

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The recent advances in perinatology have shifted the goal of neonatal intensive care from improving the survival rates of newborns to enhancing their quality of life. Neurodevelopmental outcomes of surviving high-risk infants play an important role in determining their quality of life.1 Previous studies have reported that white matter injury of the brain in preterm infants is correlated with sequelae, such as cerebral palsy or cognitive deficit.2 Several studies examining the neurodevelopment of preterm infants have attempted to determine the pathogenesis of white matter injury. The spectrum of periventricular white matter injury (PWMI) includes cerebral injuries ranging from focal to extensive cerebral white matter lesions.3 Although the overall incidence of cystic necrosis and microcysts has decreased, diffuse white matter injury is commonly observed in preterm infants.4 According to Volpe,2 the etiology of diffuse PWMI involves less severe episodes of ischemia associated with short penetrating arteries of superficial end zones, which may account for extensive myelination impairment.
Lipopolysaccharide (LPS), a component of the gram-negative bacterial membrane, exerts cytotoxic effects on preterm brain cells, especially on those belonging to the oligodendrocyte lineage. In addition to activating the microglial cells through Toll-like receptor 4, LPS mediates brain damage by inducing the release of various cytotoxic mediators, including cytokines and reactive oxygen species.567 Previous studies have investigated the maturation-dependent susceptibility of oligodendrocytes to hypoxia-ischemia insult.8910 Miller et al.11 treated the primary oligodendrocyte cell cultures with LPS-activated microglial cells and reported that immature and mature oligodendrocytes at different developmental stages exhibited differential responses to activated microglial cells. Several studies have examined the effect of perinatal and neonatal infection and inflammation on the preterm brain. Brain magnetic resonance imaging analyses have revealed that perinatal and/or neonatal infections may alter white matter development. Additionally, perinatal and/or neonatal infections can adversely affect neurodevelopmental outcomes to varying degrees in preterm infants depending on the gestational age.1213141516
The oligodendrocyte lineage cells exhibit differential susceptibility to LPS-induced inflammation depending on the developmental stage. Diffuse white matter injury caused by infection and inflammation involves acute cell death, followed by the rapid regeneration of preoligodendrocytes (pre-OLs), which fail to mature into myelinating oligodendrocytes. A similar pattern of impaired cellular maturation has been reported in studies examining the effect of proinflammatory cytokines in preterm-equivalent rodents.17 However, the effects of age at which inflammation or infection is induced and repeated episodes of infection and inflammation on the differentiation of oligodendrocytes throughout the critical developmental stages have not been previously reported.
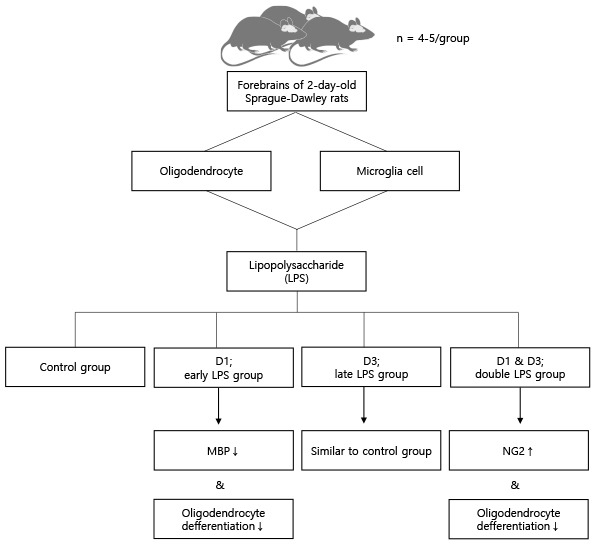
This study aimed to investigate the effect of LPS on oligodendrocyte lineage cells at different developmental stages using a microglial cell and oligodendrocyte co-culture model. To establish the recurrent inflammation model, the oligodendrocyte lineage cells were exposed to LPS at two developmental stages (double-hit LPS insult). Furthermore, the effects of direct and recurrent LPS stimulations were evaluated at the cellular level.
