

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Restoring a sufficient liver volume is essential for patient survival and quality of life after surgical resection. The peak liver regeneration after surgical resection occurs on an average within 7 days. A substantial proportion of the patients undergoing liver resection often face liver dysfunction, such as liver fibrosis or cirrhosis [1], increasing patient morbidity and mortality resulting from the reduced regenerative capacity of the liver. Therefore, maximizing liver regeneration after surgical resection is a prerequisite for preventing complications and tracking morbidity and mortality [2].
Recent studies have shown the potential of stem cells to enhance liver regeneration [3]. Among various stem cell sources, adipose-derived stem cells (ASCs) have distinct advantages, including the ease of attainment and processing, long-term in vitro maintenance, and high proliferative capacity [4]. However, despite the numerous benefits of stem cells, their clinical application is still challenging due to several limitations, such as their short in vivo viability and potential for malignant transformation [5]. Recently, the use of stem cell secretome instead of stem cells has been actively studied in an attempt to overcome these limitations [6] secretome refers to substances secreted by the stem cells into the surrounding extracellular space [7]. The use of the secretome is supported by evidence that they mediate the main mechanism of action of stem cells [8]. Conditioned media (CM) obtained from MSCs were found to contain high levels of paracrine cytokines and growth factors that promote tissue repair in numerous in vitro and in vivo experimental models [6].
Survivin is a member of the inhibitor of apoptosis (IAP) protein family and is one of the representative antiapoptotic proteins [9]. It is copiously expressed during human fetal development but is infrequently present in adult tissues. Survivin inhibits cell apoptosis by inhibiting caspase activity [10] and is highly expressed in various malignancies. However, it is also expressed in various differentiated adult tissues, particularly during proliferation and regeneration [11]. We aimed to investigate whether the regenerative capacity of the liver was enhanced by the secretome obtained from survivin-overexpressing ASCs. To achieve this, we generated the secretome from survivin-transfected ASCs and attempted to determine the in vitro and in vivo regenerative capacity of the secretome.
Go to : 
METHODS
Culture of adipose-derived stem cells
ASCs were plated into culture flask in Dulbecco’s Modified Eagle Medium with low glucose (DMEM/low glucose; Hyclone, Logan, UT, USA) supplemented with 0.1 mg/mL of streptomycin (Gibco BRL, Carlsbad, CA, USA), 100 U/mL of penicillin (Gibco), 10% fetal bovine serum (FBS; Hyclone) at 37°C in humidified chamber containing 5% carbon dioxide [812].
Transfection and preparation of secretome
ASCs that reached 70%–80% into culture flask were transfected with 1 µg survivin. After 24 hours, ASCs were refed serum-free DMEM/low glucose for 24 hours. Then, the secretome was obtained 200 µL amount at ASCs, the CM were concentrated by 25 fold, by ultrafiltration device with a 3-kDa cutoff (Amicon Ultra-15; Millipore, Bedford, MA, USA) [8].
Cell viability assay
The alpha mouse liver 12 (AML12) was cultured in the DMEM/F12 (Ham) media (Gibco) supplemented with 100 µg/mL streptomycin (Gibco), 100 U/mL penicillin (Gibco), 10% FBS (Hyclon), insulin-transferrin-selenium (Gibco), 40 ng/mL dexamethasone (Sigma-Aldrich, St. Louis, MO, USA). AML12 was plated 1 × 104 cells/well in 96 well plates. After 24 hours, the 96-well plates were treated with 2.5 µL of secretomes after incubation for 24 hours at 37°C. Cell viabilities of secretome-treated AML12 was measured by the Ez-Cytox cell viability assay kit (Daeil Lab Service Corp., Ltd., Seoul, Korea). The absorbance was determined with an enzyme-linked immunosorbent assay (ELISA) reader (model microplate reader; BioTek Instruments Inc., Winooski, VT, USA) at 450 nm.
