

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Psychomotor stimulants produce rewarding and locomotor activating effects by increasing dopaminergic neurotransmission in the nucleus accumbens (NAcc) [1-4]. In addition to classic signaling pathways that use cAMP, dopamine is also known to have a distinct signaling pathway involving glycogen synthase kinase 3β (GSK3β) [5,6], a serine/threonine kinase highly expressed in the brain [7]. GSK3β is inhibited by phosphorylation at serine-9 residue [8,9], and there is a negative correlation between its phosphorylation levels and dopamine- and psychomotor stimulant-induced locomotor activities [5,6]. For example, it was shown that an increase in GSK3β phosphorylation attenuated the expression of locomotor sensitization induced by psychomotor stimulants, including cocaine, amphetamine (AMPH), and meth-AMPH [10-13], whereas a decrease in GSK3β phosphorylation enhanced the increase in locomotor activity induced by acute cocaine administration [14]. Interestingly, the expression of locomotor sensitization induced by psychomotor stimulants selectively accompanied the decrease in GSK3β phosphorylation in the NAcc core [11,12,14], in two distinct sub-regions of the NAcc (the core and the shell) [15-17], suggesting that increased GSK3β activity in this region is necessary for psychomotor stimulants to produce locomotor sensitization.
In addition to sensitization, another important feature of drugs of abuse contributing to the development of craving and consequent relapse is their ability to confer rewarding effects to the environment associated with them [18-21]. Similar to its role in locomotor sensitization, GSK3β has also been implicated in mediating conditioned rewards formed by associations of psychomotor stimulants and the place that they were given [22-25]. For example, it was shown that the developments of conditioned place preference induced by cocaine and AMPH was prevented by GSK3β inhibitors in mice [22] and rats [24], respectively. However, although the NAcc is known as one of the critical neuronal substrates mediating conditioned behaviors [26,27], it remains undetermined which sub-region of the NAcc GSK3β contributes to mediating conditioned behaviors. Thus, in the present study, we measured the GSK3β phosphorylation levels separately in two sub-regions of the NAcc following the expression of AMPH-induced conditioned locomotor activity. Further, we examined whether conditioned behavior and the accompanying changes in GSK3β phosphorylation recovered following treatment with a GSK3β inhibitor.
METHODS
Subjects and drugs
Male Sprague–Dawley rats (6 weeks old) weighing 220–250 g on arrival were obtained from Orient Bio Inc. (Seongnam, Korea). They were housed three per cage in a 12-h light/dark cycle room (lights out at 8:00 pm), and all experiments were conducted during the day. Rats had access to water and food ad libitum at all times. All animal use procedures were conducted according to an approved Institutional Animal Care and Use Committee protocol (approval number: 09-140) of Yonsei University College of Medicine.
D-amphetamine sulfate (United States Pharmacopeia, Rockville, MD, USA) and lithium chloride (Sigma-Aldrich, St. Louis, MO, USA) were dissolved in sterile 0.9% saline.
Locomotor activity
Locomotor activity was measured using a bank of 9 activity boxes (35 × 25 × 40 cm) (IWOO Scientific Corporation, Seoul, Korea) made of translucent Plexiglas. Each box was individually housed in a PVC plastic sound attenuating cubicle. The floor of each box consisted of 21 stainless steel rods (5 mm diameter), spaced 1.2 cm apart, center-to-center. Two infrared light photo beams (Med Associates, St. Albans, VT, USA) were positioned 4.5 cm above the floor and spaced evenly along the longitudinal axis of the box, to estimate horizontal locomotion. A single locomotor activity was only considered to be legitimate when a rat interrupted two beams consecutively, so as to avoid any possible confounding activities, such as grooming in a spot covering just a single beam.
Design and procedures
Upon arrival, all rats passed a week-long adaptation period to the new housing environment before conducting any experiments.
