

 PDF
PDF Citation
Citation Print
Print


Hepatocellular carcinoma (HCC) is the sixth most prevalent neoplasm and is the second most common cause of cancer-related death worldwide [1]. Due to its invasiveness and metastatic potential, demonstration of the pathogenesis of HCC is essential. In addition to environmental factors, a variety of genetic or epigenetic alterations including histone modification are the cause of HCC [2,3]. Histone modification involves post-translational modification of N-terminal tails of histone proteins by acetylation, methylation, phosphorylation, ubiquitylation, sumoylation, adenosine diphosphate ribosylation, biotinylation, and so on [4]. Histone deacetylases (HDACs) are critical regulators of gene expression that promote formation of heterochromatin by deacetylating histone and non-histone proteins. Therefore, aberrant regulation of HDACs contributes to malignant transformation and progression in a wide variety of human cancers, such as HCC, gastric cancer, lung cancer, and other cancers.
Seven sirtuin family members (SIRT1-7) belong to the HDAC, and are highly conserved from bacteria to mammals [5,6]. They are also NAD-dependent, regulating a wide range of cellular processes, including aging, DNA repair, cell cycle, metabolism, and stress response [6,7]. Currently, SIRT1, 2, 3, 6, and 7 are known to be involved with HCC [8-12]. Among them, SIRT7 has been reported to be proportionally upregulated in pre-neoplastic nodules to overt human HCC tissues [12]. In addition, down-regulated H3K18ac has been observed in human HCC samples [13]. SIRT7 specifically deacetylates H3K18ac and contributes to the maintenance of tumor integrity by reducing the transcription of genes at specific loci, including tumor suppressor genes [12,14]. SIRT7 requires cofactors such as ETS-like transcription factor 4 (ELK4). ELK4 belongs to the ternary complex factor subfamily in ETS domain transcription factors [15]. About 60% of the SIRT7-binding sites contain an ELK4-binding motif, and the SIRT7-ELK4 complex induces deacetylation of H3K18ac. Gene-based studies have been performed on SIRT7, but there has been no research on SIRT7 with its deacetylation indicated by immunohistochemical (IHC) study in HCC. Therefore, in the present study we examined the IHC expression of ELK4, SIRT7, and H3K18ac in paraffin-embedded HCC samples.
MATERIALS AND METHODS
Case selection and review
A total of 278 HCC patients, who underwent curative surgery at Korea University Guro Hospital between 2000 and 2013 were enrolled in this study. All hematoxylin and eosin–stained slides were reviewed. To classify the pathologic staging, the American Joint Committee on Cancer (AJCC) staging system (seventh edition) was applied [16]. Tumor differentiation was graded according to the Edmondson-Steiner grading system [17]. Tumor necrosis was evaluated by gross photo and microscopic examination. The results were categorized as ‘present’ when necrosis was detected, even in minor amounts. Missing data were excluded, and clinicopathologic data including sex, age and TNM stage were obtained from patients’ medical records. This study was approved by the Institutional Review Board of Korea University Guro Hospital (KUGH14140-001).
Tissue microarray construction and immunohistochemistry
Representative tumor areas in all patients (n=278), and nontumor areas in 52 patients were obtained and embedded on paraffin blocks. Tissue cores 2.0 mm in diameter were collected from paraffin-embedded donor blocks and embedded in tissue microarray (TMA) blocks. IHC staining was performed on 4 μm TMA section slides using Bond-Max auto-stainer (Leica, Wetzlar, Germany). After incubation with primary antibodies, Bond Polymer Refine Detection kit (Leica) was used for chromogenic reaction. The following primary antibodies were used in this study; ELK4 (1:50, rabbit polyclonal, Novus, Littleton, CO, USA), SIRT7 (1:100, rabbit polyclonal, Abcam, Cambridge, MA, USA), and H3K18ac (1:1000, rabbit polyclonal, Abcam).