Animal experiments
Using 5-week-old male BALB/c mice (Orient Corp., Seongnam, Korea), we generated the model of 70% partial hepatectomy (PH) by surgically removing 2/3 of the liver [13]. The hepatectomy mice were divided into 4 groups; control (n = 25), PH-only (n = 25), control secretome (n = 25), and survivin-secretome (n = 25). Approximately 1 hour after PH, control secretome and survivin-secretome groups were intravenously administered 0.1 mL of control secretome and survivin-secretome. The 70% PH mice were euthanized to obtain serum, liver at the determined days.
The experiments protocol was approved by the Institutional Animal Care and Use Committee of School of Medicine, The Catholic University of Korea (No. 2020-0303-01).
Western blot analysis
AML12 and PH mice liver specimens were lysed by the Ez RIPA-Lysis kit (ATTO Corp., Tokyo, Japan). Proteins were performed by western blot using primary antibodies: signal transducer and activator of transcription 3 (STAT3), phospho-STAT3 (p-STAT3), hepatocyte growth factor (HGF), vascular endothelial growth factor (VEGF), myeloid cell leukemia-1 (Mcl-1), proliferating cell nuclear antigen (PCNA), bcl-2-like protein 4 (Bax), survivin, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and β-actin from Cell Signaling Technology (Beverly, MA, USA), HRP conjugated anti-rabbit immunoglobulin G (IgG) and anti-mouse IgG from Vector Lab. (Burlingame, CA, USA).
Real-time polymerase chain reaction
Using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), RNA was extracted according to the manufacturer’s instructions. Using RT-premix kit (TOYOBO, Osaka, Japan), reverse transcription was performed with 1 µg RNA according to the manufacturer’s instructions [12]. The primers used for SYBR Green real-time quantitative polymerase chain reaction (PCR) were as follows: human HGF forward 5′-TGCTGTCCTGGATGATTTTG-3′ and reverse 5′-AGTGTAGCCCCAGCCATAAA-3′; human VEGF forward 5′-TCTTCAAGCCATCCTGTGTG-3 and reverse 5′-ATCTGCATGGTGATGTTGGA-3′; human survivin forward 5′-CAAGGAGCTGGAAGGCTGG-3′ and reverse 5′-GTTCTTGGCTCTTTCTCTGTCC-3′; Mcl-1 forward 5′-GGGCAGGATTGTGACTCTCATT-3′ and reverse 5′-GATGCAGCTTTCTTGGTTTATGG-3′; human Bax forward 5′-TGGAGCTGCAGAGGATGATTG-3′ and reverse 5′-GAAGTTGCCGTCAGAAAACATG-3′; human GAPDH forward 5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse 5′-TGGTGAAGACGCCAGTGGA-3′; mouse HGF forward 5′-GGCAAGGTGACTTTGAATGA-3′ and reverse 5′-CACATGGTCCTGATCCAATC-3′; mouse VEGF forward 5′-CCGAAACCATGAACTTTCTGA-3′ and reverse 5′-CCATTCATGGGACTTCTGCTC-3′; mouse Mcl-1 forward 5′-AAAGGCGGCTGCATAAGTC-3′ and reverse 5′-TGGCGGTATAGGTCGTCCTC-3′; mouse Bax forward 5′-CTGCAGAGGATGATTGCCG-3′ and reverse 5′-TGCCACTCGGAAAAAGACCT-3′; mouse hepatocyte nuclear factor 4 α (HNF4α) forward 5′-AGGCAATGACTACATCGTCCC-3′ and reverse 5′-CAGACCCTCCGAGAAGCATC-3′ [14]; mouse albumin forward 5′-TGAAGTTGCCAGAAGACATCC-3′ and reverse 5′-CAAGTTCCGCCCTGTCATCTG-3′ [14]; mouse epithelial cell adhesion molecule (EpCAM) forward 5′-AGCTCAGGAAGAATGTG-3′ and reverse 5′-TGAAGTACACTGGCATTGACG-3′ [15]; mouse GAPDH forward 5′-CGACTTCAACAGCAACTCCCACTCTTCC-3′ and reverse 5′-TGGGTGGTCCAGGGTTTCTTACTCCTT-3′. Reactions were performed by the Applied Biosystems Step one plus Real-time PCR system (Thermo, Carlsbad, CA, USA).
