

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors participate in the development and maintenance of central sensitization following tissue injury or chronic inflammation [1]. AMPA receptors are ionotropic transmembrane heterotetrameric cation channels consisting of four subunits: GluA1–4. The GluA1 and GluA2 subunits are mainly located on the laminae I and II of the spinal dorsal horn, a primary location for nociception and central sensitization [2,3]. Peripheral inflammation can upregulate the AMPA receptors within the post-synaptic membranes [4]. The upregulation and trafficking of the GluA1 subunit increase excitatory postsynaptic current (EPSC) within the spinal dorsal horn. An increase in EPSC means an increase of nociception. Therefore, investigating the activity of AMPA receptors is crucial for understanding the rapid, activity-dependent interaction of the post-synaptic neurons during peripheral inflammation.
Glial cells interact with the neurons of the dorsal horn via a complex communication network [5]. The activation of nociceptors following the onset of inflammatory pain results in glial response. Activated glial cells release cytokines and chemokines, which trigger the transmission of pain to the post-synaptic neurons in the brain [6]. Astrocytes are in close contact with the neurons, and the positive feedback loop during nociception can cause direct alteration of neuronal communication [6]. Simultaneously, activated glial cells promote the release of anti-inflammatory factors that facilitate the clearance of apoptotic cells and the subsequent halting of pro-inflammatory reactions [7]. Glial cells are regarded as a therapeutic target in the treatment of chronic neuropathic pain. However, temporal changes in glial cell activity and function after disc herniation have not been identified.
Nucleus pulposus (NP) is immunogenic; thus, herniated NP may release a substance capable of inducing an autoimmune response, thereby generating chronic inflammation around the dorsal root ganglia (DRG) [8]. The rat model of lumbar disc pain can be used to induce inflammation around the spinal nerve root and DRG, following the implantation of NP harvested from the tail [9-12]. Inflammation initiates the propagation of action potentials from the peripheral nociceptors and pain perception, which, subsequently, promotes central sensitization of nociception [4,13]. Among the biomarkers and cells involved in central sensitization, we investigated the changes in AMPA receptors, as rapidly activated ion channels responding to inflammation and glial cell activation are the key processes involved in maintaining chronic pain.
Previous research reported increased AMPA receptor activity in a pain model of lumbar disc herniation 3 days after surgery following protein immunoblotting analysis [14]. However, to the best of our knowledge, no studies have investigated the sequential activation of the AMPA receptors and glial cells following disc herniation. Therefore, we aimed to investigate the time-dependent changes associated with the activation of the AMPA receptors and glial cells in the spinal dorsal horn after the implantation of NP in the rat model of disc herniation.
Go to : 
MATERIALS AND METHODS
Animals
Eighty-three Sprague-Dawley rats (200–250 g) were randomly assigned to the control, sham-operation, and NPexposed groups (Fig. 1). For the analysis of AMPA receptor activity, the rats (n=40) were assigned to the control (n=6), sham-operation (n=4), and NP-exposed (n=30) groups. For assessing the immunohistochemical expression of OX-42 and glial fibrillary acidic protein (GFAP), the rats (n=40) were assigned to the control (n=10) and NP-exposed (n=30) groups. Moreover, three rats were used to investigate the progression of neuropathic pain. They were housed, two per cage, with diurnal light-dark cycles, and food pellets and water were provided ad libitum.