IHC assessment
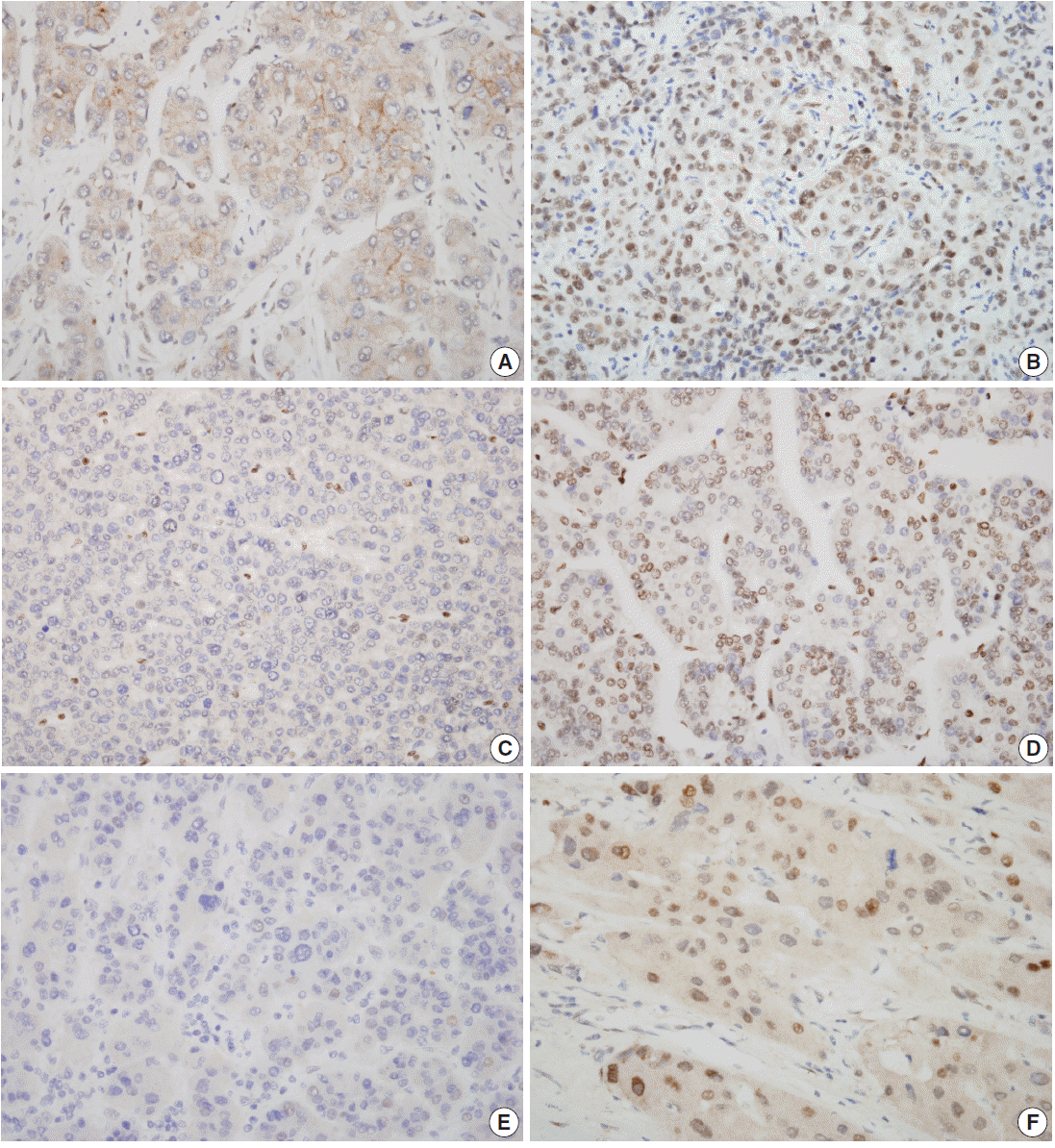
Nuclear staining patterns of tumor cells for ELK4, SIRT7, and H3K18ac were analyzed (Fig. 1). More than 10% of stained tumor cells were categorized as positive. Scoring of intensity was done according to the following scale: 0, no or equivocal; 1, weak; 2, moderate; and 3, strong staining. Cases showing moderate (score 2) or strong (score 3) staining in more than 10% were regarded as “high expression.” Cases with no or weak staining were regarded as “low expression.” This assessment was performed by two pathologists blind to all clinicopathologic data and outcome variables. Due to tissue loss in microarray cores, three samples of SIRT7 (n=275) and two of H3K18ac (n=276) were excluded. None of the ELK4 (n=278) samples were excluded.