Serology test
Blood was collected for each mouse. Then, centrifuged for 9,500 ×g in 10 minutes serum was collected. We performed liver injury concentrations using ALT and AST with the VetTest Chemistry Analyzer (IDEXX Laboratories Inc., Westbrook, ME, USA) [8].
Enzyme-linked immunosorbent assay
The concentrations of mouse tumor necrosis factor (TNF)-α and interleukin (IL)-6 were measured by ELISA kit (Labiscoma Corp., Seoul, Korea) [12].
Immunohistochemistry
For immunohistochemical (IHC) analysis, formalin-fixed, paraffin-embedded tissue sections were deparaffinized, rehydrated in graded series of alcohols. The antibodies used for this experiment were PCNA mouse polyclonal antibodies (1:3,000; Abcam, Cambridge, UK), B-cell lymphoma-extra large (Bcl-xL) rabbit polyclonal antibodies (1:1,000; Abcam), Bim rabbit polyclonal antibodies (1:200; Abcam), survivin rabbit polyclonal antibodies (1:400; Abcam). Then, the samples were examined using a laser-scanning microscope (Eclipse TE300; Nikon, Tokyo, Japan).
Statistical analysis
Analysis of all data was made by SPSS ver. 11.0 (SPSS Inc., Chicago, IL, USA). Data were presented as mean ± standard deviation. Using the Kruskal-Wallis test, groups were compared. The P-values of <0.05 were regarded as statistically significant.
Go to :
RESULTS
Characterization of survivin-overexpressing adipose-derived stem cells
Survivin-overexpressing ASCs were generated by transfecting ASCs with survivin gene-conveying plasmids (Fig. 1A). The secretome obtained from the CM after transfecting ASCs with the survivin gene-conveying plasmid was termed the survivin-secretome. Western blot analysis showed an upregulated expression of survivin in survivin-overexpressing ASCs (Fig. 1B). In real-time PCR, survivin-overexpressing ASCs showed increased HGF and VEGF messenger RNA (mRNA) expression as well as survivin mRNA compared to the control ASCs (P < 0.05) (Fig. 1C).
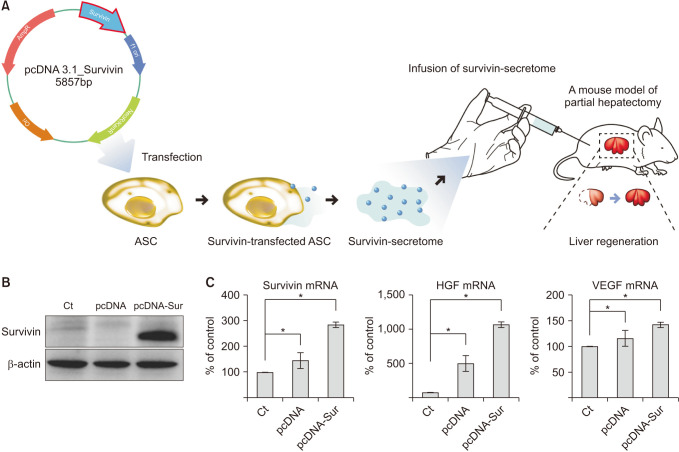 | Fig. 1Characterization of survivin-overexpressing adipose-derived stem cells (ASCs). (A) Study design. After generation of survivin-secretome from survivin-overexpressing ASCs, the liver regenerative potential of survivin-secretome was determined using a mouse model of 70% PH. (B) Western blot analysis showing the upregulated expression of survivin in the survivin-overexpressing ASCs. (C) Real-time polymerase chain reaction comparing messenger RNA (mRNA) expression between control secretome and survivin-secretome. Values are displayed as mean ± standard deviations from 3 independent experiments. *P < 0.05. HGF, hepatocyte growth factor; PH, partial hepatectomy; VEGF, vascular endothelial growth factor.