Lumbar disc herniation operation
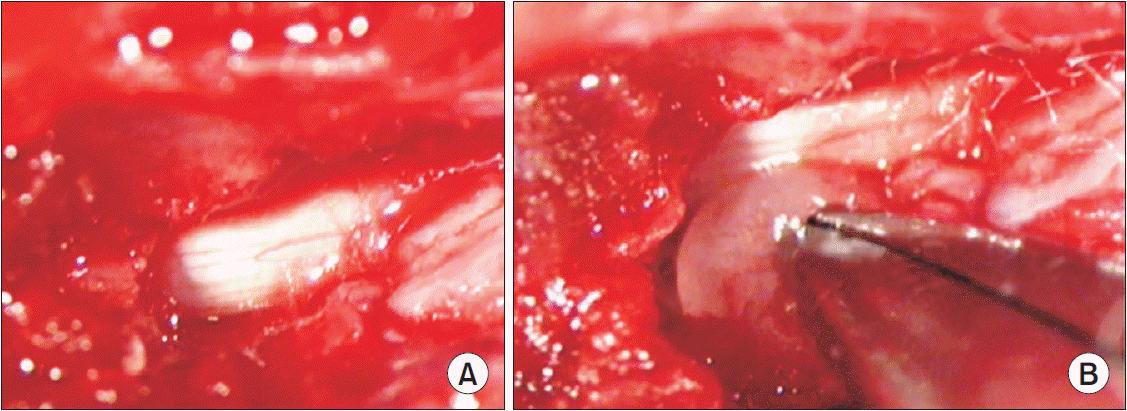
All experimental procedures were conducted according to the guidelines of the Institutional Animal Care and Use Committee at Yeungnam University, Republic of Korea (No. YUMC-2017-010). The rats were anesthetized through the intraperitoneal injection of Zoletil 50 mg/kg (Virbac, Carros, France). With the animals placed in the prone position, a midline dorsal incision was made within the pelvic area, and the multifidus muscles were separated along the L4–S1 spinous processes. Laminectomy was performed to expose the left L5 nerve roots and DRG. An incision, approximately 1 cm in length, between the second and third coccygeal vertebrae of each tail was made on the dorsal surface of the proximal tail for harvesting the autologous NP (Fig. 2). The NP was harvested, using a curette, and implanted beside the left L5 nerve root, immediately proximal to the DRG. The sham surgery was performed similarly, but without the implantation of autologous NP in the left L5 nerve root.
Pain behavior evaluation
Mechanical allodynia was determined by measuring the paw withdrawal response to the mechanical stimulation of the ipsilateral hind paws with von Frey filaments (North Coast Medical Inc., Morgan Hill, CA, USA); it was calibrated in grams. The rats were placed in a clear plastic cage with a metal mesh floor and permitted to adapt to the testing environment for 15 minutes, and the plantar surface of each hind paw was stimulated to cause a slight bending of the von Frey filaments for 3–5 seconds. The filaments were stimulated in the order of increasing and decreasing thicknesses, beginning with a 0.1 g probe, and the stimulation was maintained until a consistent withdrawal response to >3 out of 5 stimuli was observed. If no withdrawal response was elicited by the 15 g filament, the mechanical threshold was set at 15 g. The mechanical allodynia at the plantar surfaces of the ipsilateral hind paws was assessed three times a week until the animals were sacrificed. Three rats were tested for up to 38 days postoperatively to identify the pattern of progression of neuropathic pain (Fig. 3).
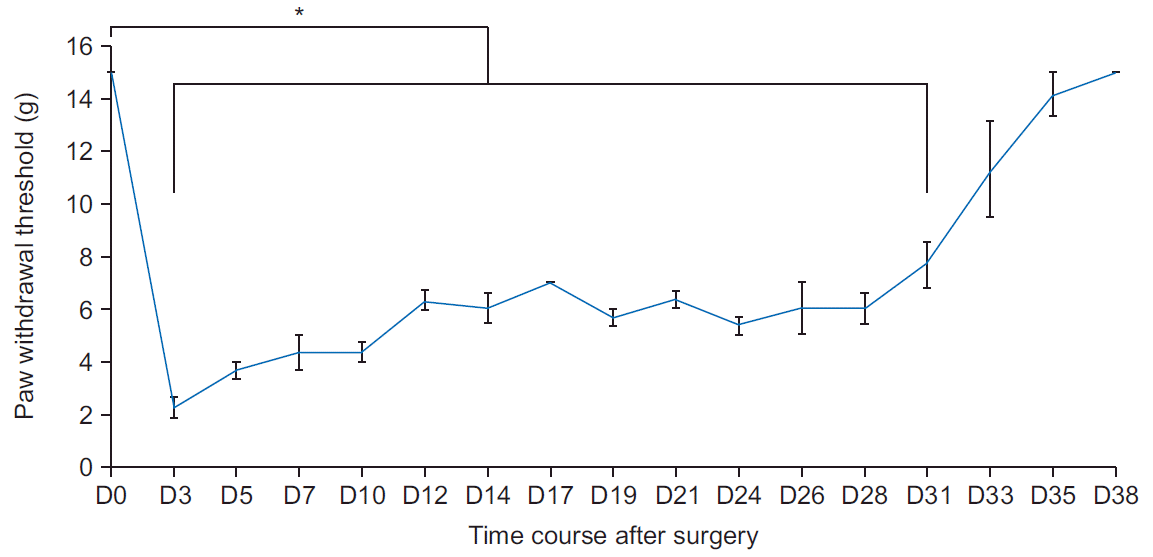 | Fig. 3.Time course of neuropathic pain behaviors. Hyperalgesia in a rat model of lumbar disc herniation induced by implantation of the nucleus pulposus to the left L5 nerve root. The withdrawal threshold in the ipsilateral paw is noted at 3 to 38 days following surgery. D, postoperative day. *p<0.05 versus the baseline.