Statistical analysis
To examine univariate overall survival, the log-rank test with the Kaplan-Meier survival curve was used. The Cox’s proportional hazards regression model was used for univariate and multivariate analyses of overall and disease-free survival. Overall survival rate was calculated from the date of surgery to the date of the latest follow-up or death. The disease-free survival rate was also calculated from the date of surgery to the date of recurrence on follow-up images or pathologic confirmation. For comparing means, the independent t test was used. Pearson’s correlation coefficient was applied to the correlation between IHC expression and the clinicopathologic data. The p-value less than .05 was considered to be statistically significant. All statistical analyses were performed using the SPSS ver. 20 (IBM Corp., Armonk, NY, USA).
Go to : 
RESULTS
Patient characteristics
In the present study, there were 231 (83%) male and 47 (17%) female HCC patients between 26–84 years of age (median±standard deviation, 57±10.2 years) who underwent curative operation. The median postoperational follow-up period was 34 months. A total of 56 patients (20%) died during the follow-up period. Among all patients, 149 (54%) were in AJCC stage I, 95 (34%) were in stage II, nine (3%) were in stage III, and 25 (9%) were in stage IV.
Expressions of SIRT7, H3K18ac, and ELK4 in non-tumor and tumor cells
The results of all three antibodies showed significant nuclear expression patterns (Fig. 1). In non-tumor hepatocytes of 52 patients, high SIRT7, H3K18ac, and ELK4 expressions were observed in seven (13%), four (8%), and one (2%) patients, respectively; however, in tumor cells, 51 (19%), 143 (52%), and 66 (24%) patients showed high expressions for SIRT7, H3K18ac, and ELK4, respectively (Table 1). Interestingly, H3K18ac showed significantly high expression in tumor cells (mean value, 1.08 and 1.71, respectively; p<.001). In addition, SIRT7 was higher in tumor cells (mean value, 1.06 and 1.08, respectively; p=.858) and ELK4 was higher in non-tumor hepatocytes (mean value, 0.96 and 0.87, respectively; p=.120). However, these results were not significant.
Table 1.