|
In vitro characterization of the survivin-secretome
The survivin-secretome was obtained from the CM of survivin-overexpressing ASCs after processing at a 25-fold concentration using ultrafiltration units with a 3-kDa cutoff. Subsequently, thioacetamide (TAA), a hepatotoxin, was administered to AML12 hepatocytes, and the cell viability between the groups treated with either control secretome or survivin-secretome was compared. The survivin-secretome group showed significantly increased cell viability compared to the control group (P < 0.05) (Fig. 2A). Real-time PCR showed that the survivin-secretome group significantly decreased the expression of a proapoptotic marker (Bax) and increased the expression of an antiapoptotic marker (Mcl-1) (P < 0.05) (Fig. 2B). Next, the effects of survivin-secretome were determined by western blot analysis. It showed that the survivin-secretome group significantly increased the expression of p-STAT3, HGF, VEGF, Mcl-1, and survivin compared to the control secretome group (P < 0.05) (Fig. 2C).
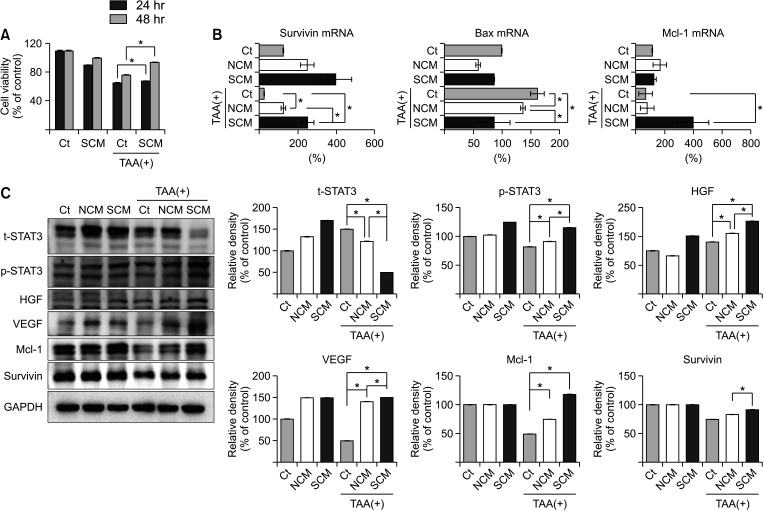 | Fig. 2In vitro validation of survivin-secretome. (A) Cell viability test of AML12 hepatocytes treated with either control secretome or survivin-secretome. (B) Real-time polymerase chain reaction comparing messenger RNA (mRNA) expression of AML12 hepatocytes treated with either control secretome or survivin-secretome. (C) [Left] Western blot of various markers in the AML12 hepatocytes treated with either control secretome or survivin-secretome. [Right] The relative densities of markers in each group. Using ImageJ software, the relative density of individual markers was quantified and then normalized to the density of β-actin in each group. Values are displayed as mean ± standard deviations from 3 independent experiments. *P < 0.05. Ct, control; SCM, survivin conditioned media (survivin-secretome); NCM, normal conditioned media (control secretome); TAA, thioacetamide; Bax, bcl-2-like protein 4; Mcl-1, myeloid cell leukemia 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; STAT3, signal transducer and activator of transcription 3; t-STAT3, total STAT3; p-STAT3, phospho-STAT3; HGF, hepatocyte growth factor; VEGF, vascular endothelial growth factor.
|
In vivo validation of survivin-secretome
A mouse model of 70% PH was generated by surgical resection of the hepatic lobules [13]. One hour after PH, the mice were administered normal saline (PH-only group), control secretome (control secretome group), and survivin-secretome (survivin-secretome group) through the tail vein. Each group included 2 subgroups (continuous and interrupted groups) according to the ways of sample acquisition (Fig. 3A). The expression levels of IL-6 and TNF-α (systemic inflammatory markers) in the sera of each mouse group were compared (Fig. 3B). The treatment groups (control secretome and survivin-secretome groups) showed significantly decreased serum levels of these markers on PH 2 and 3 days compared to the PH-only group (P < 0.05). Of the 2 secretome-administered groups, the survivin-secretome group showed the lowest levels of these markers (P < 0.05). Subsequently, the serum levels of liver transaminases (ALT and AST) were compared (Fig. 3C). Likewise, the survivin-secretome group showed the lowest levels of ALT and AST on PH 2 days (P < 0.05). Figure 3D shows that the size of the extracted liver was increased in the treatment group (control secretome and survivin-secretome) than in the PH-only group. Liver regeneration rate (percentage) was calculated as the ratio of the extracted liver weight to 0.7 times the excised liver weight [16]. The 2 treatment groups showed a significantly higher liver regeneration rate than the PH-only group (P < 0.05). Of the 2 treatment groups, the survivin-secretome group showed the highest liver regeneration on PH 3 and 7 days (P < 0.05) (Fig. 3E).
