region of this gene were hypermethylated in SNU-1, SNU-601, SNU-620, SNU-638, and SNU-719 cell lines. They also found that COX-2 was not expressed and was hypermethylated in SNU-601, SNU-620, and SNU-719 cell lines (105). CpG islands of the promoter region of the RUNX3 gene were found to be hypermethylated in SNU-1 and unmethylated in SNU-5, SNU-16, and SNU-719 cell lines (151). DLC-1 was completely methylated in SNU-1, SNU-16, SNU-601, and SNU-791 cell lines, and partially methylated in SNU-620 cell line (103). Moreover, Integrin α4 was found to be unmethylated in SNU-484, partially methylated in SNU-668, and completely methylated in SNU-1, SNU-5, SNU-601, SNU-638, and SNU-719 (104). Choi et al. (100) recently reported that AKAP12A is completely methylated in SNU-1, SNU-601, and SNU-638 and partially methylated in SNU-16, SNU-620, and SNU-719, and that AKAP12B is completely methylated in SNU-1, SNU-484, SNU-620, and SNU-719 and partially methylated in SNU-601.
3) SNU hepatocellular carcinoma cell lines
SNU hepatocellular carcinoma cell lines have proven to be of value in carcinogenic studies associated with hepatocellular carcinogenesis and hepatitis B virus X (43,152,153). Our laboratory has developed and reported upon 12 hepatocellular carcinoma cell lines (SNU-182, SNU-354, SNU-368, SNU-387, SNU-398, SNU-423, SNU-449, SNU-475, SNU-739, SNU-761, SNU-878, and SNU-886) (3,7). At present, these cell lines have been widely used to investigate HBV expression, IGF expression (153), and methylation status of genes (154,155), and other issues (51,155~166) in hepatocellular carcinogenesis.
Two SNU hepatocellular carcinoma cell lines SNU-354 and SNU-354 have been shown to express hepatitis B virus and HBV X transcripts (3). These cell lines have been used widely in the study of hepatocellular carcinogenesis and hepatitis B virus. SNU-354 and SNU-368 hepatocellular carcinoma cell lines have been found to express high levels of the MDR1 gene (43).
The hepatocellular carcinoma cell line, SNU-368, has already been shown to express hepatitis B virus X antigen, and Lee et al (167) reported that the expression of HBX is induced by hypoxia in SNU-368 cells. These workers also found that HBX stimulates the transcription of vascular endothelial growth factor (VEGF mRNA), a potent angiogenic factor in hypoxia, which suggests that HBX may play a critical role in hypoxia induced angiogenesis through the transcriptional activation of VEGF during hepatocarcinogenesis.
The integration statuses of the hepatitis B virus (HBV) into the human telomerase reverse transcriptase (hTERT) gene in five SNU HBV-positive hepatocellular carcinoma cell lines (SNU-387, SNU-387, SNU-423, SNU-449, and SNU-475) were investigated. In SNU-449, the 1452bp initial RS (restriction site)-PCR product containing the HBV/cellular DNA junction sequence had a HBV sequence of 815bp joined to a 637bp sequence from intron 3 of the hTERT gene. An analysis of the sequence of the entire HBV integration with flanking human host sequences revealed a 5bp deletion in hTERT intron 3. Moreover, hTERT mRNA expression was upregulated in SNU-449 versus benign liver tissue. This work suggests that the sites of oncogenic viral integration are nonrandom and that genes at the sites of viral integration may play important roles in carcinogenesis (168).
Park et al. (163) recently found that TGF-β treatment significantly enhances the viability of SNU-354, SNU-475, and SNU-449 cell lines, and they showed that these cell lines retain high endogenous c-Src activity. Moreover, SNU-449 cell line demonstrates an elevated c-Src activity despite its relatively low c-Src protein level. These results indicate the possibility that the responsiveness of hepatocellular carcinoma cells to the growth inhibitory effects of TGF-β1 may be controlled by c-Src activity.