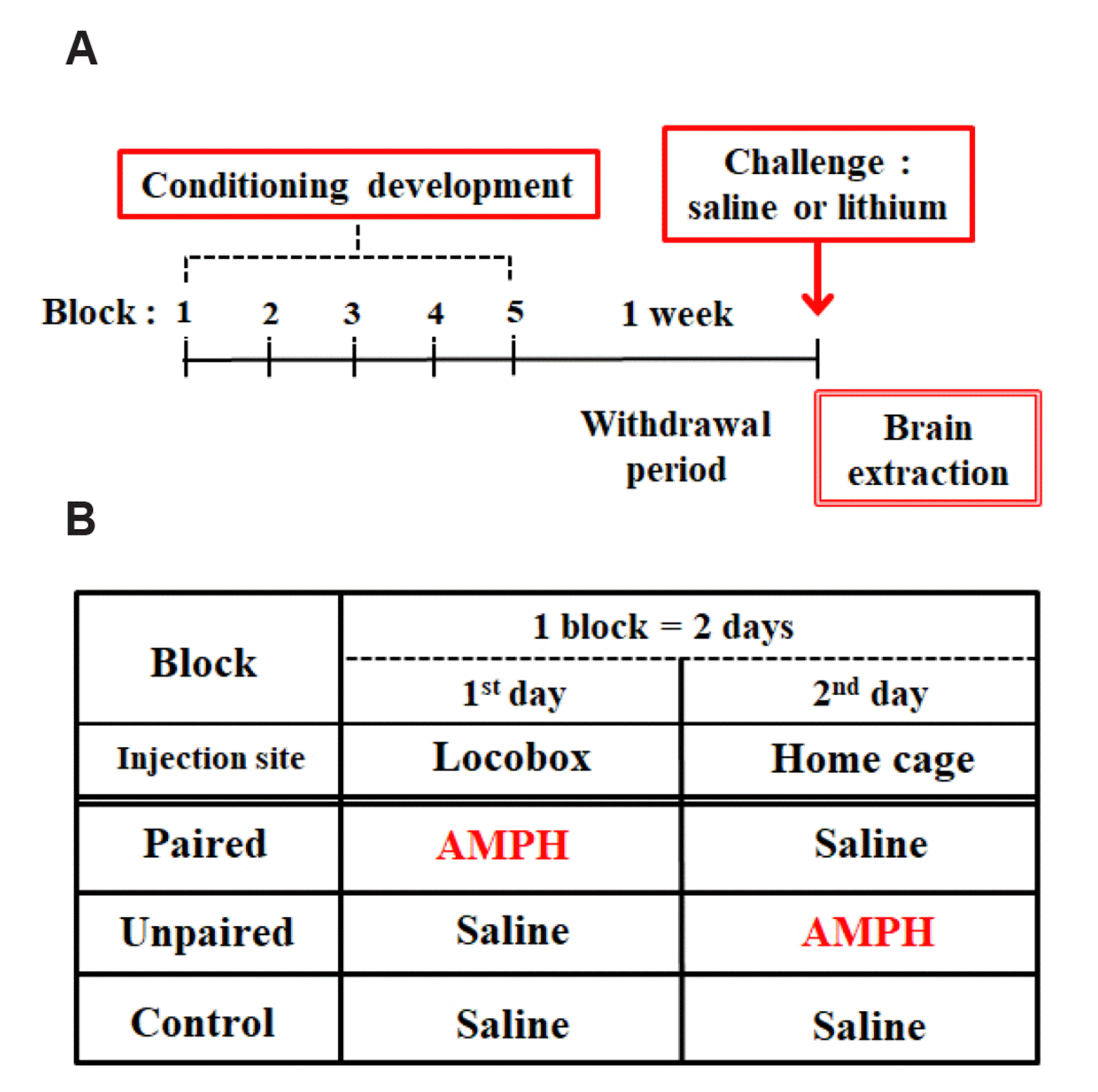
In Experiment 1, rats were randomly assigned to three groups: Paired, Unpaired, and Control. During conditioning, rats in these three groups were administered injections in five 2-day blocks. Rats received intraperitoneal injection (IP) of AMPH (1 mg/kg) in locomotor activity boxes on day 1 and saline in their home cages on day 2 (Paired), saline in the activity boxes on day 1 and AMPH in their home cages on day 2 (Unpaired), or saline in both environments (Control). This procedure that we used here has previously been shown to produce conditioned locomotor activity [27,28]. On the test, one week after the last conditioning block, all rats were tested for their conditioned locomotor response in the activity boxes for one hour following an IP saline injection.