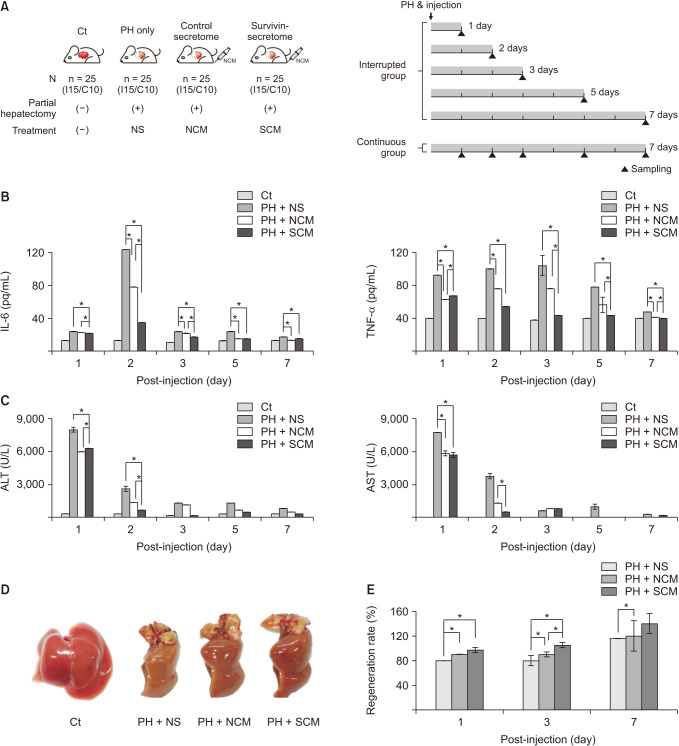 | Fig. 3In vivo validation of survivin-secretome. (A) Study design of in vivo experiment. One hour after partial hepatectomy (PH), the mice were administrated with normal saline (PH-only group), control secretome (control secretome group), and survivin-secretome (survivin-secretome group) through the tail vein, respectively. Each group included 2 subgroups (continuous and interrupted groups) according to the method of acquiring samples. (B) Enzyme-linked-immunosorbent assay showing the serum concentration of systemic inflammatory markers (tumor necrosis factor [TNF]-α and interleukin [IL]-6) in each group. (C) Serologic test showing the serum concentration of transaminases (ALT and AST) in each group. (D) Size comparison of the representative liver specimens in each group. (E) Comparison of liver regeneration rates based on the weights of extracted liver specimens in each group. Values are displayed as mean ± standard deviations from 3 independent experiments. *P < 0.05. Ct, control; NS, normal saline; NCM, normal conditioned media (control secretome); SCM, survivin conditioned media (survivin-secretome).
|
In vivo effects of survivin-secretome on the liver specimens
After euthanizing the mice 2 on PH 2 days, we harvested liver specimens and examined the expression of various markers related to liver regeneration at the RNA and DNA levels. Real-time PCR revealed that the survivin-secretome group showed the highest RNA expression levels of HGF, VEGF, Mcl-1, HNF4α, and albumin in all the mouse groups with PH (P < 0.05) (Fig. 4A). Also, the survivin-secretome group showed the lowest RNA expression of Bax and EpCAM (P < 0.05). Subsequently, western blot analysis was performed to compare the expression of various proteins. The survivin-secretome group showed the highest expression levels of p-STAT3, HGF, VEGF, PCNA, and Mcl-1 in all mouse groups with PH (P < 0.05) (Fig. 4B). The survivin-secretome group also showed the lowest expression of Bax (P < 0.05). Taken together, it appears that the intravenous injection of survivin-secretome cumulates in higher antiapoptotic, proliferative, and angiogenic effects than the injection of control secretome in 70% hepatectomized mice.