4) SNU uterine cervical carcinoma cell lines
We have developed and reported upon 12 uterine cervical carcinoma cell lines (SNU-17, SNU-487, SNU-523, SNU-682, SNU-703, SNU-778, SNU-902, SNU-1000, SNU-1005, SNU-1160, SNU-1245, and SNU-1299) (8). These 12 carcinoma cell lines of the human uterine cervix were established from 5 keratinizing and 5 nonkeratinizing squamous-cell carcinomas, and 2 small-cell carcinomas. Ten of these cell lines grow as adherent cells and 2 as floating aggregates. All 12 SNU uterine cervical carcinoma cell lines are infected by human papilloma virus (HPV), seven by HPV-16, three by HPV-18, one by HPV-31, and one by HPV-33. Interestingly, 6 of the 7 cell lines containing HPV-16-type DNA harbor the same alteration of E7 at nucleotide position 647 (amino acid 29, AAT→AGT, Asn→Ser), whereas the 3 HPV-18-positive lines do not. Theses 3 cell lines have been proven to contain intact E1/E2 of HPV, suggesting the presence of episomally replicating HPV DNA as well as the integrated form, whereas the other 9 lines have been shown to have integrated HPV. Most of the HPVs are integrated into the genome of the cells. Episomally replicating HPV infected uterine cervical carcinoma cell lines are rare, but the SNU-1000 cell line contains the episomal form and the nonepisomal form of HPV. These SNU uterine carcinoma cell lines have also been used in carcinogenesis studies associated with cervical carcinogenesis and human papilloma virus.
BPV1 E2 was expressed in SNU-17, SNU-902, and SNU-1299 cell lines after the acute infection of an SV40-BPV1 recombinant virus. EPV1 E2 expression caused a significant decrease in E6/E7 transcription in these cell lines, which was accompanied by an increase in the level and activity of p53 protein and a decrease in the expression of Cdc25A, a Cdk2-activating phosphatase; moreover, E2F1 activity and cellular DNA synthesis capacity were significantly reduced. These results indicate that the inhibition of E6/E7 gene expression in HPV16-positive cervical carcinoma cells suppresses cell proliferation by activating the growth inhibitory factors p53 and Rb, and also by downregulating the cell cycle stimulatory factor Cdc25A (169).
5) SNU ovarian carcinoma cell lines
In 1997, we reported upon five human ovarian carcinoma cell lines (SNU-8, SNU-119, SNU-251, SNU-563 and SNU-840) (10). Of these, SNU-251 harbors mutations in p53, BRCA1, and hMLH1. Of these five cell lines, we consider that SNU-251 is both unique and valuable. This cell line has a nonsense mutation in the BRCA1 gene at codon 1815 in exon 23, which results in a truncated protein. It is the only reported ovarian carcinoma cell line with a nonsense mutation in this gene. SNU-251 has proven valuable in studies of the function of BRCA1 (170~174).
The role of BRCA1 in the development of ovarian cancer is poorly understood, partially owing to a lack of ovarian cancer cell lines with defective BRCA1. Zhou et al. (171) recently further characterized the SNU-251 cell line to determine the role of BRCA1 in resistance to both ionizing radiation and paclitaxel. The BRCA1 mutation in the SNU-251 cell line is predicted to cause the loss of the C-terminal 49 amino acids of BRCA1 protein, which contains the second half of BRCT (the BRCA1 C-terminal) repeat. Western blot analysis detected a 210-KDa BRCA1 protein and a high mutant p53 protein expression level in SNU-251 cell line. This mutation of BRCA1 caused a loss of the transcriptional activation of the endogenous p21 gene, and prevented sustained arrest in the G2/M phase of the cell cycle. Moreover, the BRCA1 mutation inhibited BRCA1 subnuclear assembly for DNA-damage repair and increased cellular sensitivity to ionizing radiation and paclitaxel. These results suggest that the deletion of the C-terminal 49 amino acids of BRCA1 results in a loss of BRCA1 function in SNU-251. Therefore, SNU-251 may be a useful model for studying the molecular mechanisms of BRCA1 with respect to the resistance of ovarian cancer to ionizing radiation and chemotherapy, and for studying the development of hereditary human ovarian cancer.
