

 PDF
PDF Citation
Citation Print
Print



Introduction
Materials and Methods
Analysis of Public Repository Data
Immunohistochemical (IHC) Staining
Cell Culture and Reagents
Short Interfering RNA (siRNA) Transfection
RNA Isolation and Real-Time PCR
Statistical Analysis
Results
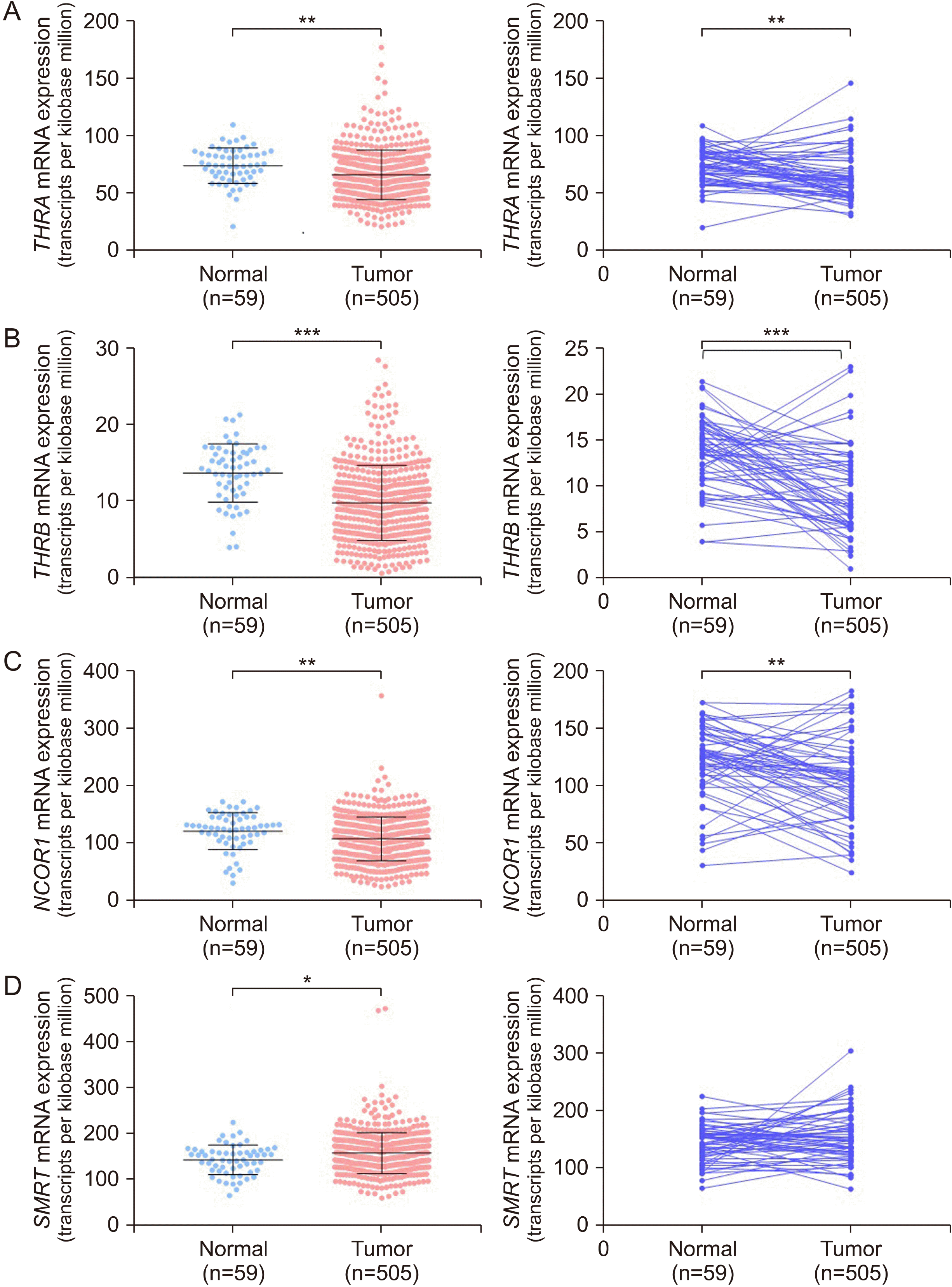
Cooperative Subtype Switch of THR and NCOR1/SMRT Expression in Papillary Thyroid Cancer
Fig. 1
![]()
Fig. 2
![]()
Clinicopathological Characteristics according to THRA, THRB, and NCOR1/SMRT Expression in Papillary Thyroid Cancer
Table 1
| Characteristics | THRA | p-value | |
|---|---|---|---|
|
|
|||
| Low expression n=126 (%) | High expression n=126 (%) | ||
| NCOR1 expression | 112.33±35.59 | 103.92±43.00 | 0.092* |
| SMRT expression | 167.14±41.05 | 153.82±40.66 | 0.010* |
| Age (years) | 46.64±17.22 | 49.56±15.84 | 0.180* |
| Gender (Male:Female) | 34 (27.0):92 (73.0) | 34 (27.0):92 (73.0) | 0.999† |
| Tumor size (cm) | 3.02±1.64 | 2.99±1.75 | 0.960* |
| MACIS score | 5.59±1.66 | 5.30±1.38 | 0.173* |
| Histologic subtype | <0.0001† | ||
| Follicular variant | 5 (4.0) | 37 (30.3) | |
| Conventional | 100 (80.6) | 80 (65.6) | |
| Tall cell variant | 18 (14.5) | 2 (1.6) | |
| Othersς | 1 (0.8) | 3 (2.5) | |
| Extrathyroidal extension | <0.0001† | ||
| No | 65 (57.5) | 89 (80.2) | |