In Experiment 2, rats were randomly assigned to three groups: Paired, Unpaired, and Control, as defined in Experiment 1. They followed the same procedure as described in Experiment 1 for the development of conditioning. On the test, one week after the last conditioning block, each group of rats were sub-divided into two. Then, they were tested for their conditioned locomotor response in the activity boxes for one hour following an IP saline or lithium chloride (50 mg/kg) injection.
Brain tissue preparation
Immediately after completion of behavioral experiments, rats were decapitated by guillotine. Brains were rapidly removed and coronal sections (1.0 mm thick extending 1.60–2.60 mm from bregma) were obtained with an ice-cold brain slicer (Model SA-2160; Roboz Surgical Instrument Co., Gaithersburg, MD, USA). The NAcc core (circular punch with 1.0 mm diameter) and the shell (rectangular shape cut) regions were obtained on an ice-cold plate. Tissues were all immediately frozen on dry ice and stored at –80°C. They were prepared bilaterally and pooled for each individual animal’s protein isolation.
Western blotting
Tissues were homogenized in lysis buffer containing 0.32 M sucrose, 2 mM EDTA, 1% SDS, 10 µg/ml aprotinin, 10 µg/ml leupeptin, 1mM phenylmethylsulfonyl fluoride, 10 mM sodium fluoride, and 1 mM sodium orthovanadate. The concentration of protein was determined by using Pierce Coomassie Protein Assay Kit (Thermo Scientific Inc., Rockford, IL, USA). Samples were then boiled for 10 min and subjected to SDS-polyacrylamide gel electrophoresis. Proteins were separated and transferred electrophoretically to nitrocellulose membranes (Bio-Rad, Hercules, CA, USA), which were then blocked with 5% bovine serum albumin (BSA) in PBS-T buffer (10 mM phosphate-buffered saline plus 0.05 % Tween-20). Antibodies used to probe the blots were as following: total GSK3β (1:4,000), phosphor-GSK3β (specific to detect phosphorylated GSK3β at serine 9, 1:2,000), purchased from Cell Signaling (Beverly, MA, USA) and diluted in PBS-T with 5% BSA; β-actin (1:10,000), purchased from Abcam (Cambridge, UK) and diluted in PBS-T with 5% skim milk. Two separate gels were used to detect total and phosphorylated proteins, respectively. Primary antibodies were detected with peroxidase-conjugated secondary antibodies, anti-rabbit IgG (1:2,000; KOMA Biotech, Seoul, Korea) diluted in PBS-T with 5% skim milk, followed by enhanced chemiluminescence reagents (Amersham Biosciences, Arlington Heights, IL, USA) and exposure to X-ray film. Band intensities were quantified based on densitometric values using Fujifilm Science Lab 97 Image Gauge software (version 2.54) (Fujifilm, Tokyo, Japan).
Statistical analyses
Statistical analyses were performed using Sigma Plot version 12.0 (Systat Software, San Jose, CA, USA). The data were analyzed with one-way or two-way analysis of variance (ANOVA) followed by post-hoc Bonferroni comparisons. Differences between experimental conditions were considered statistically significant at p < 0.05.
