

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Neuropathic pain is defined as pain caused by a lesion or disease of the somatosensory nervous system [1]. Neuropathic pain is one of the chronic intractable pain syndromes. If pain is not controlled at an early stage, it may progress to refractory pain and limit activities of daily living independence [23]. Existing pharmacologic treatments for neuropathic pain are limited, with partial relief of pain in 40%-60% of patients [2]. Therefore, there is growing attention to non-pharmacologic managements for neuropathic pain.
Radiofrequency (RF) has long been used for the treatment of neuropathic pain [45]. RF treatment is classified as conventional RF (CRF) and pulsed RF (PRF) [6]. CRF uses frictional heat generated from the catheter needle. A wide range of thermal damage due to RF stimulation can cause nerve damage. Unlike CRF, PRF has the advantages of safe application and fewer side effects, because application sites maintain the temperature below 42℃ [78]. Biological effects of PRF differ from CRF, but action mechanism of PRF is still unknown. One of the current hypotheses is neuromodulation through gene expression change [9]. Tumor necrosis factor-α (TNF-α) as a pain modulating factor, is involved in the development of neuropathic pain after peripheral nerve injury [101112]. TNF-α protein is increased in injured peripheral nerve and dorsal root ganglion (DRG) and increased immunoreactivity is detected in glial cell and nerve cell body after chronic constriction injury (CCI)-induced neuropathic pain [13]. Our hypothesis was that PRF applied proximal to the injured peripheral nerve would decrease neuropathic pain symptoms, which affect the level of TNF-α in the peripheral nerve and spinal cord.
While RF to DRG requires fluoroscopic guidance, approach to the peripheral nerve is relatively easy with recent prevalence of musculoskeletal ultrasonography. If PRF applied to the peripheral nerve is effective for pain control, it could be developed as an easy and comfortable therapeutic tool for neuropathic pain patients. Recent human studies showed positive results [1415]. Animal studies are helpful in understanding the therapeutic effects and the action mechanisms of various treatments, however, animal studies on the application of PRF to the injured peripheral nerve are rare. We aimed to investigate the effect of PRF applied to sciatic nerve on neuropathic pain in CCI rat model.
Go to : 
MATERIALS AND METHODS
Subjects
Nineteen male Sprague-Dawley rats, weighing 250-300 grams, aged 10 weeks old, were used in the study. The rats were housed at a temperature of 22℃±2℃ and a humidity of 55% with a 12-hour light/dark cycle (light on 8 AM to 8 PM) with free access to food and water ad libitum.
All rats underwent CCI procedure as described by Bennett and Xie [16]. Withdrawal frequency and withdrawal latency were measured to evaluate whether neuropathic pain was induced on the 7th day after surgery. Rats were randomly divided into two groups according to the application of PRF: group A, CCI group (n=9) and group B, CCI treated with PRF group (n=10). Group B was treated with PRF. All animal experiments were conducted in accordance with the guidelines of the Chonnam National University Animal Care and Use Committee.
Induction of neuropathic pain
Unilateral CCI of right sciatic nerve was induced in all rats [16]. Rats were anesthetized with tiletamine HCl/zolazepam HCl (Zoletil; Virbac, Carros, France) 50 mg/kg and xylazine (Rompun; Bayer Korea, Seoul, Korea) 7 mg/kg intraperitoneally. In brief, we removed the hair of right gluteal region and disinfected the skin with povidone iodine. The skin was opened over a length of 2 cm in the proximal half of the line between trochanter and knee joint. The biceps femoris muscle was longitudinally separated and the sciatic nerve was exposed under the muscle. Proximal to the trifurcation of sciatic nerve (tibial, sural and common peroneal nerves), 4 loose ligatures of 4-0 chromic catgut at an interval of 1 mm were placed around the sciatic nerve. After the induction of CCI, the skin and muscle fascia were sutured. For the prevention of infection, gentamicin sulfate (5 mg/kg) was administered into the gluteal muscle for 3 days. All surgical procedures were performed by a researcher.