| Yes | 48 (42.5) | 22 (19.8) | |
| Multifocality | 0.048† | ||
| No | 73 (58.9) | 55 (46.2) | |
| Yes | 51 (41.1) | 64 (53.8) | |
| T stage | 0.013† | ||
| T1 | 27 (23.3) | 38 (32.8) | |
| T2 | 33 (28.4) | 41 (35.3) | |
| T3 | 48 (41.4) | 33 (28.4) | |
| T4 | 8 (6.9) | 4 (3.4) | |
| N stage | <0.0001† | ||
| N0 | 36 (33.0) | 64 (64.6) | |
| N1 | 73 (67.0) | 35 (35.4) | |
| M stage | 0.480† | ||
| M0 | 69 (98.6) | 59 (96.7) | |
| M1 | 1 (1.4) | 2 (3.3) | |
| TNM stage | 0.127† | ||
| I | 63 (53.8) | 62 (54.4) | |
| II | 5 (4.3) | 20 (17.5) | |
| III | 33 (28.2) | 25 (21.9) | |
| IV | 16 (13.7) | 7 (6.1) | |
NCOR1: nuclear receptor co-repressor 1, MACIS: distant metastasis, patient age, completeness of resection, local invasion, and tumor size, SMRT: silencing mediator for retinoid or thyroid hormone receptor, SD: standard deviation, TCGA: The Cancer Genome Atlas, THCA: thyroid cancer, THRA: thyroid hormone receptor alpha, TNM: tumor-node- metastasis
![]()
Table 2
| Characteristics | THRB | p-value | |
|---|---|---|---|
|
|
|||
| Low expression n=126 (%) | High expression n=126 (%) | ||
| NCOR1 expression | 94.22±37.54 | 120.88±34.07 | <0.0001* |
| SMRT expression | 141.19±38.16 | 176.54±49.18 | <0.0001* |
| Age (years) | 43.62±13.84 | 49.29±15.88 | 0.004* |
| Gender (Male:Female) | 28 (22.2):98 (77.8) | 25 (19.8):101 (80.2) | 0.643† |