Go to : 
METHODS
Primary oligodendrocyte-microglial cell co-culture
The primary cultures of oligodendrocytes and microglia cells were prepared from the forebrains of 2-day-old Sprague–Dawley rats by using a shaking method of the mixed glial cultures, following the protocols of Miller et al.11 with minor modifications. The meninges were removed and the brain tissue was triturated and dissociated mechanically. The dissociated brain tissue was maintained in basal media comprising Dulbecco’s modified Eagle’s medium supplemented with GlutaMAX (Gibco, Carlsbad, CA, USA) and 10% heat-inactivated fetal bovine serum (FBS; Invitrogen, Carlsbad, CA, USA) at 37°C and 5% CO2. The medium was replaced once every 3 days.
After 9 days of culture, the mixed glial culture comprised a layer of astrocytes at the bottom of the uncoated T-75 flasks with oligodendrocyte progenitor cells (OPCs) and microglial cells growing over the astrocytes. The OPCs were isolated from the mixed glial cells by shaking for 16 hours overnight. The detached OPCs were collected by centrifugation at 50 rpm and resuspended at a density of 70,000 cells/cm2 in oligodendrocyte culture medium to induce the maturation of OPCs into oligodendrocytes. The oligodendrocyte culture medium (insulin [I6634; Sigma, St. Louis, MO, USA], holo-transferrin [T0665; Sigma], B27 [Gibco], GlutaMAX [Gibco], 0.5% FBS [Invitrogen], and 10 µg/mL ciliary neurotrophic factor [PeproTech, Cranbury, NJ, USA]) was supplemented with 100 × oligodendrocyte-supplement (bovine serum albumin, progesterone, putrescine, sodium selenite, and 3, 3′, 5-triiodo-L-thyronine; all from Sigma). Microglial cells were purified from the astrocyte layers of the same mixed glial culture that was used for harvesting OPCs. To isolate the microglial cells, the culture was shaken for 2 hours at 200 rpm. Microglial cells were resuspended in the same basal medium as OPCs, but with insulin (I1882; Sigma). Purified OPCs contained < 5% glial fibrillary acidic protein-positive astrocytes and < 0.5% microglial cells, while purified microglial cells contained < 0.1% astrocytes and < 1% oligodendrocytes. The next day 250 µL of oligodendrocyte culture medium in the mixed glial culture (500 µL) was replaced with 250 µL of microglial cell (D1 stage) suspension at a cell density ratio of 1:1. The cells were maintained in a humidified incubator under 8.5% CO2 conditions.
LPS application
The OPC-microglial cell co-culture was treated with LPS at D1 and/or D3 developmental stages. Microglial cells produce cytokines and reactive oxygen species, which adversely affect the maturation and survival of immature oligodendrocytes.
Our model is a modified type of an LPS-induced brain injury model proposed by Miller et al.11 They used 0.001, 0.01, and 0.1 µg/mL dose of LPS in their experiment. There were other 3 studies which used 1 µg/mL dose of LPS,6718 and 1 study used 0.01 µg/mL.19 So, in the preliminary experiment, we set the LPS dose within the range of 0.01 to 1 µg/mL, and decided to increase or decrease it if necessary.
To simulate double-hit exposure model, we needed to find the dose which did not affect cells significantly solely, but made any effect when the dose was repeated. To determine the dose of LPS, the microglia and oligodendrocyte co-culture was treated with various concentrations of LPS (0.01, 0.1, and 1 µg/mL) at the D3 stage. The LPS dose of 0.01 µg/mL was used to simulate double-hit inflammation because this dose solely did not make significant effect on myelin basic protein (MBP)+ and ionized calcium binding adaptor molecule 1 (IBA1, also known as allograft inflmmatory factor 1) cell numbers. Next, the co-culture was treated with 0.01 µg/mL LPS, which was the lowest dose of LPS that did not impair oligodendrocyte differentiation, on various dosing schedules. To evaluate the developmental stage-dependent effect of LPS stimulation, the early LPS group was treated with 0.01 µg/mL LPS at the D1 and with vehicle (normal saline) at the D3. And the late LPS group was treated with vehicle at the D1 and with 0.01 µg/mL LPS at the D3. Double LPS group, which was designed to simulate the double-hit effect of LPS stimulation, was treated with 0.01 µg/mL LPS at both the D1 and D3 (Fig. 1).