6) SNU uterine malignant mixed mllerian tumor
We have also reported on the establishment and characterization of three cell lines (SNU-539, SNU-685, and SNU-1077) derived from uterine malignant mixed Müllerian tumor (MMMT). SNU-539 and SNU-1077 cells stain positively for both epithelial and mesenchymal antigens, whereas SNU-685 cells only stain positively for mesenchymal antigens. One missense mutation from AAC to GAC at codon 239 of exon 7 in the p53 gene was identified in SNU-539. These permanent uterine MMMT cell lines should be considered as a valuable resource for a multitude of in vitro investigations, and could be used for clarifying the obscure histogenetic origin and the biologic behavior of this aggressive tumor (11).
7) SNU B-lymphoblastoid cell lines from cancer patients
Seventeen lymphoblastoid cell lines (SNU-9, SNU-20, SNU-99, SNU-247, SNU-265, SNU-285, SNU-291, SNU-299, SNU-315, SNU-374, SNU-445, SNU-447, SNU-538, SNU-817, SNU-889, and SNU-1103) have also been established from cancer tissues obtained from Korean patients. During the establishment of tumor cell lines from cancer tissues, outgrowths of lymphoblasts growing as cell clumps in suspension were noted, in the absence of tumor cells growth. These lymphoblastoid cells were EBV-transformed cells, and arose from EBV-infected B cells present in the incubated cancer tissues. The presence of latently-infectious EBV in these cell lines was verified by the detection of the EBNA1gene and EBV-encoded nuclear antigens (EBNA1 and -2), and by the inducibility of an EBV early antigen (9). These cell lines have been found to be useful for studies on the role of EBV in carcinogenesis (149,175~178).
8) SNU laryngeal squamous cell carcinoma cell lines
A total of six human laryngeal squamous cell carcinoma cell lines (SNU-46, SNU-585, SNU-899, SNU-1066, SNU-1076, and SNU-1214) established from Korean patients have been described (12). All cell lines grew as adherent cells; five lines as monolayers and one line (SNU-585) as stratified colonies. p53 mutations were found in SNU-46 (Gly 245 Cys), SNU-585 (Arg 273 Leu), and in SNU-1066 (a deletion of 46bp from nucleotides 514 to 559 resulting in a premature stop codon at 62bp down stream. p16 mutations were observed in five cell lines, including a one base deletion resulting in a premature stop codon in SNU-1066 and a stop codon in SNU-1214 cell lines. Two mutations in TGF-β RII were found in two lines: SNU-46 cell line has a frameshift mutation in exon 3, and SNU-1066 has a missense mutation (Ser 409 Phe) in the kinase domain. These laryngeal squamous cell carcinoma cell lines have been found useful by those investigating the biologic characteristics of laryngeal cancer (179~183). SNU-1041 cell line derived from pharyngeal cancer was also established in our laboratory. In addition, these cell lines were used to study the effect of nitric oxide on COX-2 overexpression, and the inhibitory effect of p27KIP1 gene transfer in head and neck squamous cell carcinoma (179).
9) SNU renal cell carcinoma cell lines
The establishment and characterization of a total of 7 SNU renal cell carcinoma cell lines were reported in 2000 (13). Five of these cell lines were derived from clear cells (SNU-228, SNU-267, SNU-328, SNU-349, and SNU-1272), one from granular cells (SNU-482), and one from a mixed clear and granular cell type (SNU-333). Mutations of the VHL gene were found in SNU-349 (a frameshift mutation at codon 108 resulting in a premature truncating protein) and in SNU-1272 (His 186 Gln), and in their tumor tissues. The SNU-267 line has a frameshift mutation (at codon 219 resulting in a prematurely truncated protein) in the p53 gene. A missense mutation (Cys 121 Arg) of the TGF-β RII gene was detected in the SNU-1272 line and in the corresponding tissue. Analysis of the repeat sequences of the TGF-β RII gene showed one cell line (SNU-349) to have MSI (microsatellite instability) and the other 6 microsatellite stability. As MSI is a hallmark of the inactivation of mismatch repair genes, the presence of hMSH2 and hMLH1 mutations was investigated in all 7 cell lines. An inactivating homozygous single base-pair deletion of the hMLH1 gene (a frameshift mutation at codon 711 resulting in a premature truncating protein) was found only in SNU-349 cell line and in corresponding tissue. Moreover, a frameshift mutation within an 8-bp polyadenine repeat present in the hMSH3 coding region was found only in the MSI cell line and tumour tissue. These well characterized renal cell carcinoma cell lines should provide a useful in vitro model for studies related to human renal cell carcinoma.