Correlation between immunohistochemical markers and clinicopathologic data
| Characteristic | No. (%) |
Immunohistochemistry expression |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
SIRT7 (n = 275) |
H3K18Ac (n = 276) |
ELK4 (n = 278) |
||||||||
| Low (n = 224, 81%) | High (n = 51, 19%) | p-value | Low (n = 133, 48%) | High (n = 143, 52%) | p-value | Low (n = 222, 76%) | High (n = 66, 24%) | p-value | ||
| Age (yr) | ||||||||||
| < 65 | 214 (77) | 176 | 37 | .721 | 80 | 115 | .184 | 80 | 115 | .184 |
| ≥ 65 | 64 (23) | 50 | 12 | 35 | 28 | 35 | 28 | |||
| Sex | ||||||||||
| Female | 47 (16.9) | 36 | 9 | .448 | 16 | 31 | .021* | 16 | 31 | .033* |
| Male | 231 (83) | 190 | 40 | 117 | 112 | 117 | 112 | |||
| Edmondson-Steiner grade | ||||||||||
| I–II | 178 (64) | 154 | 23 | .002* | 107 | 76 | < .001* | 135 | 43 | .828 |
| III–IV | 100 (36) | 72 | 26 | 26 | 67 | 77 | 23 | |||
| Tumor size (cm) | ||||||||||
| < 5 | 214 (77) | 176 | 37 | .721 | 103 | 111 | .972 | 163 | 51 | .948 |
| ≥ 5 | 64 (23) | 50 | 12 | 30 | 32 | 49 | 15 | |||
| AJCC stage | ||||||||||
| I–II | 244 (87.8) | 199 | 42 | .653 | 119 | 123 | .384 | 183 | 61 | .188 |
| III–IV | 34 (12.2) | 27 | 7 | 14 | 20 | 29 | 5 | |||
| Vascular emboli | ||||||||||
| Absent | 183 (65.8) | 153 | 29 | .255 | 98 | 85 | .012* | 148 | 35 | .056 |
| Present | 95 (34.2) | 73 | 20 | 35 | 58 | 64 | 29 | |||
| Margin | ||||||||||
| Negative | 263 (94.6) | 214 | 46 | .821 | 125 | 136 | .683 | 199 | 64 | .727 |
| Positive | 15 (5.4) | 12 | 3 | 8 | 7 | 13 | 2 | |||
| Tumor necrosis | ||||||||||
| Absent | 158 (61.7) | 131 | 25 | .261 | 80 | 77 | .133 | 113 | 45 | .211 |
| Present | 98 (38.3) | 76 | 21 | 40 | 57 | 77 | 21 | |||
![]()
Analyses of overall and disease-free survival
Univariate analyses of overall survival rate using the Kaplan-Meier method showed that high expression in both SIRT7 and H3K18ac were associated with poor overall survival rate (p=.034 and p=.001, respectively) (Fig. 2A, B); however, high expression in ELK4 was associated with good overall survival (p=.021) (Fig. 2C). High AJCC stage (p<.001) (Fig. 2D), large tumor size (≥5 cm) (p=.001) (Fig. 2E), and the presence of vascular emboli (p=.003) (Fig. 2F) were associated with poor overall survival rate. Additionally, younger age (<65 years) (p=.028) and positive surgical margin (p=.001) were correlated with poor outcomes. Analyses using Cox’s proportional hazards are shown in Table 2.