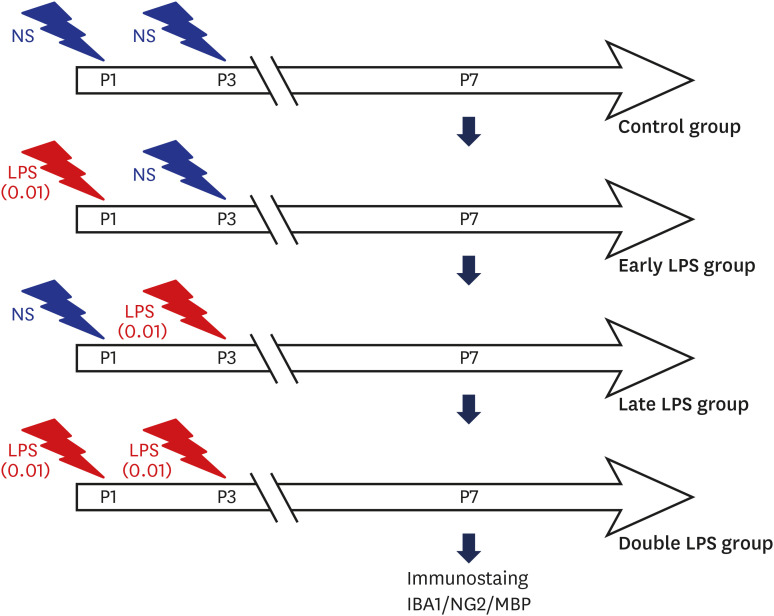 | Fig. 1Schematic outline of the experimental protocol. The oligodendrocyte-microglial cell co-culture was treated with the lowest dose of LPS (0.01 µg/mL), which does not inhibit oligodendrocyte differentiation, at the developmental stages D1 (early LPS group), D3 (late LPS group), or D1 and D3 (double LPS group).LPS = lipopolysaccharide, MBP = myelin basic protein.
|
Immunostaining
The cells were fixed with 2% formaldehyde prepared from paraformaldehyde. The anti-neural glial antigen 2 (NG2, also known as chondroitin sulfate proteoglycan 4), anti-MBP, and anti-IBA1 antibodies were purchased from Millipore (Burlington, MA, USA; AB5320), BioLegend (San Diego, CA, USA; 808402), and Abcam (Cambridge, UK; ab153696), respectively. The secondary antibodies were purchased from Thermo Fisher Scientific (Carlsbad, CA, USA). The antibodies were diluted in phosphate-buffered saline (pH 7.4) supplemented with 2% goat serum. The coverslips were rinsed twice between antibody applications.
For NG2 immunostaining, the cells were permeabilized with 0.3% Triton X-100 (Sigma) for 10 minutes and incubated with anti-NG2 antibody (1:500) for 2 hours at room temperature, followed by incubation with goat anti-rabbit Alexa 488 (1:200; Thermo Fisher Scientific) for 2 hours. To perform immunostaining of MBP and IBA1, the cells were permeabilized with 0.3% Triton X-100 for 10 minutes and incubated with anti-MBP antibodies (1:500) for 1 hour, followed by incubation with goat anti-rabbit Alexa 488 or anti-goat cy3 (1:100; Sigma) for 1 hour. The cells were counted by an investigator blinded to the experimental conditions using a Zeiss Axioplan 2 microscope (Carl Zeiss, White Plains, NY, USA). Five random, consecutive fields were counted in each coverslip under 200 × magnification. More than 1,000 cells were counted in the control group. On day 7 of differentiation, oligodendrocytes were subjected to NG2 (green) and MBP immunostaining (red) to identify OPCs and mature oligodendrocytes, respectively. To stain the nuclei, the cells were incubated with 10 µg/mL 4′,6-diamidino-2-phenylindole (blue) for 5 minutes. The cells were mounted on glass slides with Immu-Mount (Thermo Fisher Scientific, Pittsburgh, PA, USA) after antibody application. The numbers of NG2+ and MBP+ cells relative to the total number of cells in the control, early LPS, late LPS, and double LPS groups were quantified.