Pulsed radiofrequency
All rats of group B were anesthetized intraperitoneally and the sciatic nerves with CCI were exposed again. The rats underwent PRF procedure on the sciatic nerve using a 10-cm long electrode with a diameter of 0.55 mm and exposure length of 1 mm powered by an RF generator (PMG-230; Baylis Medical Company Inc., Montreal, Canada). Probe for PRF stimulation was fixed to a stereotactic frame and 1-2 mm of the distance between probe and the sciatic nerve was placed to avoid direct mechanical stimulation [17]. The lesion generator was set to deliver RF proximal 1 mm to the ligatures in the exposed sciatic nerve [18] at 2 bursts per second of 20 ms duration each with the automatic temperature control activated and set to 42℃ for 120 seconds. PRF was applied twice at an interval of 1 minute [19].
Evaluation of neuropathic pain
Withdrawal frequency and withdrawal latency were measured before and at 12 hours and 7 days after PRF. For the evaluation of mechanical allodynia, withdrawal frequency of the hind paw was measured by determining paw withdrawal after probing the paw with a series of calibrated monofilaments, Touch Test Sensory Evaluator (North Coast Medical Inc., Gilroy, CA, USA) [20]. All rats were placed on a metal mesh and the plantar surface of the operated hind paw was touched with Von Frey monofilament with a bending force of 5.07 mN. The occurrence of withdrawal in response to innocuous mechanical stimuli was measured at a stimulation rate of 0.2 Hz in 20 trials.
For the evaluation of thermal hyperalgesia, withdrawal latency to the heat stimulus was measured by paw withdrawal from a focused beam of radiant heat, Plantar test (Ugo Basile, Varese, Italy) onto the surface of the hind paw [21]. All rats were placed in a plastic box with a 3-mm thick glass floor and allowed to acclimate to their environment before testing. A mobile radiant infrared source under the transparent glass floor was applied to the plantar surface of right hind paw. Withdrawal latency was defined as the time from the onset of the heat stimulus to the withdrawal of the hind paw. The intensity of the heat stimulus was 55. A cut-off time of 30 seconds was set to avoid possible tissue damage. Each paw was tested twice with at least 5 minutes interval between consecutive tests. The mean value of the 2 trials was used for data analysis.
Immunohistochemistry
The sampled rats of each group were sacrificed at 12 hours and 7 days after PRF. Rats were deeply anaesthetized with 2.5% pentotal sodium, and the heart was then exposed. Each rat was transcardially perfused with 100 mL saline, followed by phosphate buffer (0.1 mol/L PB, pH 7.4) of 4% paraformaldehyde 400 mL. The sciatic nerves were removed from 5 mm proximal to the lesion to 3 mm distal to the lesion and the spinal cords (2 cm long) were removed from lumbar enlargement, fixed in 10% formalin solution for 48 hours and embedded in paraffin wax. Paraffin embedding tissues of sciatic nerve and spinal cord were cut axially at 5 µm thickness. After being posted on the slide, tissues were treated with xylene and deparaffinized. The sections were dehydrated in graded alcohol. After sections were rinsed in phosphate-buffered saline (PBS, pH 7.4), nerve sections were then incubated with 3% hydrogen peroxide for 30 minutes. Sections were incubated with TNF-α protein antibody as primary antibody diluted 1:500. Incubations with the primary antibody were conducted overnight at 4℃, sections were then washed thrice with PBS and incubated with quick anti-rabbit/anti-mouse immunoglobulin for 30 minutes. Sections were subsequently washed thrice with PBS and incubated with streptavidin peroxidase for 10 minutes. The same sections were washed and reacted with 3,3'-diaminobenzidine (DAB; DAKO Corp., Carpinteria, CA, USA) for 10 minutes at room temperature and counter-stained with Mayer's hematoxylin. TNF-α expression was evaluated under a light microscope (Nikon, Tokyo, Japan) at a magnification of 200.