|
Immunohistochemical examination
Five rats each were euthanized 3, 7, and 14 days after surgery for the immunohistochemical staining of the spinal cord for the GluA1 and GluA2 subunits of the AMPA receptor OX-42 and GFAP. The rats in the control group were euthanized without performing sham surgery. Under anesthesia, a catheter was inserted from the left ventricle of the heart to the aorta, and the animals were perfused with heparinized saline followed by perfusion with 500 mL of 4% paraformaldehyde in phosphate buffer (0.1 M, pH 7.4). The L5 spinal cord segment was excised and fixed for 24 hours with the perfused fixative, and the tissue samples were cryoprotected in 30% sucrose made in phosphate buffer. Transverse sections of the spinal cord (30-μm thick) were sliced on a cryostat and blocked with 10% normal donkey serum (Jackson ImmunoResearch Laboratories Inc., West Grove, PA, USA) in 0.01 M phosphate-buffered saline (PBS) containing 1% Triton X-100 for 40 minutes to decrease the non-specific binding of the primary antibody. For performing immunofluorescence microscopy, the sections were incubated overnight at room temperature with a mixture of AntiGluR1 (ABN241, dilution 1:1000; Millipore, Burlington, MA, USA) and anti-GluR2 (AB1768-I, dilution 1:2000; Millipore) antibodies, or with mouse anti-OX-42 (dilution 1:200; Chemicon, Temecula, CA, USA) and anti-GFAP (dilution 1:1000; Dako, Santa Clara, CA, USA) antibodies. After primary antibody incubation, the sections were rinsed with PBS 3 times (5 min/rinse) and incubated with 2% normal donkey serum for 15 minutes followed by an appropriate combination of Alexa 488-conjugated donkey anti-rabbit secondary antibodies (dilution 1:200; Invitrogen, Eugene, OR, USA) containing 1:200 dilution of Triton X-100 for 2 hours. After several rinses postincubation, the sections were mounted on coverslips covered with a VECTASHIELD Mounting Medium (Vector Laboratories, Burlingame, CA, USA) and observed under a fluorescence microscope (Digital Module R, Leica Microsystems, Switzerland). Images were visualized and captured by a charge-coupled device camera (Olympus, Tokyo, Japan) attached to the microscope.
Quantitative image analysis
To analyze the immunopositivity of AMPA receptors and glial cells in the spinal dorsal horn, images obtained from the L5 spinal cord sections were captured. One image (898×660 μm) captured by the camera was encoded to blind the investigator before analysis. The number of pixels positive for AMPA receptors and glial cells was segmented by applying an appropriate threshold. The intensity of green fluorescence for each frame was obtained using an image analysis software (Leica application suite V4.2; Leica Microsystems, Switzerland). The superficial dorsal horn of the spinal cord had a connection with nociceptive neurons [15]. AMPA receptors are more in the superficial dorsal horn of the spinal cord [16]. AMPA positive and glial cell activity in the superficial dorsal horn were evaluated [17,18]. The mean values of each group and the time point versus control mean value in the dorsal horn of the L5 spinal level were calculated as percentages. The numbers of OX-42-positive and GFAP-positive cells were counted using the image analysis software.
Statistical analysis
Data from each rat were pooled and processed using Excel version 16.26 (Microsoft Corporation, Redmond, WA, USA). The collected data were uploaded into SPSS software (IBM Corporation, Armonk, NY, USA) and analyzed. For comparison, the control values were considered as 100. Comparisons of the findings from the quantitative image analysis of immunoreactivities and the measurements of mechanical withdrawal thresholds were performed using the Kruskal-Wallis test. Statistical significance was set at p<0.05.