Statistical analysis
All data are expressed as mean ± standard error of mean (n = 4–5/group). The means of different groups were analyzed using one-way analysis of variance analysis of variance with Bonferroni corrections or Mann-Whitney U testing (nonparametric) based on the homogeneity of variance testing (Levene’s test) using SPSS 25 statistical analysis software (SPSS Inc., Chicago, IL, USA). The differences were considered significant at P < 0.05.
Ethics statement
The procedures used and the care of animals were approved by the Institutional Animal Care and Use Committee in Hanyang University (approval No. 14-0050).
Go to :
RESULTS
To simulate the dose-dependent effect of LPS on oligodendrocytes and microglial cells, the effects of different doses of LPS on oligodendrocyte differentiation in vitro on day 3 were examined by subjecting the oligodendrocyte-microglia co-culture to immunostaining for MBP and IBA1. Representative images of oligodendrocytes, which were treated with different doses of LPS on day 3, on day 7 of differentiation are shown in Figs. 2 and 3. The number of MBP+ cells in the LPS (0.01 µg/mL)-treated group was non-significantly lower than that in the control group on day 7. LPS dose-dependently decreased the proportion of mature oligodendrocytes (MBP+ cells) relative to the total number of cells. The number of microglial cells in the LPS (0.1 and 1 µg/mL)-treated groups was significantly lower than in the control group (P < 0.001 and P = 0.007, respectively). However, the number of microglial cells in the 1 µg/mL LPS-treated group was significantly higher than that in the 0.1 µg/mL LPS-treated group (P = 0.009) (Fig. 2).
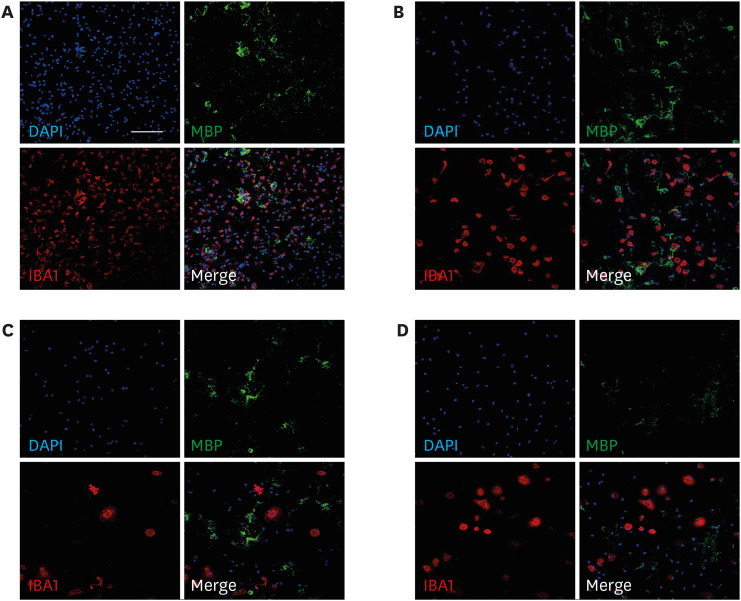 | Fig. 2Effect of various doses of LPS on oligodendrocyte differentiation. On day 3, the co-culture was treated with 0.01, 0.1, and 1 µg/mL LPS. On day 7 of differentiation, the nuclei were counterstained with 4′,6-diamidino-2-phenylindole (blue). The oligodendrocyte-microglia co-culture was subjected to MBP (green; mature oligodendrocyte marker) and IBA1 (red; microglia marker) immunostaining. The co-culture was treated with vehicle (control) (A) and 0.01 (B), 0.1 (C), and 1 µg/mL LPS (D). Scale bar = 100 µm (200 × magnification).LPS = lipopolysaccharide, MBP = myelin basic protein.