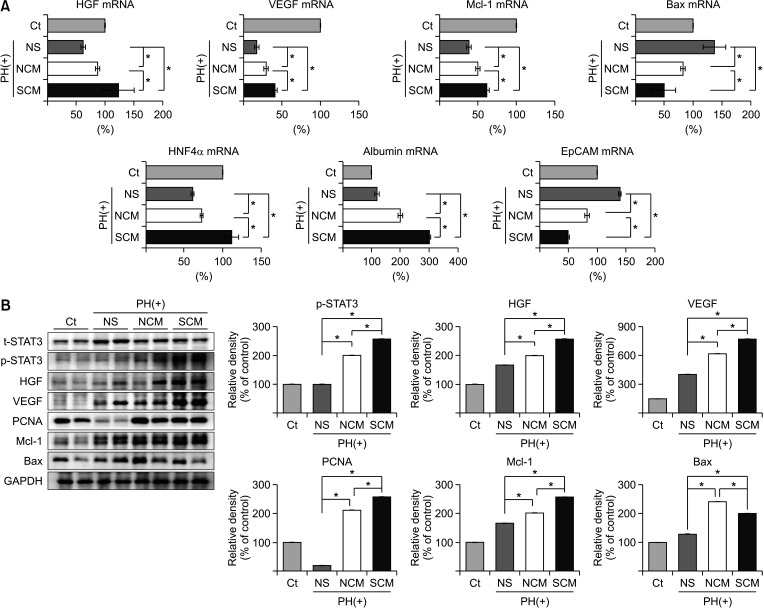 | Fig. 4In vivo effects of survivin-secretome on the liver specimens. (A) Real-time polymerase chain reaction comparing messenger RNA (mRNA) expression in the liver specimens treated with either control secretome or survivin-secretome. (B) [Left] Western blot showing the expression of various markers in the liver specimens in each group. [Right] The relative densities of markers in each group. Using ImageJ software, the relative density of individual markers was quantified and then normalized to the density of β-actin in each group. Values are displayed as mean ± standard deviations from 3 independent experiments. *P < 0.05. PH, partial hepatectomy; Ct, control; NS, normal saline; NCM, normal conditioned media (control secretome); SCM, survivin conditioned media (survivin-secretome); HGF, hepatocyte growth factor; VEGF, vascular endothelial growth factor; Mcl-1, myeloid cell leukemia 1; Bax, bcl-2-like protein 4; HNF4α, hepatocyte nuclear factor 4 α; EpCAM; epithelial cell adhesion molecule; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; STAT3, signal transducer and activator of transcription 3; t-STAT3, total STAT3; p-STAT3, phospho-STAT3; PCNA, proliferating cell nuclear antigen.
|
In vivo effects of survivin-secretome on the histology of the liver
H&E and IHC staining were performed to determine the effects of survivin-secretome on the liver of partially hepatectomized mice (Fig. 5). H&E staining showed that the survivin-secretome group exhibited the lowest expression of Bcl-xL (P < 0.05). Survivin IHC staining demonstrated that the survivin-secretome group exhibited the highest level of survivin in all groups (P < 0.05). The survivin-secretome group showed the highest expression of a proliferation marker (PCNA) (P < 0.05), suggestive of the proproliferative effects of the survivin-secretome. The survivin-secretome group also showed the highest expression of an antiapoptotic marker (Bcl-xL) and the lowest expression of a proapoptotic marker (Bim), demonstrating the antiapoptotic effect of survivin-secretome (P < 0.05). Taken together, the results of various IHC stains show that the survivin-secretome promotes liver regeneration and prevents cell apoptosis higher than control secretome.
