

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Premenstrual syndrome (PMS) is a common cyclic disorder, in which psychological and/or physical symptoms occur in the luteal phase of the menstrual cycle. The American College of Obstetricians and Gynecologists published clinically significant diagnostic criteria for PMS in 2000. According to the criteria, PMS is diagnosed when at least one of the emotional symptoms (depression, angry outbursts, irritability, or confusion) and one or more of the physical symptoms (breast tenderness, abdominal bloating, headache, or swelling of extremities) appear 5 days before the onset of menstruation [1]. About 1.1–6.4% of women of childbearing age have more severe symptoms of PMS, including poor social coordination, which the American Psychiatric Association terms premenstrual dysphoric disorder [2]. It is not clear which particular sex steroid changes cause PMS symptoms in the luteal phase; however, hormonal changes, neurotransmitters, prostaglandins (PGs), prolactin secretion, genetic factors, diets, drugs and the lifestyle of women have all been implicated as contributing factors [345]. Recent studies have shown that changes in circulating estradiol and progesterone concentrations are associated with a variety of PMS symptoms [67]. Abnormal fluctuations in the circulating sex hormones during the menstrual cycle may also be associated with some of the physical and emotional symptoms [8].
Prolactin plays an essential role in the regulation of the immune system, pancreatic development, and metabolism [9]. Prolactin is produced in the lactotropic cells of the anterior pituitary gland and is secreted in a pulsatile fashion. Prolactin release from the pituitary gland is maintained by tonic hypothalamic inhibition and secretion is controlled by dopamine receptor D2 (DRD2), which is found in lactotropic cells [10]. Prolactin has a wide range of effects in the body: the focus of this study was the neuropeptide activities of prolactin to induce mammary gland enlargement during pregnancy and milk secretion after delivery, and in stress adaptation and neuroprotection [11]. Prolactin secretion increases during pregnancy, lactation, exercise, eating, sexual intercourse, and acute stress [12]. Increased prolactin induces PMS symptoms [13]: hyperprolactinemia induces premenstrual breast pain, weight gain, metabolic alterations, ovulation disorders and often leads to abnormal immune responses [1014]. Excessive prolactin secretion also causes inflammatory reactions. The prolactin receptor is a member of the type 1 cytokine/hematopoietic receptor superfamily and is widely expressed throughout the immune system [15]. Macrophages incubated with prolactin secrete greater amounts of interleukin (IL)-1β and IL-12, and of chemokines like the macrophage inflammatory protein. These immune and inflammatory changes promote PG release and the intense pain associated with uterine cramps; PGs may also be responsible for the vomiting, diarrhea, and headaches that frequently accompany painful periods as additional symptoms of PMS [15].
On the other hand, reduced prolactin secretion plays an important role in the suppression of PMS symptoms. In the previous studies, Hordeum vulgare L. showed anti-hyperprolactinemia activity and increased DRD2 expression [1617]. Chrysanthemum zawadskii var. latilobum has been used in traditional medicine for the treatment of various diseases, including pneumonia, inflammation, bronchitis, pharyngitis, rheumatoid arthritis, and gynecological disorders [1819]. In our previous study, to search for new natural substances to inhibit PMS symptoms, 61 herbal medicines extracts were screened for inhibition of prolactin secretion and induction of DRD2 expression. Among them, H. vulgare extract and C. zawadskii extract was confirmed to help inhibit prolactin secretion and induce DRD2 expression. Using this, the effects of Lomens-P0 on pro-inflammatory cytokines, prostaglandin E2 (PGE2), and hormone levels, which are factors affecting PMS symptoms (including uterine cramps, headache, and depression), were confirmed.
MATERIALS AND METHODS
Chemicals and reagents
All chemicals used in this work were purchased from commercial sources. All solvents were distilled according to standard methods prior to use. Dulbecco's modified Eagle's medium (DMEM) and fetal bovine serum (FBS) were obtained from Gibco (Waltham, MA, USA). Trypsin-EDTA was purchased from Welgene Inc. (Gyeongsan, Korea). The prefemin tablet used as a positive control was purchased from Chong Kun Dang Pharmaceutical Corporation (Seoul, Korea). Tricin, chlorogenic acid, and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolim bromide (MTT) were purchased from Sigma-Aldrich (St. Louis, MO, USA). The prolactin enzyme-linked immunosorbent assay (ELISA) kit was purchased from Molecular Innovations Inc. (Novi, MI, USA). The primary (prolactin and DRD2) and secondary antibodies were obtained from Cell Signaling Technology (Beverly, MA, USA). All other chemicals were purchased from Sigma-Aldrich.
Preparation of extracts
The H. vulgare and C. zawadskii were purchased from Gyung Dong Market (Seoul, Korea) in October 2017, and were identified by Professor Se Chan Kang, Kyung Hee University (Yongin, Korea). The voucher specimens (H. vulgare as JBR616 and C. zawadskii as JBR639) were deposited in the Laboratory of Natural Medicine Resources at the BioMedical Research Institute, Kyung Hee University. The extracts were analyzed in the following manner: the plants were washed 3 times, and then dried while avoiding direct sunlight, and cut into lengths of about 2 cm (in the case of C. zawadskii). The H, vulgare L. extract (HVE; Lot #. BPD181127) was obtained by extracting H. vulgare with 50% ethanol (10 times the weight of the H. vulgare) for 5 h at 65–75°C. The C. zawadskii var. latilobum extract (CZE; Lot #. BPD181128) was obtained by extracting C. zawadskii with 70% ethanol (10 times the weight of the C. zawadskii) at 70°C for 5 h. These extracts were concentrated at reduced pressure and less than 60°C using a rotary evaporator, and then the extracts were lyophilized to obtain the resulting powder and stored at −20°C before use. The Lomens-P0 (Lot #. BPD181130) was a mixture of the powdered extracts of HVE and CZE mixed 1:1. The extracts were dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich) and used as substrates for screening tests.