Go to :
RESULTS
Pain behavior
The mechanical withdrawal threshold of the control and sham-operated groups was 15 g. Rats that underwent sham surgery did not exhibit a decreased mechanical withdrawal threshold 3 days after surgery. Rats in the NP-exposed group that underwent lumbar disc herniation surgery showed statistically significant reductions in the mechanical withdrawal thresholds of the ipsilateral hind paw 3 days after surgery, and these were maintained until postoperative day 31 (Fig. 2).
GluA1 and GluA2 expression
The immunoreactivities of GluA1 and GluA2 were well-visualized in the spinal dorsal horn, and they showed intense staining in the superficial lamina of the dorsal horn, the region that received nociceptive input from the periphery (Fig. 4). Compared with the control group, the immunoreactivity of GluA1 significantly increased in the animals that underwent surgery within postoperative days 3 to 14. Three and 7 days after the surgery, the increase in immunoreactivity of GluA1 in the spinal dorsal horn was statistically significant (Fig. 5). The immunoreactivity of GluA2 also increased within postoperative days 3 to 7; however, the magnitude of increase was relatively small compared with the immunoreactivity of GluA1. The differences between the immunoreactivities of GluA1 and GluA2 at each time point were not statistically significant. Fourteen days after surgery, the increased immunoreactivities of the GluA1 and GluA2 subunits observed in the rats that underwent surgery gradually normalized to levels observed in homeostatic conditions similar to those of the control group.
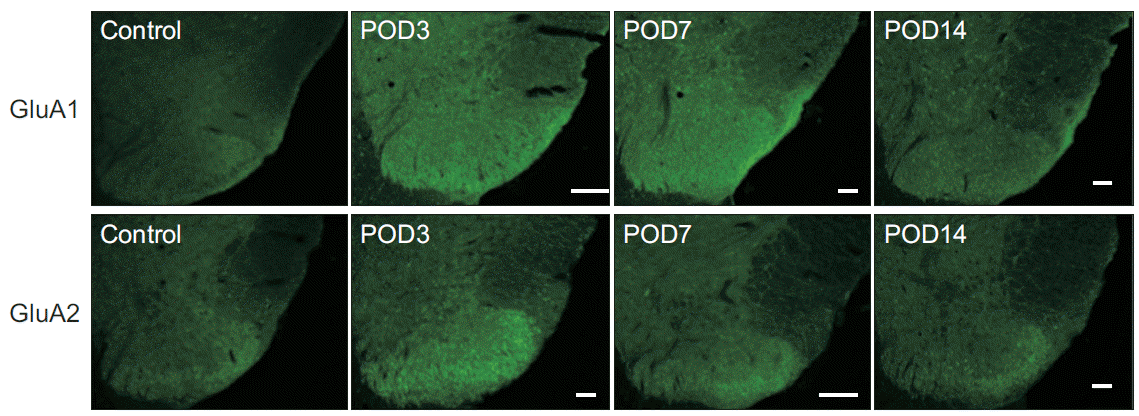 | Fig. 4.Immunohistochemical expression of GluA1 and GluA2 in the dorsal horn of the L5-level spinal cord before (control), 3, 7, and 14 days after surgery. GluA1 are GluA2 are intensely stained in the superficial lamina of the dorsal horn. POD, postoperative day.