|
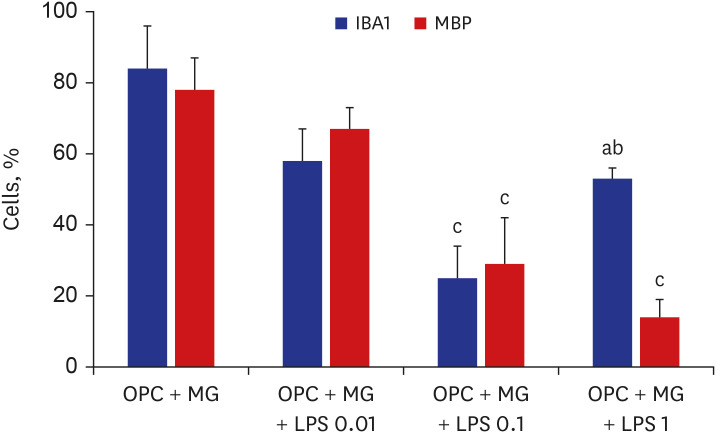 | Fig. 3Representative images of oligodendrocytes treated with various doses of LPS on day 7 of differentiation (n = 4/group).LPS = lipopolysaccharide, OPC = oligodendrocyte progenitor cell, MBP = myelin basic protein, MG = microglia.
P values from Mann-Whitney U test: a0.007 vs. control; b0.009 vs. OPC + MG + LPS 0.1. P values from one-way analysis of variance analysis of variance with Bonferroni: c< 0.001 vs. control.
|
Based on these results, 0.01 µg/mL LPS was used to simulate double-hit inflammation after preterm birth because this dose solely did not make significant effect on MBP+ and IBA1+ cell numbers. To develop the repetitive LPS exposure model for developing oligodendrocytes, the oligodendrocyte-microglia co-culture was treated with 0.01 µg/mL LPS at D1 (early LPS group), D3 (late LPS group), or D1 and D3 (double LPS group). The 0.01 µg/mL LPS in the late LPS group did not affect significantly both the MBP+ cell numbers and the NG2+ cell numbers. The number of MBP+ cells in the early LPS group was significantly lower than that in the control group and late LPS group (all P < 0.001). On day 7 of differentiation, compared with those in the control group and the late LPS group, the MBP+ cell numbers were significantly lower (all P < 0.001) and the NG2+ cell numbers were significantly higher (all P < 0.001) in the double LPS group (Figs. 4 and 5). The decreased MBP+ cell numbers and the increased NG2+ cell numbers were significant in the double LPS group compared to the early LPS group (P = 0.004 and P < 0.001, respectively).