| Tumor size (cm) | 3.00±1.64 | 2.97±1.61 | 0.893* |
| MACIS score | 5.01±1.42 | 5.64±1.65 | 0.003* |
| Histologic subtype | 0.047† | ||
| Follicular variant | 29 (23.0) | 20 (16.0) | |
| Conventional | 92 (73.0) | 90 (72.0) | |
| Tall cell variant | 3 (2.4) | 15 (12.0) | |
| Othersς | 2 (1.6) | 0 (0.0) | |
| Extrathyroidal extension | 0.005† | ||
| No | 92 (78.6) | 69 (61.6) | |
| Yes | 25 (21.4) | 43 (38.4) | |
| Multifocality | 0.276† | ||
| No | 66 (54.1) | 75 (61.0) | |
| Yes | 56 (45.9) | 48 (39.0) | |
| T stage | 0.129† | ||
| T1 | 35 (29.4) | 32 (27.4) | |
| T2 | 49 (41.2) | 37 (31.6) | |
| T3 | 32 (26.9) | 41 (35.0) | |
| T4 | 3 (2.5) | 7 (6.0) | |
| N stage | 0.065† | ||
| N0 | 63 (57.8) | 46 (45.1) | |
| N1 | 46 (42.2) | 56 (54.9) | |
| M stage | 0.941† | ||
| M0 | 63 (95.5) | 67 (95.7) | |
| M1 | 3 (4.5) | 3 (4.3) | |
| TNM stage | 0.040† | ||
| I | 74 (62.2) | 56 (47.9) | |
| II | 11 (9.2) | 13 (11.1) | |
| III | 23 (19.3) | 34 (29.1) | |
| IV | 11 (9.2) | 14 (12.0) | |
NCOR1: nuclear receptor co-repressor 1, MACIS: distant metastasis, patient age, completeness of resection, local invasion, and tumor size, SMRT: silencing mediator for retinoid or thyroid hormone receptor, SD: standard deviation, TCGA: The Cancer Genome Atlas, THCA: thyroid cancer, THRB: thyroid hormone receptor beta, TNM: tumor-node- metastasis