|
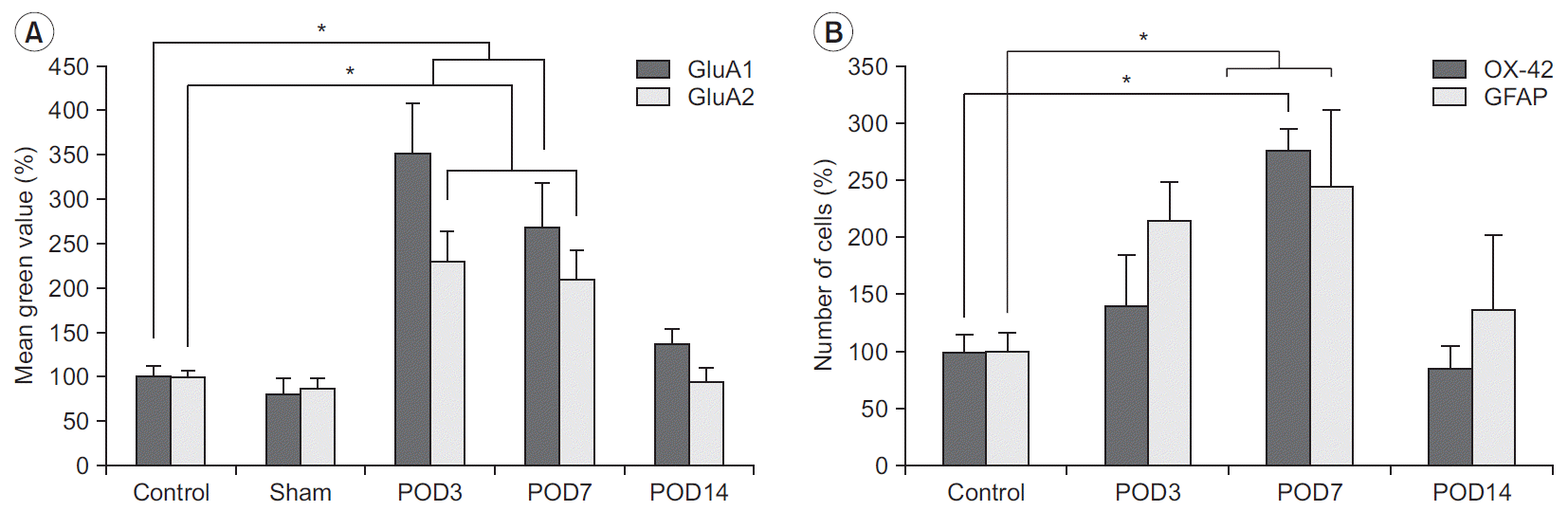 | Fig. 5.Semi-quantitative analysis of immunoreactivity of AMPA receptors and microglial cells in the dorsal horn of the spinal cord at postoperative day (POD) 3, 7, and 14. (A) Mean green value of GluA1 and GluA2 is significantly higher at POD3 and POD7, compared with control. (B) Immunoreactivity of OX-42 and GFAP are also increased and return to the control value at 14 days after surgery, with a pattern similar to that of AMPA receptors. For quantification, values and numbers were counted and averaged. Values are presented as mean±standard error. AMPA, alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; GFAP, glial fibrillary acidic protein. *p<0.05 versus the control group.
|
OX-42 and GFAP
Immunohistochemical examination of the spinal dorsal horn for OX-42 and GFAP demonstrated reactive activation of the microglia and astrocytes after surgery (Fig. 6). The immunoreactivity of OX-42 within the superficial lamina of the spinal dorsal horn had significantly increased on postoperative day 7. The immunoreactivity of GFAP also significantly increased on postoperative days 3 and 7 (Fig. 5). On postoperative day 14, the numbers of OX-42-positive and GFAP-positive cells decreased to levels observed in the control group.
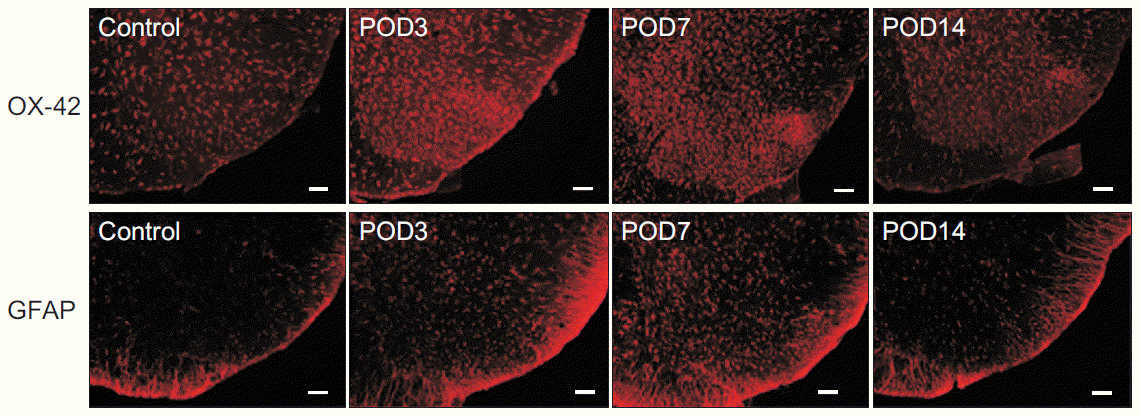 | Fig. 6.Immunohistochemical expression of OX-42 and glial fibrillary acidic protein (GFAP) in the dorsal horn of the L5-level spinal cord before (control), 3, 7, and 14 days after surgery. Increased OX-42-positive microglia are noted in superficial ipsilateral dorsal horns at 3 and 7 days following surgery compared with control. GFAP-positive astrocytes are also diffusely elevated in the spinal dorsal horn following surgery. POD, postoperative day.