10) SNU brain tumor cell lines
In 2001, we reported on the characteristics of newly-established human brain tumor cell lines including six glioblastoma (SNU-201, SNU-444, SNU-466, SNU-489, SNU-626, and SNU-1105), two oligodendroglioma (SNU-738 and SNU-791), and one primitive neuroectodermal tumor (PNET) (SNU-1118) (14). All were established from pathologically proven brain tumors. In terms of p53 mutations the following were found; a point mutation at a splice acceptor of intron 4 in SNU-201, a missense mutation (Cys 275 Tyr) in SNU-444, a missense mutation (Pro 77 Arg) in SNU-626, and a one base C deletion at codon 157 resulting in a premature truncating protein in SNU-1118. In terms of PTEN mutations, a missense mutation (Arg 130 Gln) in SNU-626 and a genomic deletion from exons 49 in SNU-1105 were identified. The mRNA of PTEN in SNU-1105 was also absent. Homozygous deletion of the p15 gene was observed in the SNU-201, SNU-444, SNU-466, and SNU-1105 cell lines. And, the homozygous deletion of the p16 gene wholly or in part, was observed in SNU-201, SNU-444, SNU-466, SNU-738, and SNU-1105. This suggests that alterations of both or either of the p16 and p15 genes are common genetic alterations in human brain cancer cell lines. DCC expression was absent in seven cell lines (SNU-201, SNU-466, SNU-489, SNU-626, SNU-738, SNU-791, and SNU-1119). Alterations in BAT-25, BAT-26, and BAT-40 were observed in all cell lines, indicating that all have microsatellite stability. It is hoped that these cell lines will be found useful tools by those investigating the biological characteristics of human brain tumors (184).
11) SNU biliary tract cancer cell lines
Human cell lines established from biliary tract cancers are rare; only 5 have been reported previously. We recently described the characterization of six new biliary tract cancer cell lines (designated SNU-245, SNU-308, SNU-478, SNU-869, SNU-1079, and SNU-1196), which were established from primary tumor samples taken from Korean patients (15). These cell lines were isolated from two extrahepatic bile duct cancers [one adenocarcinoma of common bile duct (SNU-245), one hilar bile duct cancer (SNU-1196), two adenocarcinomas of ampulla of Vater (SNU-468 and SNU-869), one intrahepatic bile duct cancer (cholangiocarcinoma) (SNU-1079), and one adenocarcinoma of the gall bladder (SNU-308). p53 mutations were found in SNU-478 (266~267insT), SNU-869 (Asp 48 Gly), and SNU-1196 (Arg 273 Cys) and homozygous deletions of both p16 and p15 genes were found in 3 lines (SNU-478, SNU-1079, and SNU-1196). The SNU-478 cell line was found to have a heterozygous missense mutation (Asp3 85 Val) in hMLH1, and the E-cadherin gene was found to be hypermethylated in two lines (SNU-478 and SNU-1079). These newly established and well characterized biliary tract cancer cell lines are likely to be found highly useful for studying the biology of biliary tract cancers. Of these cell lines, SNU-245, SNU-308, and SNU-1079 were subjected to oligonucleotide microarray analysis to identify novel cellular targets in biliary tract cancer (185). These cell lines were also used to validate differentially up-regulated genes by semi-quantitative RT-PCR.