RESULTS
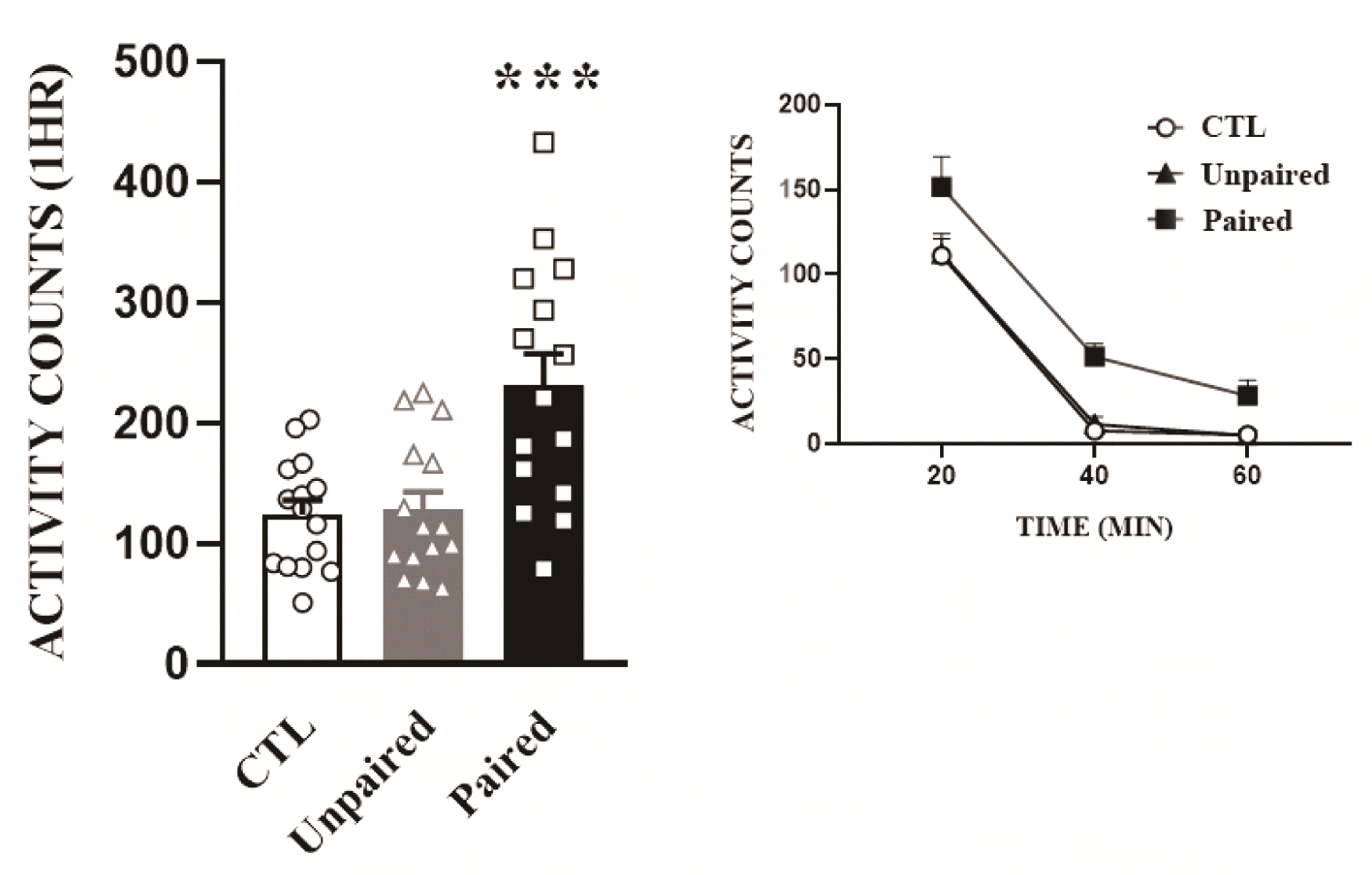
Repeated pairing of AMPH to a locomotor activity box produces conditioned locomotor activity
One week after the last block of the conditioning development schedule, all rats were saline-challenged and locomotor activities were measured (Fig. 2). One-way ANOVA conducted on these data revealed a significant effect of pairing (F2,42 = 10.67, p < 0.001). As expected, rats paired with AMPH in the locomotor activity boxes (Paired) showed higher locomotor responses than those in the saline group (Control). Further, the Paired group also exhibited higher locomotor activities when compared to rats administered the same amount of AMPH in different environments (Unpaired) (p < 0.001 by post-hoc Bonferroni comparison).