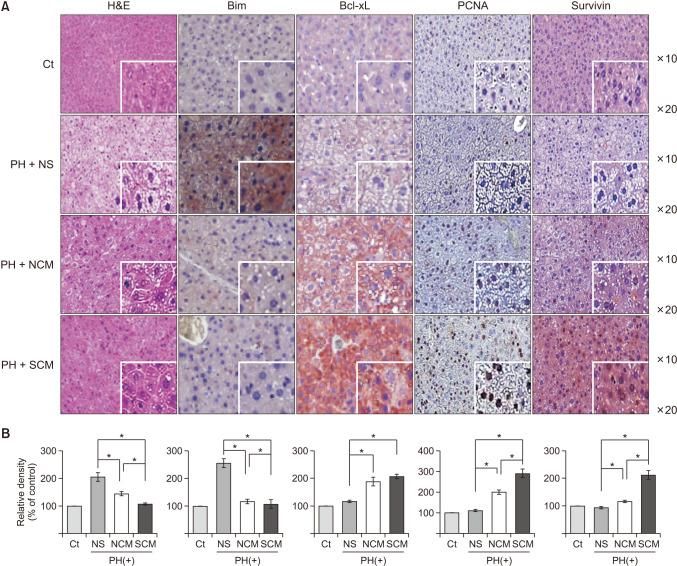 | Fig. 5Immunohistochemical stains of the liver specimens in each group. (A) H&E and immunohistochemical of bcl-2-like protein 11 (Bim), B-cell leukemia-extra large (Bcl-xL), proliferating cell nuclear antigen (PCNA), and survivin in each group. (B) Using Imag J software, the percentages of immunoreactive areas were measured and displayed as relative values to those in normal livers. Values are displayed as mean ± standard deviations from 3 independent experiments. *P < 0.05. H&E, Hematoxylin and Eosin; Ct, control; PH, partial hepatectomy; NS, normal saline; NCM, normal conditioned media (control secretome); SCM, survivin conditioned media (survivin-secretome).
|
Go to :
DISCUSSION
This study focused on whether the secretome obtained from the survivin-overexpressing ASCs enhanced the proproliferative potential in the liver compared to the control secretome. In the experiments, the survivin-secretome significantly increased the cell viability. The survivin-secretome also increased the protein expression levels of markers related to proliferation (HGF, VEGF, and p-STAT3) and antiapoptosis (Mcl-1) and reduced the expression of the proapoptotic marker, Bax, in TAA-treated AML12 hepatocytes. In the experiments using hepatectomized mice, the survivin-secretome group showed the lowest serum levels of proinflammatory cytokines, such as TNF-α and IL-6. The serum levels of liver transaminases (ALT and AST) were also the lowest in the survivin-secretome group. The survivin-secretome group also exhibited the highest liver regeneration based on the liver weight on PH 3 and 7 days. In the subsequent liver specimen analysis, the specimens of survivin-secretome exhibited the highest expression of p-STAT3, HGF, VEGF, PCNA, and Mcl-1, and the lowest expression of Bax, suggestive of the enhanced proproliferative and antiapoptotic potential of survivin-secretome. Taken together, it appears that the survivin-secretome has an enhanced potential for proproliferation and antiapoptosis compared to the control secretome in both in vitro and in vivo models of liver hepatectomy.
Survivin is the smallest member of the IAP family of proteins and is encoded by the BIRC5 gene. The survivin gene spans 15 kb of DNA and is located on chromosome 17 at band q25. Structurally, survivin includes a single baculovirus IAP repeat (BIR), a B70-amino-acid zinc-finger fold that is the hallmark of all IAPs, and an extended –COOH terminus α-helical coiled-coil, which are intimately linked with its function as an IAP [17]. A mutation in the BIR domain leads to an abrogation of its ability to inhibit apoptosis [18]. Survivin plays important roles in various cellular mechanisms, including cell cycle regulation, proliferation, apoptosis, and angiogenesis [13]. Survivin is present in the cytoplasm, nucleus, and mitochondria [19]. Survivin is highly exhibited in most human tumors and fetal tissues; however, it is entirely absent in differentiated cells.