Western blotting
Rats were rapidly decapitated under intraperitoneal anesthesia. Sciatic nerves and spinal cords were removed and immediately placed in liquid nitrogen and stored at -70℃ until use. Rapidly frozen sections were then washed with PBS and kept until manually homogenized in liquid nitrogen to powder form; tissue homogenates were then placed in a 1.5-mL tube. PRO-PREP Protein Purification Kit (iNtRON Biotechnology, Seongnam, Korea) extraction buffer 600 µL solution was added and dispersed. Samples were agitated and centrifuged for 30 minutes at 13,000 rpm to separate supernatant 200 µL at 4℃ and transferred to a new tube. Protein was diluted and quantified with reading by ELISA using Pierce BCA Protein Assay Kit (Thermo Fisher Scientific Inc., Waltham, MA, USA). After mixing the sample buffer, NuPAGE (Invitrogen, Carlsbad, CA, USA), protein denaturation was induced by heating for 5 minutes at 38℃. Denatured proteins were separated by electrophoresis on 12% acrylamide gel and transferred to nitrocellulose membranes. The membranes were blocked in 5% skim milk/TBS-T for 1 hour at room temperature and TNF-α antibody 1:1,000, in 3% skim milk/TBS-T (Novus Biologicals, Littleton, CO, USA) was incubated overnight at 4℃. The membranes were washed in Tris-bufferd saline with 0.1% Tween-20 (TBS-T). The blots were incubated in membrane for 1 hour at room temperature with goat anti-rabbit IgG-HRP 1:5,000, in 1% skim milk/TBS-T (Santa Cruz Biotechnology, Santa Cruz, CA, USA). After washing with Tris-bufferd saline (TBS-T), immunoreactive protein bands were visualized using the enhanced chemiluminescence, Immobilon Western Chemiluminescent Kit (Merck Millipore, Billerica, MA, USA) and LAS-3000 Imaging System (Fujifilm, Tokyo, Japan). After the images of bands were scanned in grayscale at a resolution of 600 dpi, the bands were quantified using ImageJ (National Institute of Health, Bethesda, MD, USA).
Statistical analyses
Statistical analyses were performed using SPSS ver. 21 statistical software (IBM SPSS, Armonk, NY, USA). The results were expressed as mean±standard deviation. Repeated measures analysis of variance (RM-ANOVA) was used to study the differences between groups and the changes with time in behavioral tests. Post hoc testing consisted of multiple comparisons using the Bonferroni method. Mann-Whitney U-test was used to compare the TNF-α immunoreactivity between both groups. A difference was accepted as significant if p<0.05.
Go to :
RESULTS
Before PRF, there were no significant differences between group A and group B in withdrawal frequency (p>0.05) and withdrawal latency (p>0.05), respectively. Withdrawal frequency decreased over time in group B. There was significant within-subjects effect (p=0.002) and between-subjects effect (p=0.004). In pairwise comparisons, there were significant difference of withdrawal frequency between before and 12 hours after PRF (p=0.042) and between before and 7 days after PRF (p=0.026). There was significant interaction between time and group for withdrawal frequency (p<0.001) (Fig. 1). Withdrawal latency increased over time in group B. There was significant within-subjects effect (p<0.001) and between-subjects effect (p<0.001). In pairwise comparisons, there were significant difference of withdrawal latency between before and at 7 days after PRF (p=0.001). There was significant interaction between time and group for withdrawal latency (p<0.001) (Fig. 2).
 | Fig. 1The effect of pulsed radiofrequency (PRF) on mechanical allodynia. Withdrawal frequency decreases over time in group B. There is significant interaction between time and group for withdrawal frequency. There are significant differences in withdrawal frequency between before and 12 hours after PRF, and between before and 7 days after PRF. *p<0.05.
|
 | Fig. 2The effect of pulsed radiofrequency (PRF) on thermal hyperalgesia. Withdrawal latency increases over time in group B. There is significant interaction between time and group for withdrawal latency. There are significant differences in withdrawal latency between before and 7 days after PRF. *p<0.05.