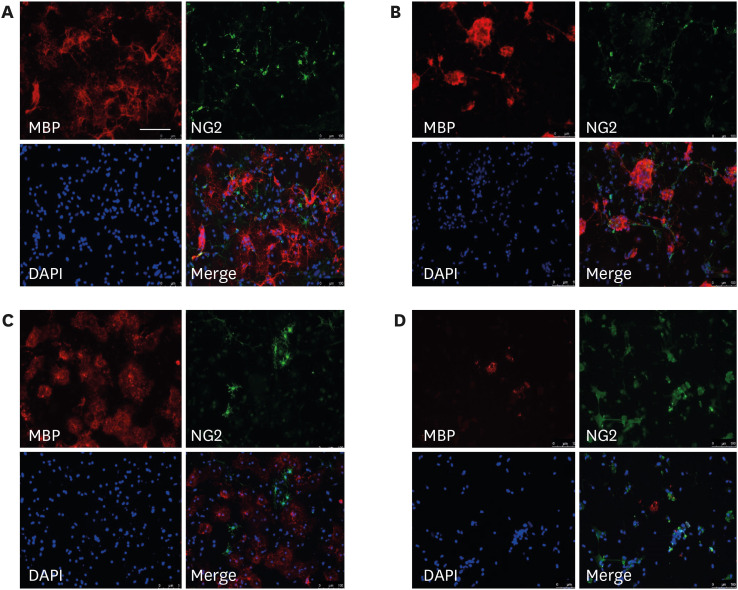 | Fig. 4Representative images of oligodendrocyte-microglia co-culture following different schedules of LPS treatment. Representative images of the oligodendrocyte-microglia co-culture subjected to immunostaining after in vitro treatment with vehicle (control) (A) or 0.01 µg/mL LPS at D1 (early LPS group) (B), D3 (late LPS group) (C), or D1 and D3 (double LPS group) stages. Scale bar = 100 µm (200 × magnification).LPS = lipopolysaccharide, MBP = myelin basic protein.
|
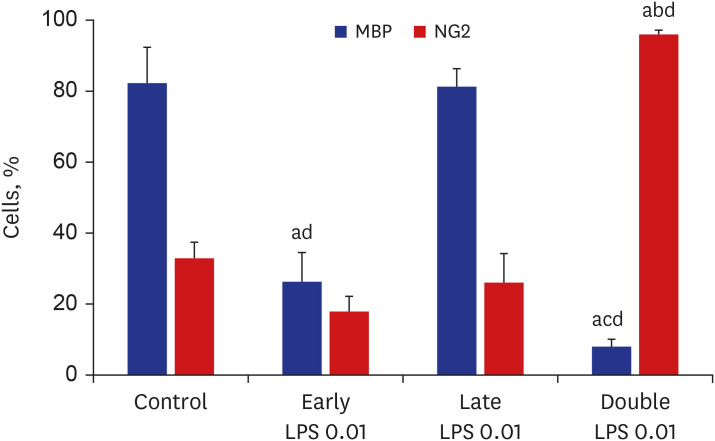 | Fig. 5Quantification of the relative numbers of NG2+ and MBP+ cells. Relative numbers of NG2+ and MBP+ cells were quantified after treatment with vehicle (control) or 0.01 µg/mL LPS at D1 (early LPS group), D3 (late LPS group), or D1 and D3 (double LPS group) stages. The number of MBP+ cells in the early LPS group was significantly lower than that in the control group. Compared with those in the control group, the MBP+ cell numbers were significantly lower and the NG2+ cell numbers were significantly higher in the double LPS group at day 7 of differentiation (n = 5/group).LPS = lipopolysaccharide, MBP = myelin basic protein.
All P values from one-way analysis of variance analysis of variance with Bonferroni; a< 0.001 vs. control; b< 0.001 vs. early LPS; c0.004 vs. early LPS; d< 0.001 vs. late LPS.
|
Go to :
DISCUSSION
To the best of our knowledge, this is the first study to demonstrate the effect of repeated LPS exposure on oligodendrocyte differentiation using the microglial cell and oligodendrocyte co-culture model. Some animal studies have demonstrated developmental stage-dependent susceptibility or LPS-hypoxic ischemia-induced brain injury. However, the effect of LPS exposure at the early and late developmental stages on the induction of OPC differentiation and oligodendrocyte injury in vitro has not been previously reported. Repetitive LPS stimulation during the developmental stage significantly inhibited brain cell development and consequently impaired oligodendrocyte differentiation. In contrast, single LPS exposure at the late developmental stage did not affect brain cell development. These findings suggest that inflammation at the early developmental stage of oligodendrocytes increases the susceptibility of the preterm brain to inflammation-induced injury.