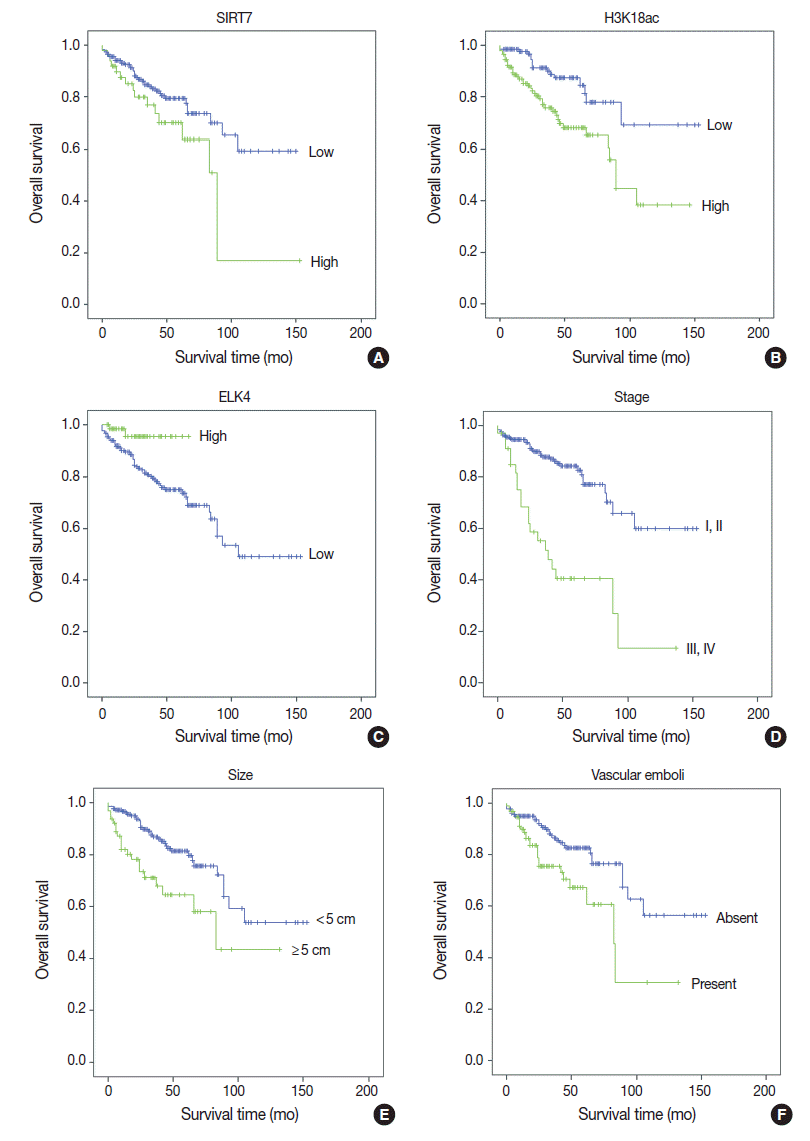 | Fig. 2.Kaplan-Meier survival curves of immunohistochemical (IHC) markers and clinicopathologic data. With IHC markers (A–C), high SIRT7 expression (A) and high H3K18ac expression (B) were associated with poor prognosis (p = .034 and p = .001, respectively). However, high ETS-like transcription factor 4 (ELK4) expression was associated with good prognosis (p = .021) (C). With clinicopathologic factors (D–F), high American Joint Committee on Cancer stage (III, IV) (p < .001) (D), large tumor size (maximal diameter ≥ 5 cm) (p = .001) (E), and the presence of vascular emboli (F) were associated with poor overall survival (p = .003).
|
Table 2.
Univariate and multivariate analyses of overall survival using Cox’s proportional hazards model
| Variable |
Univariate analysis |
Multivariate analysis |
||
|---|---|---|---|---|
| HR (95% CI) | p-value | HR (95% CI) | p-value | |
| High SIRT7 expression | 1.853 (1.036–3.313) | .038* | 1.323 (0.720–2.430) | .368 |
| High H3K18Ac expression | 2.569 (1.438–4.592) | .001* | 2.731 (1.469–5.076) | .001* |
| High ELK4 expression | 0.219 (0.053–0.907) | .036* | 0.162 (0.037–0.704) | .015* |
| Tumor size (≥ 5 cm) | 2.427 (1.400–4.207) | .002* | 2.044 (1.104–3.787) | .023* |
| Vascular emboli | 2.176 (1.273–3.720) | .004* | 1.287 (0.707–2.343) | .409 |
| Margin positive | 3.371 (1.582–7.183) | .002* | 2.943 (1.300–6.661) | .010* |
| AJCC stage III–IV | 4.302 (2.489–7.438) | < .001* | 3.007 (1.650–5.483) | < .001* |
![]()
In multivariate analyses (Table 2), high expression in H3K18ac was determined to be a poor prognostic factor (p=.001), whereas high expression in ELK4 was indicated as a good prognostic factor (p=.015). However, high SIRT7 expression was unremarkable (p=.368). Tumor size ≥5 cm, positive margin, and advanced stage (III–IV) (p=.024, p=.012, and p<.001, respectively) were related independently as poor prognostic factors. Finally, the presence of vascular emboli was not significant (p=.409).