|
TNF-α immunoreactivities were observed in the sciatic nerves and the neurons of spinal cord dorsal horn in both groups. Few TNF-α-positive stained cells were detected in the spinal cord at 7 days after PRF stimulation (Fig. 3). The levels of TNF-α protein expression were not significantly different between both groups at 12 hours after PRF. However, the protein expression of TNF-α was down-regulated in the spinal cord (p=0.04) and sciatic nerve (p=0.03) of group B than group A at 7 days after PRF (Fig. 4).
 | Fig. 3Tumor necrosis factor-α (TNF-α) immunoreactivities in the sciatic nerves and the neurons of spinal cord dorsal horn (×200). Few TNF-α-positive stained cells are detected in the spinal cord at 7 days after pulsed radiofrequency stimulation. Left column is group A; right column is group B. (A, B) Sciatic nerve at 12 hours. (C, D) Dorsal horn of spinal cord at 12 hours. (E, F) Sciatic nerve at 7 days. (G, H) Dorsal horn of spinal cord at 7 days.
|
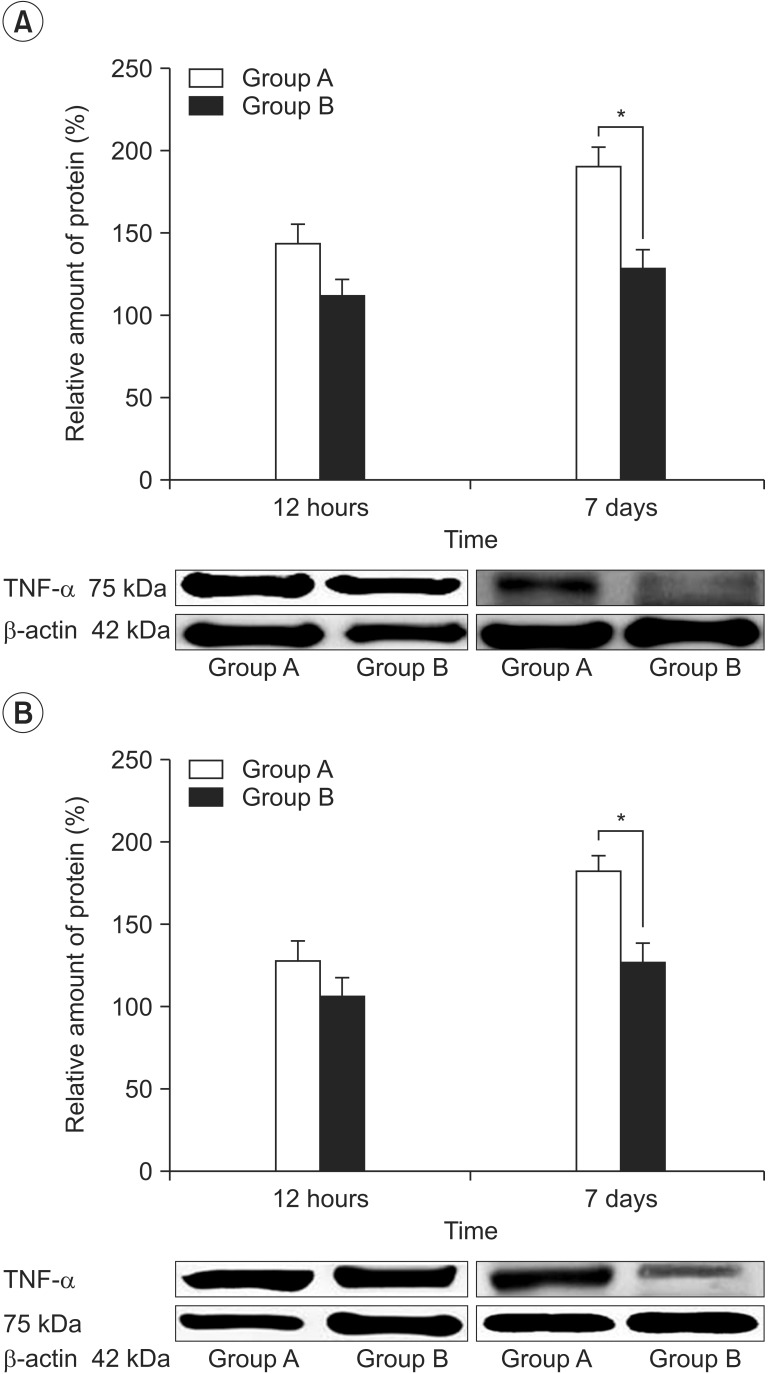 | Fig. 4The levels of tumor necrosis factor-α (TNF-α) protein expression are not significantly different between both groups at 12 hours after pulsed radiofrequency (PRF). However, the protein expression of TNF-α is down-regulated in the sciatic nerve (A) and spinal cord (B) of group B than group A at 7 days after PRF. *p<0.05.