In this study, an LPS-induced brain injury model proposed by Miller et al.11 was modified. In this model, 1 µg/mL of LPS exerted deleterious effects on OPC survival and proliferation. This study examined the response of oligodendrocytes to various concentrations of LPS in the microglial cell-oligodendrocyte co-culture model. At concentrations of 1 and 0.1 µg/mL, LPS significantly decreased the number of oligodendrocytes in the microglial cell-oligodendrocyte co-culture at the D3 stage. A similar effect was not observed upon treatment with 0.01 µg/mL LPS (Fig. 2). Additionally, LPS dose-dependently promoted cell death in oligodendrocytes, which was consistent with the results of a study by Lehnardt et al.19 The number of IBA1+ cells in the groups treated with 0.1 or 1 µg/mL LPS was lower than that in the control group. However, the number of IBA1+ cells in the groups treated with 1 µg/mL LPS was higher than that in the group treated with 0.1 µg/mL LPS. The enhanced number of microglial cells can be attributed to the LPS-induced microglial activation. Conflicting findings regarding the microglial responses to LPS stimulation have been reported. In addition to inducing cell death in oligodendrocytes and microglial cells, LPS can simultaneously stimulate microglial cells. The microglial response to LPS has been previously reviewed by Hoogland et al.20 LPS-induced activation of microglia is reported to increase the number and/or size of the microglial cells or upregulate the expression of a microglial marker. In contrast, Liu et al.21 reported that LPS concentration-dependently and time-dependently decreased the viability of microglia. Some studies have suggested that the overactivation of microglia adversely affects the survival of microglia.222324 It is possible that the inversed decreasing tendency of IBA1+ cell number with 1 µg/mL LPS is understood as the summation of the microglia cell death by LPS and the reactive proliferation of microglia to LPS stimulation.
The dose of LPS that did not affect the development of oligodendrocyte lineage in the microglial cell-oligodendrocyte co-culture model was 0.01 µg/mL, which was used for repeated stimulation.
The susceptibility of the brain white matter to systemic inflammation is dependent on the presence of pre-OLs at the gestational age of 23–32 weeks in humans. A similar pre-OL phenotype is reported in newborn rats.25 The effects of LPS exposure at D1 (early LPS group) and D3 (late LPS group) developmental stages, which correspond to 23–32 weeks of gestational age, on OPCs and oligodendrocyte injury were analyzed in vitro. LPS exposure at the D3 stage did not significantly affect the numbers of MBP+ and NG2+ cells. In contrast, LPS exposure at the D1 stage significantly decreased the number of MBP+ cells (Fig. 5). This suggests that OPCs and oligodendrocytes exhibit differential responses to LPS at different developmental stages and that these cells exhibit maturation-dependent susceptibility to LPS.9 Miller et al.11 reported that the oligodendrocytes exhibited differential responses to the activated microglial cells in vitro depending on the developmental stage. Activated microglial cells promote cell death of OPCs but simultaneously enhance survival and downregulate apoptosis in mature oligodendrocytes. This result may be because OPCs, which are susceptible to LPS-activated microglia, form the majority of cells in the co-culture at the D1 stage. At the D3 stage, a large proportion of OPCs can develop into mature oligodendrocytes, which are resistant to LPS-activated microglial cells. This explains the decreased number of MBP+ cells in the early LPS group and the preserved number of MBP+ cells in the late LPS group. However, this study did not confirm the accurate developmental stages of MBP+ cells at the D1 and D3 stages.