![]()
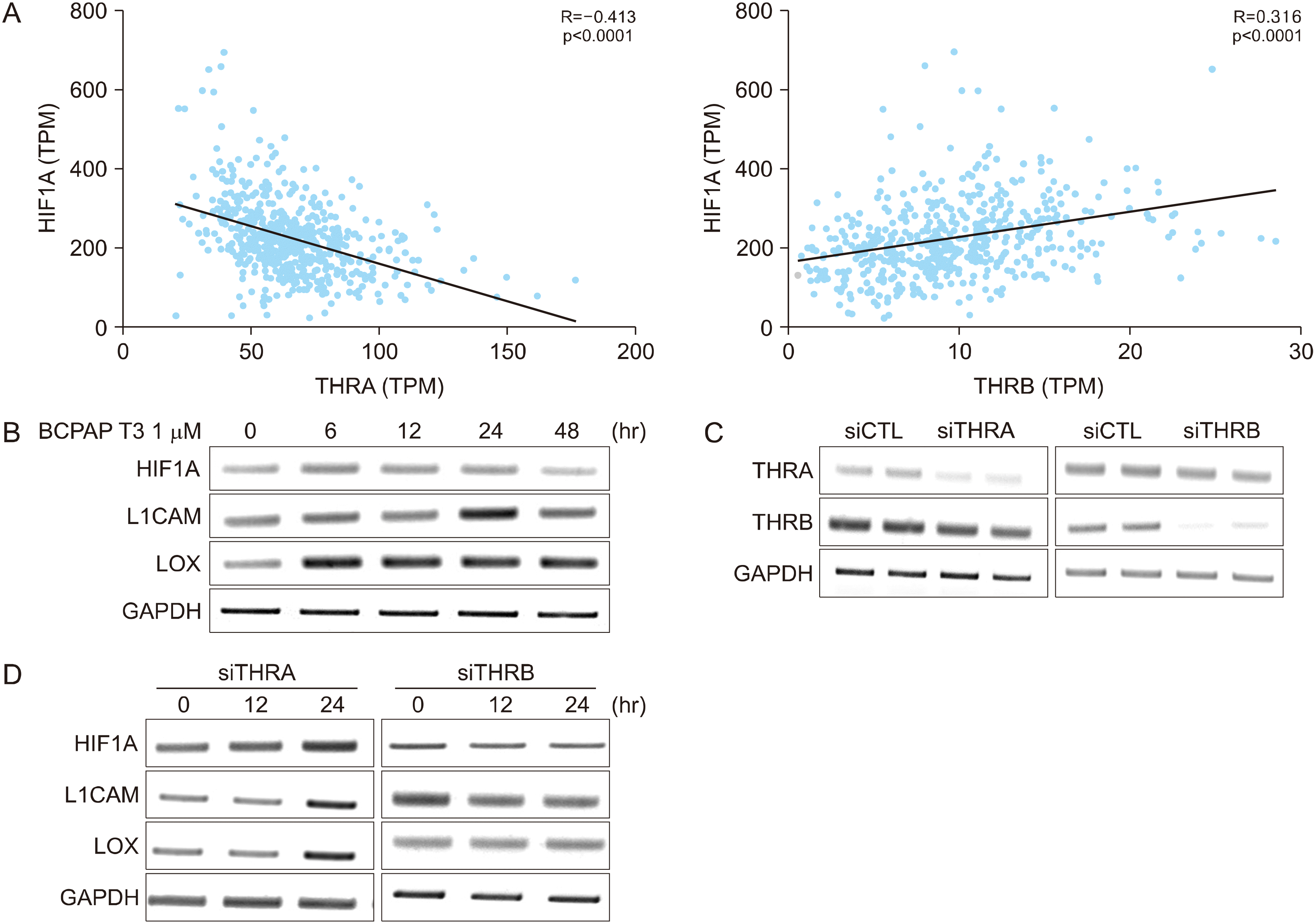
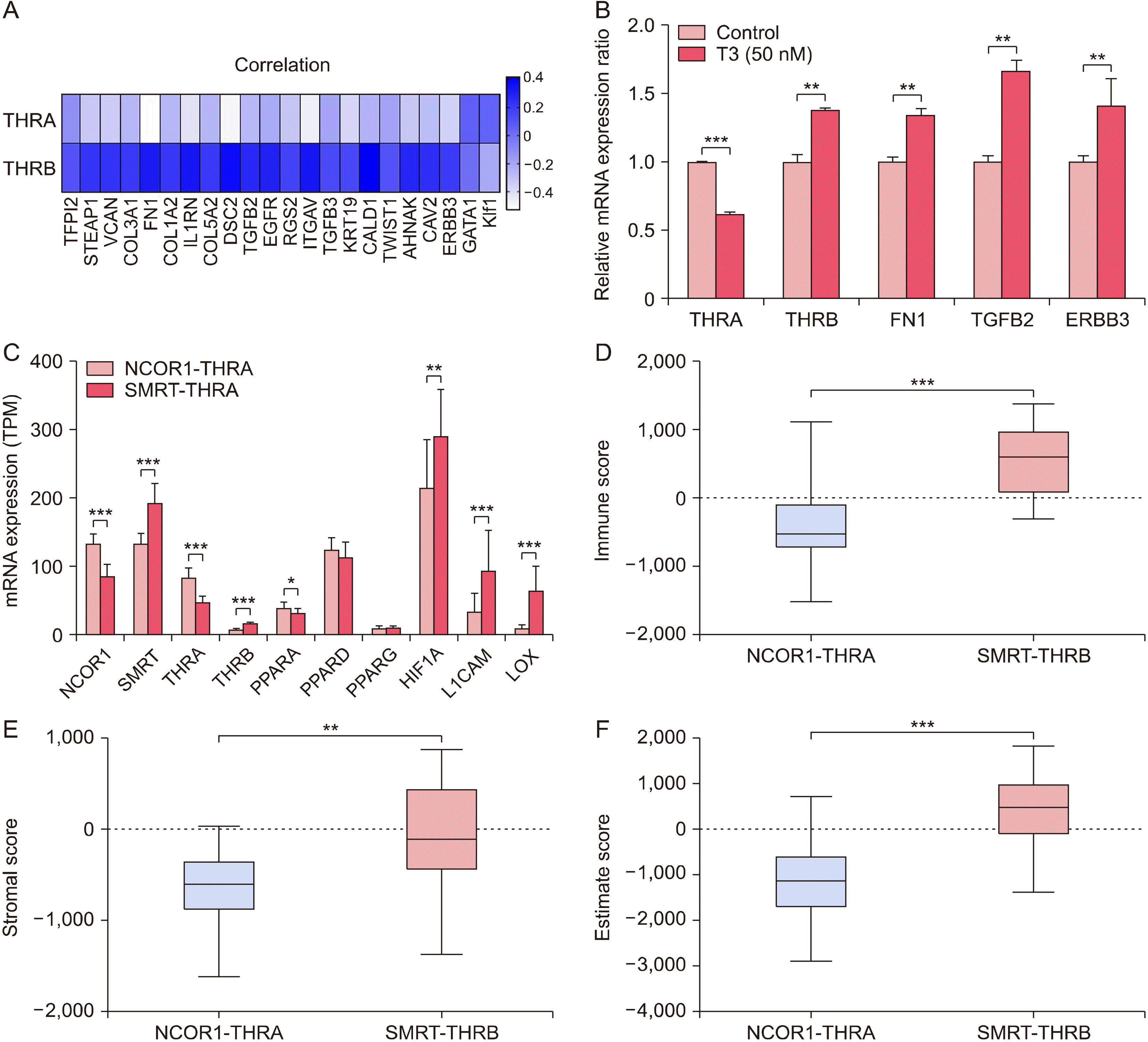
Subtype Change of TR from THRA to THRB is Linked with Epithelial to Mesenchymal Transition
Fig. 3
![]()
Fig. 4
![]()
Clinicopathological and Molecular Characteristics of the Cooperative Subtype Switch from NCOR1/THRA to SMRT/THRB
Table 3
| Characteristics | NCOR1/THRA n=24 (%) | SMRT/THRB n=24 (%) | p-value |
|---|---|---|---|
| Age (years) | 48.36±14.56 | 52.78±16.98 | 0.356* |
| Gender (Male:Female) | 3 (12.5):21 (87.5) | 10 (41.7):14 (58.3) | 0.023† |
| Tumor size (cm) | 2.68±1.50 | 3.02±1.38 | 0.475* |
| MACIS score | 5.16±1.24 | 6.05±2.01 | 0.097* |
| Histologic subtype | 0.172† | ||
| Follicular variant | 4 (18.2) | 1 (4.3) | |
| Conventional | 17 (77.3) | 18 (78.3) | |
| Tall cell variant | 0 (0.0) | 4 (17.4) | |
| Othersς | 1 (4.5) | 0 (0.0) | |
| Extrathyroidal extension | 0.028† | ||
| No | 19 (82.6) | 12 (52.2) | |
| Yes | 4 (17.4) | 11 (47.8) | |
| Multifocality | 0.555† | ||
| No | 11 (47.8) | 13 (56.5) | |
| Yes | 12 (52.2) | 10 (43.5) | |
| T stage | 0.038† | ||
| T1 | 9 (39.1) | 3 (13.0) | |
| T2 | 7 (30.4) | 7 (30.4) | |
| T3 | 6 (26.1) | 11 (47.8) | |
| T4 | 1 (4.3) | 2 (8.7) | |
| N stage | 0.001† | ||
| N0 | 17 (81.0) | 7 (31.8) | |
| N1 | 4 (19.0) | 15 (68.2) | |
| M stage | 0.309† | ||