|
Go to :
DISCUSSION
Neuropathic pain can result from peripheral nerve injuries, which trigger maladaptive alterations in the nervous system leading to peripheral and central sensitization that underlie transition to chronic pain [22]. Neuropathic pain can lead to a number of symptoms such as spontaneous pain, allodynia and hyperalgesia [1]. CCI model is a commonly used rat model for neuropathic pain [16]. In CCI model, thermal hyperalgesia occurs with loose ligation around the sciatic nerve without actual mechanical damage [23]. Thus, the change in inflammatory microenvironment rather than nerve injury per se is important for the development of neuropathic pain [2425]. TNF-α is a proinflammatory cytokine produced by peripheral nerve glia [26]. Local sciatic injection of TNF-α caused significant thermal hyperalgesia and mechanical allodynia for 3 days post-injection in association with nerve edema, splitting of myelin lamellae with vacuolization, Schwann cell injury, fibroblast and macrophage activation, and phagocytosis of lipid debris. TNF-α is implicated in neuropathic pain pathologies [27]. TNF-α is associated with the development of neuropathic pain in various animal models. CCI led to the significant development of neuropathic pain symptoms at 14 days after surgery [28]. CCI of the sciatic nerve induces TNF-α expression in the sciatic nerve [29]. In addition, expression of TNF-α and generation of tactile allodynia/hyperalgesia are correlated [30]. TNF-α appears early in the cytokine cascade. TNF-α protein expression displayed 2 distinct peaks (6 hours and 5 days) in its temporal profile after CCI [31]. TNF-α produced at the nerve injury site is axonally transported to DRG neurons and the dorsal horn of spinal cord after CCI of the rat sciatic nerve [31]. In this study, CCI group without PRF treatments displayed significant neuropathic pain symptoms and elevated TNF-α level in the sciatic nerve and the spinal cord. Elevated levels of TNF-α in the sciatic nerve at 14 days after CCI are consistent with other studies [28]. Increased levels of TNF-α in sciatic nerve and spinal cord in CCI rats concomitant with the development of mechanical and thermal hyperalgesia clearly indicate a role for cytokines in neuropathic pain [32].
RF treatment is increasingly used in the management of neuropathic pain. CRF is a neuro-destructive procedure using heat. It slows nerve regeneration by denaturation of protein, which occurs at >60℃ and nonspecific destruction of nervous continuity, leading to long-duration treatment effect than other nerve blocks [33]. PRF is known to have analgesic effect by mechanisms other than heat transfer [7]. Exposure of DRG to PRF induces c-Fos expression in lamina I and II of the dorsal horn, unlike CRF exposure, suggestive of an early cellular response to the electric/dielectric forces generated within the DRG rather than a thermal effect [9]. Because PRF does not produce tissue injury around catheter needle by excessive heat, there is no danger of deafferentation pain [34]. Application of PRF to sciatic nerve and DRG causes transient changes that do not progress to overt axonal injury [19]. PRF resulted in subclinical changes including endoneurial edema caused by alterations in the function of the blood brain barrier, fibroblast activation, and collagen deposition. Tissue returned to normal conditions by 7 days in sciatic nerve and 21 days in the DRG. PRF treatment does not rely on thermal injury to achieve its clinical effect [19]. PRF is known to be nondestructive, but, it may produce heat spikes exceeding 45℃-50℃, causing destructive effects on neurons [35]. In this study, although we could not completely exclude thermal injury by PRF, we maintained the temperature of needle tip at 42℃ to minimize the probable thermal injury. The parameters used for PRF in this study were the same as other reports [19].