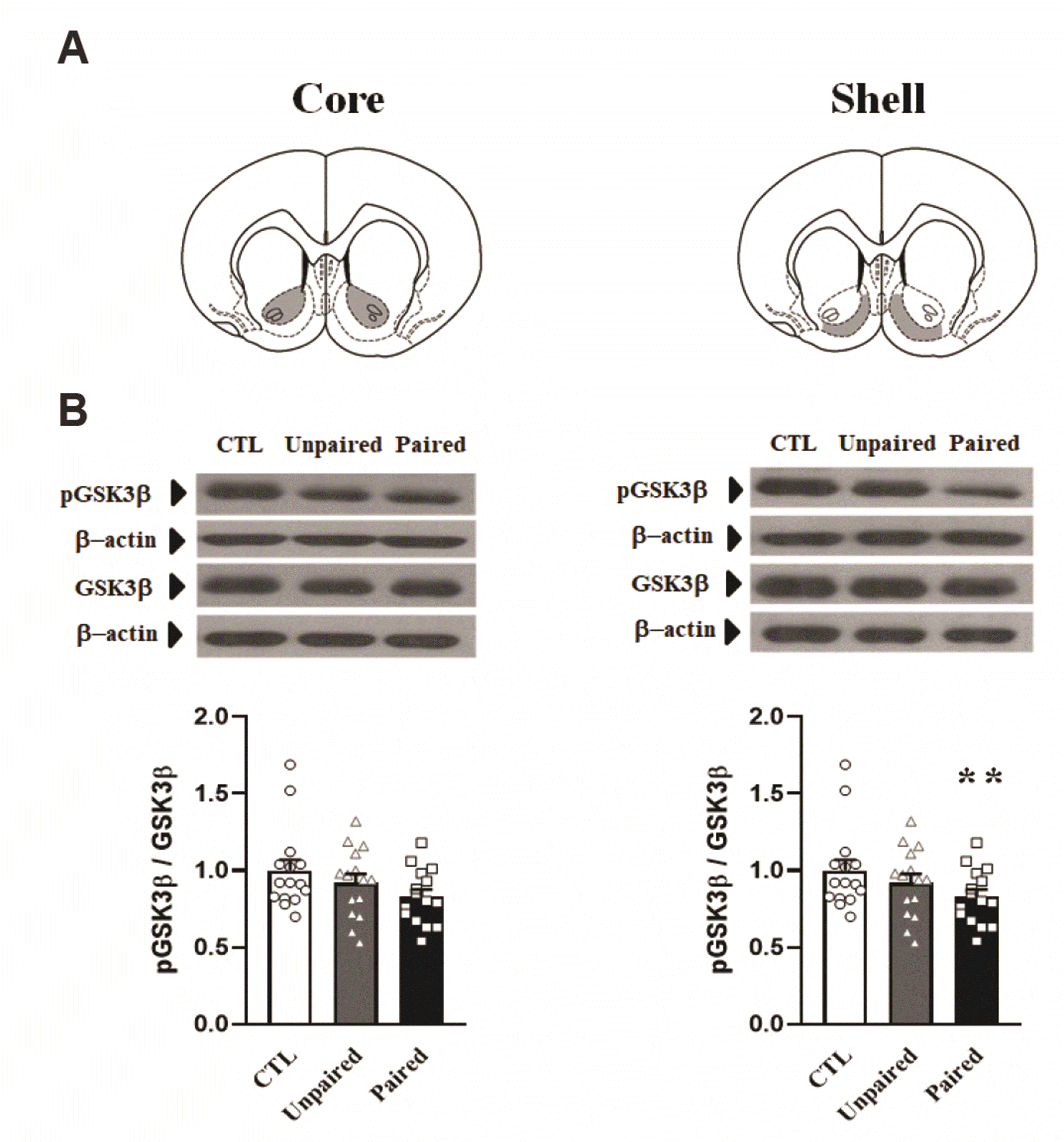
AMPH conditioning decreases GSK3β phosphorylation levels in the NAcc shell
To test whether the development of AMPH-induced conditioning may regulate GSK3β phosphorylation levels in the NAcc, the ratio of phosphorylated to total GSK3β was examined using Western blotting of samples of two sub-regions of the NAcc obtained 60 min after saline challenge (Fig. 3). The one-way ANOVA conducted on the data obtained from the core showed no significant effect of pairing (F2,42 = 2.09, p = 0.136). However, the one-way ANOVA conducted on the data obtained from the shell revealed significant effect of pairing (F2,42 = 5.28, p < 0.01). Post-hoc Bonferroni comparisons revealed that the ratio of phosphorylated to total GSK3β in the shell was significantly lower (p < 0.01) in the Paired group than in the Control group.