This study showed that the survivin-secretome has the potential to enhance regeneration of the damaged liver. The survivin-secretome increased the protein expression levels of the markers related to proliferation (HGF, VEGF, and p-STAT3) in both in vitro and in vivo models of liver hepatectomy. Moreover, the injection of survivin-secretome into the partially hepatectomized mice induced the highest liver regeneration on PH 3 and 7 days. In the analysis of both hepatocellular carcinoma (HCC) cell lines and HCC tissues, Ito et al. [20] found that survivin expression levels were strongly correlated with the proliferation index in HCC tissues. The overexpression of survivin resulted in a decreased G0/G1 phase and an increased S phase in all HCC cell lines. They also showed that survivin promoted cell proliferation by interacting with cyclin-dependent kinase 4 (Cdk4) and releasing p21 from Cdk4. Our study suggests that survivin-secretome contains higher levels of survivin, thus exerting similar proproliferative properties as survivin.
The survivin-secretome also exerted profound antiapoptotic effects in both in vitro and in vivo models of liver hepatectomy. It increased the antiapoptotic marker (Mcl-1) and decreased the pro-apoptotic marker (Bax) in TAA-treated AML12 hepatocytes. Subsequently, the liver specimens of the survivin-secretome-injected mice exhibited the highest expression of Mcl-1 and the lowest expression of Bax. Several IAP family members have been shown to suppress apoptosis by directly inhibiting caspases via the BIR domains [21]. The structure of survivin has been compared to that of another IAP family member, XIAP, which includes 3 BIR domains. XIAP inhibits caspase-3 and caspase-7 via a linker region between the first 2 BIR domains, and also binds to and inhibits caspase-9 through its third BIR domain (BIR3) [22]. The BIR domain of survivin appears closely related to the three-dimensional structure of the BIR3 domain of XIAP, suggesting the possibility that survivin binds to caspase-9 [23]. Fas receptor (CD95, TNF receptor superfamily member 6) is a death receptor localized on the surface of various cells, which triggers a signal transduction pathway leading to apoptosis. Interestingly, Fas-induced apoptosis is associated with the upregulation of survivin in the mitochondria, nucleus, and cytosol [24]. Upon Fas stimulation in cell culture, survivin interacts with Cdk4, which leads to release p21 from its complex with Cdk4 [25] making it possible for p21 to complex with caspase-3, the initial step in inactivating caspase-3 in the mitochondria. However, further research is required to determine how survivin inhibits apoptosis by suppressing caspase-3 and other caspases.
Our experiments also displayed that the survivin-secretome increased the expression of VEGF in both in vitro and in vivo models of liver hepatectomy. VEGF is a key transcription factor that regulates the angiogenesis of endothelial cells. Wang et al. [26] discovered that survivin expression in glioma cells prompted the production and secretion of VEGF and basic fibroblast growth factor, implying that survivin plays a vital role in transmitting angiogenic stimuli to the associated vasculature in glioma cells. Besides, survivin exerts favorable effects on angiogenesis by promoting the survival and proliferation of endothelial cells [2728]. In addition, VEGF participates in the process of liver regeneration. Injection of VEGF promoted the proliferative activity of hepatocytes, demonstrating the significant role of VEGF in liver regeneration [29]. Taniguchi et al. [30] showed that VEGF promoted the proliferation of sinusoid endothelial cells during liver regeneration, which leads to the enhanced proliferation of hepatocytes. Therefore, the survivin-secretome appears to promote liver regeneration and angiogenesis, in part, by increasing VEGF levels.
In conclusion, this study purposed to determine the role of the secretome obtained from survivin-overexpressing ASCs. In the in vitro experiments, survivin-secretome enhanced cell viability and promoted the expression of proproliferative and antiapoptotic markers in TAA-treated AML12 hepatocytes. Subsequent intravenous injection of survivin-secretome into the partially hepatectomized mice enhanced cell proliferation, anti-apoptosis, and VEGF expression, all of which are beneficial for liver regeneration and tissue repair. Therefore, the use of secretome secreted by survivin-overexpressing ASCs could be an effective way to enhance liver regeneration and repair for the treatment of liver injury.
Go to :
 XML Download
XML Download