12) SNU pancreatic carcinoma cell lines
We previously reported on the establishment and characterization of four pancreatic carcinoma cell lines (SNU-213, SNU-324, SNU-410, and SNU-494), which were established from histopathologically varied primary (SNU-213, SNU-324, and SNU-494) or liver metastatic (SNU-410) tumor samples from Korean patients (16). Mutations of the K-ras gene were identified in SNU-213 (Gly 12 Val), SNU-410 (Gly 12 Val), and in SNU-494 (Gly 12 Val), and p53 mutations were found within the DNA-binding domain of two, SNU-213 (Arg175His) and SNU-494 (Cys 238 Tyr). Homozygous deletions in both p16 and p15 genes were found in SNU-410 and SNU-494, and SNU-324 was found to have a missense mutation (Ala22Val). No mutations were found in the DPC4 gene (deleted in pancreatic cancer). SNU-324 has a -1-bp or +1-bp mutation in 10-bp polydeoxyadenine repeat tracts of the TGF-β RII gene, and SNU-410 has a genomic deletion of this gene. SNU-324 contains two heterozygous missense mutations in the hMLH1 gene (Cys 233 Arg and Val 384 Asp). In addition, this line showed microsatellite instability and harbored frameshift mutations in simple repeated sequences of the coding regions of the TGF-β RII, BAX, and hMSH3 genes. These cell lines should be useful in studies of pancreatic carcinoma biology.
Go to :
KOREAN CELL LINE BANK
1) Role of Korean Cell Line Bank
Korean Cell Line Bank is a non-profit bioscience organization aimed for distributing the essential cell lines to the life science researchers. The major activities of KCLB are as follows; i) establishment, characterization, deposition, collection, storage, and distribution of cell lines; ii) management of database related to cell lines; and iii) education, workshops, and conferences related to cell lines. KCLB is being supported by the Korean Cell Line Research Foundation (KCLRF). On August 31 in 1993, the KCLRF acquired the status of the International Depositary Authority approved by the World Intellectual Property Organization (WIPO) as specified in Article 6 of the Budapest Treaty on the International Recognition of the Deposit of Microorganisms for the Purpose of Patent Procedure. The KCLB is currently supported by the Korean Science and Engineering Foundation and the Ministry of Science and Technology of Korean. KCLB distributes cell lines throughout Korean.
2) Patent cell lines and SNU cell lines in ATCC
Korean Cell Line Bank has obtained three patents for the invention of cell lines. i) two hepatocelluluar cell lines (SNU-354 and -368) expressing MDR1 gene and hepatitis B virus (Korean patent # 118123), ii) uterine cervical carcinoma cell line (SNU-1000) having intact episomal HPV-16 (Korean patent # 0225491), and iii) ovarian carcinoma cell line (SNU-251) having a nonsense mutation of codon 1815 in exon 23 of the BRCA1 gene (United States patent # 5,948,679). Researchers wishing to obtain these cell lines should contact to the Korean Cell Line Bank and Korean Cell Line Research Foundation (KCLRF). Researchers can also deposit cells for patent application purposes to Korean Cell Line Research Foundation (http://cellbank.snu.ac.kr).
Several gastric, colorectal, and hepatocellular carcinoma cell lines have been deposited in and are also available from the ATCC (American Type Culture Collection, http://www.atcc.org) (Table 8).

3) Distribution of SNU cell lines
All SNU cell lines have been deposited at the KCLB (Korean Cell Line Bank) and are available on request. The KCLB perform quality control on these cell line products, which includes the mycoplasma test, cell viability testing, and DNA fingerprinting analysis.
The order form for SNU cell lines is available from the KCLB website (http://cellbank.snu.ac.kr). To order cell lines from the KCLB, researchers should complete the KCLB order form (MS-Word or PDF if overseas) and fax it to the KCLB. We will on receipt prepare the cell lines and provide information on their characteristics. Cell lines will be delivered in a frozen or cultured state according to customer request. The average time reqired for dispatch is two weeks.
Go to :


 XML Download
XML Download