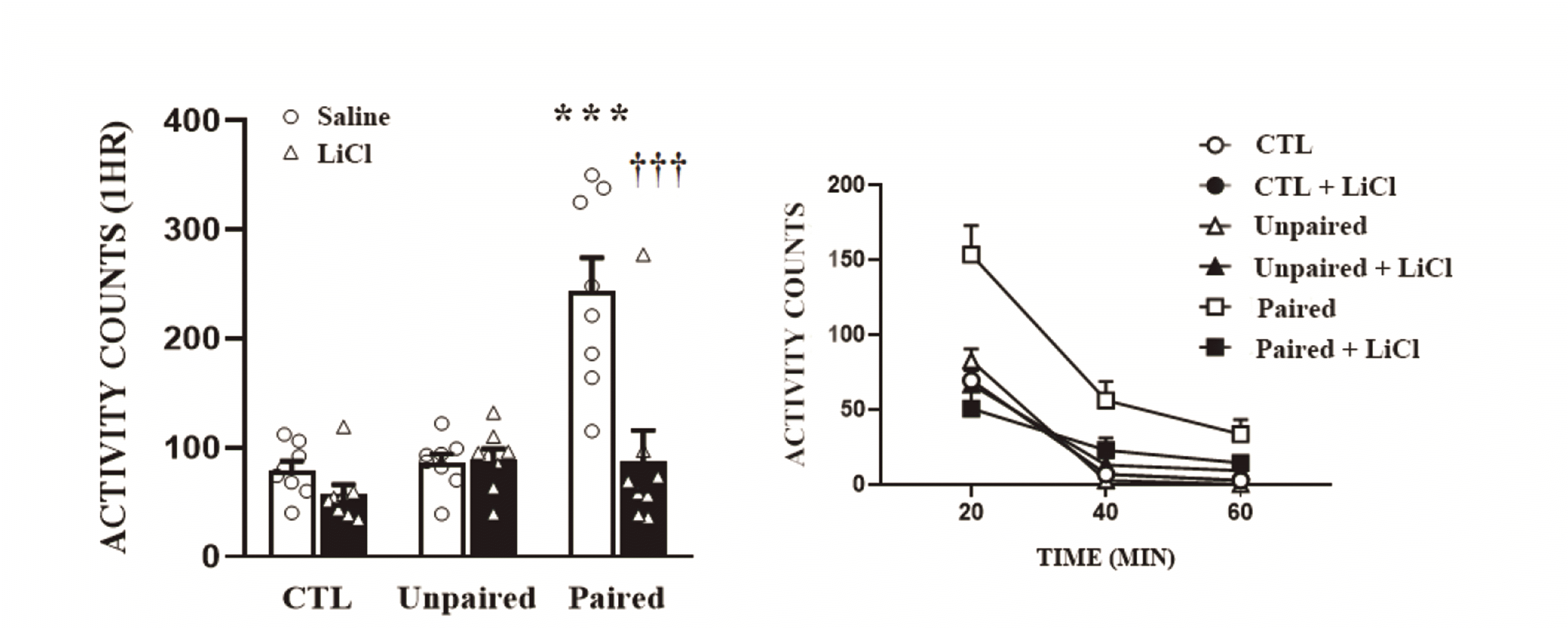
Lithium chloride inhibits the expression of AMPH-induced conditioned locomotor activity
In Experiment 2, after the conditioning development, we challenged half of the rats in each group with saline and the other half with lithium chloride, and their locomotor activities were measured (Fig. 4). The two-way ANOVA conducted on these data revealed multiple significant effects of pairing (F2,42 = 15.53, p < 0.001), lithium chloride (F1,42 = 14.71, p < 0.001), and an interaction of pairing and lithium chloride (F2,42 = 10.53, p < 0.001). Again, the Paired group showed higher locomotor responses than the other groups (Control, Unpaired). However, these effects were not observed when challenged with lithium chloride (p < 0.001 by post-hoc Bonferroni comparison).