Disease-free survival rate analyses were performed using the Cox’s proportional hazard model. High SIRT7 expression, presence of vascular emboli, AJCC stage III-IV and Edmondson-Steiner grade III–IV were associated with worse prognosis (Table 3). However, only AJCC stage was shown to be an independent factor in the multivariate analysis (p=.021).
Table 3.
Univariate and multivariate analysis using Cox’s proportional hazards analysis of disease-free survival
| Variable |
Univariate analysis |
Multivariate analysis |
||
|---|---|---|---|---|
| HR (95% CI) | p-value | HR (95% CI) | p-value | |
| High SIRT7 expression | 1.573 (1.013–2.444) | .044* | 1.440 (0.912–2.276) | .118 |
| High H3K18Ac expression | 1.229 (0.844–1.789) | .283 | 1.022 (0.680–1.537) | .915 |
| High ELK4 expression | 0.922 (0.566–1.502) | .744 | 0.901 (0.545–1.490) | .685 |
| Tumor size (≥ 5 cm) | 1.393 (0.903–2.150) | .134 | 1.110 (0.697–1.769) | .660 |
| Vascular emboli | 1.465 (1.001–2.144) | .049* | 1.192 (0.790–1.800) | .402 |
| Margin positive | 1.200 (0.558–2.580) | .640 | 0.903 (0.403–2.025) | .805 |
| AJCC stage III–IV | 2.128 (1.310–3.458) | .002* | 1.898 (1.101–3.273) | .021* |
| Edmondson-steiner’s grade | 1.513 (1.040–2.202) | .030* | 1.270 (0.839–1.923) | .258 |
![]()
Analyses of correlation
SIRT7 and H3K18ac showed a positive correlation (p=.006) (Table 4); however, ELK4 did not show a significant correlation with the other two markers. Tumor size showed a positive association with AJCC stage (p<.001), vascular tumor emboli (p=.001), and necrosis (p<.001). Additionally, tumor necrosis was positively associated with presence of vascular emboli (p=.012) as well as AJCC stage III and IV (p=.005). Edmondson-Steiner grade III and IV showed positive correlation with the existence of vascular emboli (p=.000) and high AJCC stage (p=.010). Elder patients (≥65 years) had positive association with positive surgical margin (p=.029).
Table 1 summarizes the correlation between IHC markers and clinicopathologic data. Edmondson-Steiner grade III and IV had a correlation with high SIRT7 and high H3K18ac expression (p=.002 and p<.001, respectively). A high T stage or presence of vascular emboli were associated with high H3K18ac expression (p=.027 and p=.012, respectively). In addition, high H3K18ac expression showed a significant correlation with females (p=.033) and high ELK4 expression with males (p=.021).
Go to :
DISCUSSION
The objective of the present study was to analyze H3K18ac, which is deacetylated by SIRT7 linked with ELK4, by IHC expression in HCC samples. We hypothesized that high IHC expression of SIRT7 would be associated with high ELK4 and low H3K18ac expression, and these results would lead to a poor overall survival rate. On the contrary to the hypothesis, this study found that SIRT7 expression had a weak positive correlation with H3K18ac, but no correlation with ELK4 (Table 4). In addition, high H3K18ac expression was unexpectedly associated with poor prognosis, whereas high ELK4 expression was associated with better prognosis in the overall survival analyses (Table 2). Lastly, high SIRT7 expression associated with poor prognosis was in accordance with our major hypothesis.
There is no definite explanation of the unexpected positive correlation of SIRT7 and H3K18ac, but there are several possible mechanisms. First, other pathways may exist. For example, SIRT2 and SIRT6 can also deacetylate H3K18ac in addition to SIRT7 [18,19]. Conversely, SIRT7 also deacetyl-ates p53 in addition to H3K18ac [20]. Although the prognostic direction of H3K18ac was unexpected, it seems to have significance, which is shown in the univariate and multivariate overall survival analyses. In addition, correlation analyses revealed a positive association with poor Edmondson-Steiner grade, high T stage and presence of vascular emboli (Tables 1, 2). Putting these evidences together, we can suggest that there are other important mechanisms beyond the deacetylation of H3K18ac by SIRT7. The weak correlation coefficient value may be a reflection of this (Table 4).