| M0 | 14 (100.0) | 13 (92.9) | |
| M1 | 0 (0.0) | 1 (7.1) | |
| TNM stage | 0.038† | ||
| I | 14 (63.6) | 9 (39.1) | |
| II | 3 (13.6) | 2 (8.7) | |
| III | 4 (18.2) | 7 (30.4) | |
| IV | 1 (4.5) | 5 (21.7) |
NCOR1: nuclear receptor co-repressor 1, MACIS: distant metastasis, patient age, completeness of resection, local invasion, and tumor size, SMRT: silencing mediator for retinoid or thyroid hormone receptor, SD: standard deviation, TCGA: The Cancer Genome Atlas, THCA: thyroid cancer, THRA: thyroid hormone receptor alpha, THRB: thyroid hormone receptor beta, TNM: tumor-node-metastasis
![]()
Table 4
| Characteristics | NCOR1/THRA n=24 (%) | SMRT/THRB n=24 (%) | p-value |
|---|---|---|---|
| Ras driver mutation | 0.018† | ||
| Absent | 18 (78.3) | 23 (100.0) | |
| Present | 5 (21.7) | 0 (0.0) | |
| BRAF driver mutation | 0.376† | ||
| Absent | 13 (56.5) | 10 (43.5) | |
| Present | 10 (43.5) | 13 (56.5) | |
| TERT promoter mutation | 0.086† | ||
| Absent | 16 (94.1) | 13 (72.2) | |
| Present | 1 (5.9) | 5 (27.8) | |
| RAS/RAF score | −0.024±0.71 | −0.78±0.23 | <0.0001* |
| ERK score | −2.08±18.29 | 24.25±14.04 | <0.0001* |
| Differentiation score | 0.37±1.08 | −0.87±1.01 | <0.0001* |
| Tumor status | 0.130† | ||
| Free | 23 (100.0) | 19 (90.5) | |
| With tumor | 0 (0.0) | 2 (9.5) |
![]()
 XML Download
XML Download