Lithium chloride recovers the reduction in GSK3β phosphorylation levels in the NAcc
Next, we measured the ratio of phosphorylated to total GSK3β levels in the NAcc at 60 min after saline or lithium chloride challenge (Fig. 5). There were no significant effects observed on the data obtained from the core, whereas the two-way ANOVA conducted on data obtained from the shell revealed a significant effect of lithium chloride (F1,42 = 6.23, p < 0.02). Again, the decrease in phosphorylation levels of GSK3β in the shell was only observed in the Paired group (p < 0.05, post-hoc Bonferroni comparison). Interestingly, however, these effects were absent with lithium chloride challenge in the Paired group (Fig. 5). Post-hoc Bonferroni comparisons revealed that rats in the Paired group with lithium chloride challenge showed a significant recovery of GSK3β phosphorylation levels close to Control levels in the shell (p < 0.01).
DISCUSSION
The present results revealed that the expression of AMPH-induced conditioned locomotor activity accompanied a significant decrease in GSK3β phosphorylation in the NAcc shell. Furthermore, our results revealed that these effects were reversed by lithium chloride, a GSK3β inhibitor.
It was previously shown that GSK3β phosphorylation in the NAcc was decreased when cocaine-associated contextual memory was reactivated [23], while knockdown of GSK3β in the NAcc attenuated the development of cocaine-induced place preference [25]. These results indicate that intact GSK3β activity in the NAcc is necessary for the development and expression of psychomotor stimulants-induced conditioned responses. Consistent with these results, we showed in the present results that GSK3β phosphorylation was lower in the NAcc following the expression of AMPH-induced conditioned locomotor activity. Furthermore, contrary to previous results that did not distinguish the role of GSK3β in the two sub-regions of the NAcc, we found that GSK3β phosphorylation was more significantly reduced in the shell than in the core (Fig. 3). It has been shown in the literature that the NAcc shell plays a more important role in the processing of contextual information than that of the core, and thereby contributes to the context-induced reinstatement of drugs of abuse [29-34]. The increased conditioned locomotor activity toward contextual stimuli associated with drugs of abuse, shown in our present results, can be considered as a manifestation of drug seeking behavior [35]. Accordingly, the finding that a significant decrease in GSK3β phosphorylation in the shell was only observed in the Paired but not in the Unpaired group, which received the same amount of AMPH in a different place, imply that they may reflect a specific role of GSK3β signaling in mediating conditioning effect of AMPH-associated environmental stimuli.
It was previously shown that the development of conditioned place preference induced by cocaine or AMPH was prevented by either systemic or intra-accumbal injection of a GSK3β inhibitor [22-24]. Consistent with these results, we showed that lithium chloride, a GSK3β inhibitor, completely abolished the expression of conditioned locomotor activity (Fig. 4). In addition, we also demonstrated that the decrease in GSK3β phosphorylation in a drug-associated conditioning model was normalized back to control level by lithium chloride treatment following behavioral recovery (Fig. 5). As lithium chloride is known to have the ability to increase the phosphorylation levels of Akt, an up-stream effector of GSK3β, and thereby its phosphorylation levels [36,37], our present findings suggest that GSK3β signaling in the NAcc shell may be under the general regulation of the well-known phosphatidylinositol 3-kinase (PI3 kinase) and Akt signaling pathways. Looking into downstream components, it is not yet known what signaling molecules are involved in the expression of AMPH-induced conditioned locomotor activity. Interestingly, in rats associated with AMPH, dendritic spine density was increased in the shell only in Paired rat [35]. Further, it was recently shown that ezrin-radixin-moesin (ERM) proteins, a group of proteins which play a role in cell-shape determination and spine maturation, are under the regulation of Akt-GSK3β signaling pathways [38], suggesting that there is a possibility for GSK3β to become involved in spine formation. Thus, it will be interesting to examine in the future how GSK3β is involved in these process to eventually contribute to the expression of AMPH-induced conditioned locomotion.
 XML Download
XML Download