Second, the method of approach was different. In previous studies, experiments were performed with cell-lines and were focused on the specific gene foci [12,14]. In the present study, we used surgical specimens and examined overall protein expressions by the IHC approach. Some authors have argued that gene-specific expression of histone acetylation can run contrary to the overall state of modification. This phenomenon could have resulted from compensation and made a difference in the overall expression of acetylation seen in the IHC study [21-23]. Further evaluation of the relationship of SIRT7 and H3K18ac in HCC is needed [19]. Because previous HCC studies of H3K18ac were focused on tumor suppressor genes, the roles of H3K18ac in proto-oncogene or global H3K18ac expression in HCC progression have not been sufficiently investigated. However, several studies have shown the correlation of H3K18ac with poor prognosis in other malignancies, such as pancreatic adenocarcinoma or esophageal cancer [23,24]. Moreover, in some malignant tumors, especially in HCCs, global acetylation of H3K18 could be more associated with genes related to tumor progression or proliferation than tumor suppression [25-27].
High SIRT7 expression in this study was associated with poor overall survival rate and high tumor grade but did not show a statistical correlation with H3K18ac and ELK4 expression. Most studies on SIRT7 in HCC have focused on the HDAC activity of SIRT7 in relation to tumor suppressor genes [12,14]. Recent studies have revealed other mechanisms of SIRT7 in human cells. They have reported that SIRT7 can regulate protein folding in mitochondria, and a decrease in SIRT7 could result in stem cell senescence [28]. Another study also reported that SIRT7 can interact with SIRT1, resulting in epithelial-to-mesenchymal transition via control of E-cadherin. The mechanism of SIRT7 as a poor prognostic factor in HCC may be more related to other roles in tumor cells than its role as a suppressor of tumor suppressor genes [29].
High ELK4 expression did not show a correlation with SIRT7 or H3K18ac in the present study. However, high ELK4 expression was an independent factor of good prognosis in HCC patients. The most studied mechanism of ELK4 in tumorigenesis is the role as a cofactor of SIRT7 in deacetylation of H3K18, which is located in tumor suppressor genes or ribosome-related genes. Additionally, ELK4 fundamentally forms complexes with SRF dimers on SREs found in the c-fos and other immediate early gene promoters [15], and this process is stimulated by the activated mitogen-activated protein kinase (MAPK) signaling pathway [15,30]. Therefore, we can assume that high ELK4 status would induce the oncogenic pathway, but this has not been investigated. Furthermore, ELK4 is expected to inhibit gene transcription, due to its resemblance to ELK3/NET. ELK3/NET acts as a transcription repressor of target promoters through its NET inhibitory domain (NID) and C-terminal-binding protein inhibition domain in the absence of the MAPK signal [31,32]. Thus, loss of ELK3 expression has been reported in malignant mesothelioma, suggesting the role of ELK3 as a repressor of tumors [33]. ELK4 also has a similar NID region to ELK3/NET [34,35], suggesting that ELK4 may also inhibit gene transcription during the inactivated MAPK condition [36]. The role of ELK4 in HCC requires more investigation, but this transcriptional repression could be associated with inhibition of tumor progression.
In conclusion, high expression of ELK4 and low expression of H3K18ac were independent and good prognostic factors in HCC patients. High SIRT7 expression in HCC was associated with poor overall survival rate, although it was not an independent prognostic factor. In HCC tissue samples, these proteins had significant prognostic values independently; however, we did not find an integrated tumorigenic effect. Since these proteins have not been widely investigated, more comprehensive studies for interactions and mechanisms of ELK4, SIRT7, and H3K18ac are needed in HCC.
Go to :
 XML Download
XML Download