|
Go to :
DISCUSSION
NP is immunogenic; a herniated NP may release substances that induce an autoimmune response and generate chronic inflammation. Inflammation stimulates the nociceptive nerves, which causes nociceptive pain following disc herniation. Moreover, the inflammation of an injured DRG or nerve root promotes the initiation and maintenance of neuropathic pain. Based on this concept, an epidural steroid injection following disc herniation has been accepted as a logical and effective therapeutic option [19]. However, the role and involvement of inflammation throughout the entire process of pain—from the beginning of disc herniation to the chronic neuropathic pain following disc herniation—have not been fully elucidated. Establishing a treatment method for radicular pain will facilitate a better understanding of the role of inflammation in nociception, from early nociceptive to and chronic neuropathic pain. Therefore, we investigated the activity of AMPA receptors reciprocally activated by peripheral inflammation and glial cells involved in maintaining chronic neuropathic pain.
Glutamate is an essential excitatory neurotransmitter in the central nervous system. Functional AMPA receptors have been observed in various brain regions and the spinal cord. AMPA receptors are ionotropic glutamate receptors in the post-synaptic membrane that dynamically respond to nociceptive input from peripheral inflammation and contribute to the maintenance of long-lasting transmission of somatosensory input [20,21]. AMPA receptor phosphorylation and upregulation of AMPA increase permeability to Na+ and Ca2+, which increases EPSCs [4,22]. The upregulation of AMPA receptors is caused by the translocation of AMPA receptors from the cytosol to the plasma membrane. Long-term potentiation (LTP) describes the long-lasting strengthening of synapses based on recent patterns of activity. The AMPA receptor is the main participant in the formation of LTP.
When it comes to the pain, LTP refers to changes in nociception such as hyperalgesia. The continuous increase in EPSC, which is a mechanism of LTP, stimulates central sensitization as in chronic neuropathic pain [4]. AMPA receptors are expressed by neurons and glial cells [23,24]. Classically, astrocytes are characterized by high glutamate transporter activity [25]. Considering that glial cells play an essential role in chronic pain, AMPA receptors play a role in the development of central sensitization of the neurons and glial cells within the dorsal horn of the spinal cord.
Although many cells and complex processes result in central sensitization following an insult, we have focused on the AMPA receptor as a surrogate of peripheral inflammation to investigate the duration of inflammation in the process of pain and central sensitization. The rat pain model of lumbar disc herniation showed an increased total level of the AMPA receptor within the dorsal horn. This is a remarkable finding, given those observed for other chronic pain models. Autoimmunity and hypoxia caused by the herniated NP result in inflammation around the herniated disc. Hence, the investigation of the sequential changes in the AMPA receptor levels can facilitate a better understanding of the temporal correlation between the increased EPSCs caused by inflammation and disc herniation.
Contrary to other pain models, a significant increase in the GluA1 and GluA2 subunits of AMPA receptors was observed in the rat pain model of lumbar disc herniation in this study. These increases in immunoreactivity persisted for 3 to 7 days after surgery and returned to baseline 14 days after surgery. The paw withdrawal threshold was reduced after surgery and maintained for 31 days after surgery. AMPA involvement is the highest at the early phase of neuropathic pain, and its role gradually diminishes in chronic neuropathic pain. The AMPA receptor mainly mediates fast EPSCs in dorsal horn neurons. Therefore, peripheral inflammation promotes the upregulation of AMPA receptors, which strengthens the synapses among afferent and second-order nociceptive nerves, interneurons within the dorsal horn, and glial cells. Despite the normalization of GluA1 and GluA2 14 days after surgery, which indicates a decrease in nociceptive input from inflammation, central sensitization facilitated the maintenance of mechanical hypersensitivity after surgery.
Continuous nociceptive input followed by a rise in the AMPA receptor activities increases the action potential of post-synaptic neurons, which promotes central sensitization. Subsequently, central sensitization yields chronic pain, even if there is no further increase in the activity of the AMPA receptors. This activity gradually reduces. Therefore, AMPA receptors play an important role during the period of increased EPSC due to inflammation following disc herniation.