PRF is considered as a nondestructive and reversible neuromodulation intervention [36]. Neuromodulation is a normal property of the nervous system that regulates or modifies electrical impulses flowing through neural tissues. Neuromodulation of nociceptive information takes place in spinal synapses [36]. PRF may relieve neuropathic pain by inducing long-term depression of synaptic transmission in the spinal cord [37]. Tanaka et al. [34] reported that decreased mechanical allodynia was observed at an early stage within 1 week of applying PRF. Aksu et al. [38] reported that PRF stimulation performed on 5-6 lumbar dorsal root improves hyperalgesia in sciatic neuropathy induced rabbit. However, animal study on effect of PRF conducted in peripheral neuropathy area is still lacking. In this study, improvement of mechanical allodynia and thermal hyperalgesia was observed after PRF stimulation to the injured sciatic nerve area. Both mechanical allodynia and thermal hyperalgesia was improved on 7 days after applying PRF. The TNF-α immunoreactivity was decreased in spinal cord and sciatic nerve on 7 days after PRF indicating that PRF stimulation affects modulation of inflammatory response in the injured area. Following peripheral nerve injury, immunoregulatory factors such as TNF-α are induced at the site of injury and undergo retrograde transport to the DRG and anterograde transport from DRG to the site of injury. In a sciatic neuropathy rat model, down-regulation of TNF-α in both the sciatic nerve and DRG indicates that PRF can reduce the expression of pro-inflammatory cytokines in adjacent tissues either directly or indirectly [39]. As TNF-α is one of the major players in the development and maintenance of neuropathic pain [12], the decreased expression of TNF-α, and modulation of its target cytokines, indicates that PRF potentially alleviates neuropathic pain states by attenuating neuroinflammation at the molecular level [12].
While PRF applied to DRG induces changes in the neurons of dorsal horn, percutaneous application of PRF can be targeted to neural tissues and nociceptors in the periphery [40]. Application of PRF to a patient with meralgia paresthetica under ultrasonographic guidance showed excellent pain relief [14]. A randomized controlled trial demonstrated that the application of PRF to suprascapular nerve under ultrasonographic guidance combined with physical therapy results in pain relief, reduced disability, and improvement of passive range of shoulder motion [15]. Although recent application of PRF to peripheral nerve for easy access control of neuropathic pain is now increasing, it has not been extensively studied. This prompted us to select PRF application to the peripheral nerve.
There were several limitations to this study. We did not use percutaneous approach using ultrasonography for PRF treatments. In human studies, it is obviously less invasive to use percutaneous approach to the peripheral nerve causing neuropathic pain under ultrasonographic guidance. As this was an animal study, we exposed the sciatic nerve to secure clear visibility. The changes of spinal glial cells and glial activation signaling pathway involved in the transmission of neuropathic pain after peripheral nerve injury were not studied. Activated glial cells after nerve injury release various pain-modulating substances, however, studies on other inflammatory markers were insufficient.
In conclusion, these results suggested that PRF stimulation applied proximal to the injured peripheral nerve alleviated neuropathic pain by CCI. Furthermore, the reduction of neuropathic pain after PRF treatments may be associated with neuromodulation effects through the inhibition of the expression or anterograde transportation of TNF-α.
Go to :
 XML Download
XML Download