Another notable finding of our study is that LPS exposure at the early developmental stage (D1) exacerbated the detrimental effects of the subsequent LPS exposure (at D3) on OPCs and oligodendrocyte injury. Maternal/fetal infection is reported to contribute to periventricular leukomalacia, which further exacerbates inflammation-induced brain injury, in preterm infants. Moreover, repetitive exposure to inflammation/infection is associated with aberrant white matter maturation and impaired neurodevelopment.132627 Clinical studies have reported the correlation between prenatal inflammation and white matter injury or neurodevelopmental outcomes in preterm infants.1428293031 Preterm infants are at a high risk for developing pathological conditions, such as chorioamnionitis and subsequent sepsis, during the fetal and neonatal period. However, the effects of early inflammatory insults on OPC and the subsequent modulation of inflammation have not been previously reported using the microglial cell-oligodendrocyte co-culture model in vitro. The number of MBP+ cells in the double LPS group was lower than that in the early LPS group. However, the number of NG2+ cells in the double LPS group was significantly higher than that in the control group (Fig. 5). Two mechanisms explain the results of this study. The first mechanism is based on the susceptibility of OPCs to LPS insults. In the double LPS group, most cells are OPCs that are susceptible to LPS-induced inflammation at the D1 stage. The first (early) LPS stimulation promoted OPC damage and a minimal number of OPCs can mature into MBP+ oligodendrocytes. However, the surviving OPCs are more susceptible to the next inflammatory insult. Thus, the second LPS insult at D3 further decreases the number of MBP+ cells at the D7 stage. The second mechanism involves LPS delivered through infections or in vitro injection arresting the maturation of OPCs. Hence, OPCs may lose their ability to develop into MBP+ mature oligodendrocytes in the double LPS group.
Next, we focused on the number of NG2+ cells. Compared with that in the control group, the number of NG2+ cells were significantly higher in the double LPS group. NG2+ cells are mainly OPCs, whose numbers were intuitively expected to decrease in the double LPS group. However, the increased number of NG2+ cells after double LPS insult suggests an increase in the population of OPCs. Therefore, we hypothesized that the depletion of MBP+ cells was caused by the arrested development of OPCs, which may undergo fresh proliferation after multiple insults. The increased number of NG2+ cells in the double LPS group indicates that the NG2+ cells underwent developmental arrest at the OPC stage after repetitive LPS stimulation and continue to proliferation and that these NG2+ cells did not differentiate into mature oligodendrocytes. Consequently, the number of MBP+ cells remained low as new MBP+ cells did not develop from NG2+ cells. Similar phenomena have been observed in other studies. Segovia et al.32 reported that pre-OL degeneration induced by hypoxic-ischemia was offset by a robust regenerative response that resulted in a several-fold expansion in the pool of surviving pre-OLs in chronic lesions. Interestingly, these regenerated pre-OLs exhibited persistent maturation arrest and did not differentiate or generate myelin. Billiards et al.33 demonstrated that in contrast to mature oligodendrocytes, OPCs underwent proliferation to generate new oligodendrocytes compensatively in response to multiple insults based on white matter analysis in periventricular leukomalacia. Pang et al.7 reported that hypomyelination can be resulted from decreased number or dysfunction of mature oligodendrocytes. Various studies have demonstrated the development failure of oligodendrocyte lineage after insults, such as LPS or hypoxic-ischemia.3435
This study demonstrated that LPS dose-dependently decreased the viability of oligodendrocyte lineage cells using the oligodendrocyte-microglial co-culture system. Additionally, repetitive LPS stimulation adversely affected OPCs, which can be attributed to the vulnerability of OPCs to LPS insult and the LPS-induced developmental arrest of OPCs or pre-OLs. These findings provide useful insights into the development and severity of periventricular leukomalacia in some preterm infants and the potential targets for protecting vulnerable premature brains from damage and subsequent developmental impairment.
This study has several limitations. The morphology of activated microglial cells, which can dose-dependently exacerbate the oligodendrocyte injury, was not delineated in this study. Additionally, the correlation between the morphological changes in LPS-activated microglial cells and the response of OPCs was not determined. Furthermore, the exact numbers of OPCs or mature oligodendrocytes classified according to the morphology at D1 and D3 stages, which will enable the determination of the developmental stage of oligodendrocytes, were not determined.
The findings of this study highlight the potential correlation between susceptibility to neuroinflammation at different developmental stages and the underlying neurobiological processes in preterm infants with periventricular leukomalacia. Thus, understanding the triggered and synergistic effects of early exposure to inflammation, even at low doses, will improve our understanding of impaired white matter development in premature infants.
Go to :
 XML Download
XML Download