Chen et al. [26] reported that GluA2 protein levels were significantly low in the plasma membrane in the rat model of nerve injury. Moreover, cytosolic GluR2 levels were elevated, and the amount of membrane-bound GluA2 was significantly reduced after 24 hours postFreund’s complete adjuvant (CFA) injection [27]. GluA2 is a Ca2+-impermeable ion channel. GluA2-lacking AMPA receptors in the dorsal horn induce an inward rectification of EPSCs, thereby maintaining chronic neuropathic pain. N-methyl-D-aspartate receptors (NMDARs) are critically involved in the induction and maintenance of persistent pain after a noxious event [28]. The activated NMDAR/triggered protein kinase C signaling cascade induces GluR2 internalization in dorsal horn neurons after persistent inflammation [3]. Therefore, the reduction in GluA2 subunits in the plasma membrane after nerve injury or CFA injection constitutes an important mechanism of pain. However, in this study, the immunoreactivity of GluA2 was elevated, likewise the immunoreactivity of GluA1. Although not statistically significant, the magnitude of increase in the immunoreactivity of GluA1 was greater than that of GluA2 in this study; therefore, the GluA1/GluA2 ratio was elevated, as reported by a previous study [17].
The hind paw is injected with inflammatory agents to create rat models of inflammatory hyperalgesia. The GluR1 and GluR2 trafficking patterns reportedly vary based on the inflammatory agents injected [29]. Most studies reported no definite change in the total amount within the dorsal horn, except their increased insertions into the membrane. However, these studies have investigated changes in the distribution of GluA1 within several minutes or up to 1 day after an insult. Our study revealed the upregulation of GluA1 and GluA2 after 7 days after surgery. Therefore, we speculate that this difference is due to the extended period of inflammation and the location of the inflammation.
Excitatory neurotransmitters released by presynaptic neurons within the dorsal horn activate glial cells. The activation of the Toll-like receptor 4 (TLR4) and P2X4 receptors has been implicated in chronic pain states following peripheral nerve injury [5]. Activated glial cells closely communicate with neurons. Tumor necrosis factor-α (TNF-α), interleukin-1β, and brain-derived neurotrophic factor are released from activated glial cells, and this initiates hyperactivity in dorsal horn cells [6]. Conversely, cellular debris from apoptotic and necrotic cells promote phagocytosis of glial cells, resulting in a decrease in pro-inflammatory cytokines [30]. Therefore, activated glial cells play a role in promoting neuropathic pain and have a protective mechanism at the same time.
We found an increase in astrocytes and microglia in the dorsal horn 3 and 7 days following surgery, which returned to control levels on day 14. Therefore, the activity of glial cells also increased in response to inflammation induced by the implanted NP followed by a gradual decrease. Excessive AMPA receptor activation in astrocytes causes the downregulation of astrocyte GLAST (glutamate aspartate transporter) [31]. GLAST removes glutamate from the extracellular space. The downregulation of astrocyte GLAST aggravates excitotoxic conditions by increasing extracellular glutamate levels. The increased AMPA receptor activation of microglia releases inflammatory mediators and TNF-α and reduces protective mediators [32]. TNF-α produced by microglia stimulates the upregulation of transmembrane AMPA receptors on neurons, which increases their reaction to extracellular glutamate. Therefore, the upregulation of AMPA receptors on glial cells leads to an increase in extracellular glutamate, which has a role in initiating central sensitization.
Our study has several limitations. First, the activation of AMPA receptors is not a direct and precise surrogate of peripheral inflammation following disc herniation. However, the degrees of activity of various cytokines and immune cells during inflammation are modified depending on the time. Moreover, we aimed to investigate the changes in the central, and not the peripheral, nervous system. Hence, we selected AMPA receptors to indicate the activity of the central nervous system in response to peripheral inflammation following the surgery. Second, we conducted immunohistochemistry only for measuring AMPA receptor levels. Hence the mechanisms underlying an increase in AMPA receptors and those underlying its increase in production or decrease in degradation need to be elucidated.
In summary, in the rat pain model of lumbar disc herniation, increased immunoreactivity of AMPA receptors (GluA1 and GluA2 subunits) and glial cells (microglia and astrocyte) was observed on days 3 and 7 after surgery, and they decreased to control levels after 14 days. Therefore, AMPA receptor activation and gliosis caused by inflammation after disc herniation play roles in the induction of central sensitization.
Go to :
 XML Download
XML Download