Cell culture
Rat pituitary GH3 and mouse macrophage RAW 264.7 cells were obtained from the Korean Cell Line Bank (Seoul, Korea) and were maintained in DMEM (Gibco) containing inactivated 10% FBS (Gibco), 1% penicillin (Gibco). The GH3 and RAW 264.7 cells were cultured at 37°C in a humidified atmosphere containing 5% CO2. All cells were sub-cultured twice weekly, and cell passage numbers were between 10 and 13.
Cell cytotoxicity assay
To evaluate the cytotoxicity of the extracts, an MTT assay was performed as described previously with slight modifications [20]. Briefly, the RAW 264.7 cells (5 × 103 cells/well) were seeded into 96-well plates and incubated overnight. The cells were then incubated with various concentrations of the extracts for 48 h. Ten microliters of 5 mg/mL MTT solution was added to each well and the cells incubated for an additional 4 h. The formazan crystals were dissolved in DMSO, and the absorbance at 540 nm was determined with a multi-reader (Infinite 200; TECAN, Zürich, Switzerland).
Measurement of hypothalamus cell prolactin secretion
Rat pituitary GH3 cells (3 × 105 cells/mL) were treated with a designated amount of sample for 48 h, and 100 μL of the medium was collected from each treatment well and stored at 4°C. The prolactin concentration in the GH3 cell culture medium was determined using a rat PRL ELISA kit according to the manufacturer's instructions. Briefly, 100 μL of sample and standard were placed in each well of the plate enclosed in the kit, the plates were incubated at 37°C for 90 min, and the solution was aspirated. Then, 100 μL of 1X biotinylated detection antibody was added to the empty well and the plates incubated at 37°C for 1 h. After aspirating and washing 3 times, 100 μL of 1X horseradish peroxidase conjugate solution was added to each well and allowed to react at 37°C for 30 min. After aspirating and washing 5 times, 100 μL of 1X substrate solution was added to each well, and the mixture allowed react for 5 min at 37°C in the dark. Light emission was measured as relative light units using a luminometer to confirm the amount of prolactin secretion.
Measurement of nitric oxide (NO) secretion in RAW 264.7 cells
NO production was assessed in RAW 264.7 cells: (2 × 105 cells/mL) were seeded into 96-well plates and pretreated lipopolysaccharide (LPS; 1 μg/mL) for 2 h, and the cells were then treated with the extracts for 24 h. After incubation with the extracts, 100 μL of the supernatant was removed from each well and placed into an empty 96-well plate. After adding 100 μL of Griess reagent (1% sulfanilamide and 0.1% naphthylethylenediamine dihydrochloride in 2.5% phosphoric acid; Sigma-Aldrich) to each well, the absorbance was measured at 550 nm using a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
Measurement of inflammatory markers using quantitative real-time polymerase chain reaction (PCR) analyses
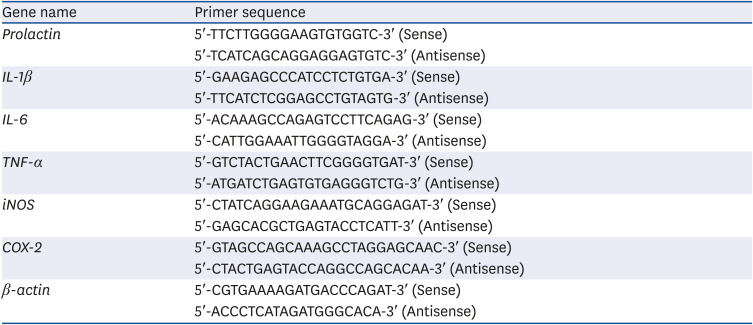
For PCR analyses using RAW 264.7 cells, the test substance was processed for 24 h, and then RNA extraction was performed. When using uterine tissue harvested from animals administered the extracts in vivo, the tissue was minced and RNA extracted. Total RNA was purified with TRIzol™ reagent (Invitrogen, Waltham, MA, USA). Total RNA (1 μg) was reverse transcribed in a 20 μL volume using oligo (dT) primers, with enzyme and buffer supplied in the PrimeScript II 1st strand cDNA Synthesis kit (Takara, Tokyo, Japan). Quantitative real-time-PCR reactions were performed on an MX3005P real-time PCR system (Stratagene, London, UK). The primers used in the experiments are listed in Table 1. For the real-time PCR, SYBR Premix Ex Taq II (Takara) was used. The final reaction volume was 25 μL containing 2 μL of cDNA template, 12.5 μL of Master Mix, 1 μL of each primer (10 μM stock solution), and 9.5 μL of sterile distilled water (DW). The thermal cycling profile consisted of a pre-incubation step at 95°C for 10 min, followed by 40 cycles of 95°C (15 s) and 60°C (60 s).
Table 1
Sequence of the primers used for real-time polymerase chain reaction
In vivo experiments in an animal model of hyperprolactinemia
A total of 60 virgin 14-week old adult female ICR mice weighing 32 g on average was used (NARA Biotech, Seoul, Korea). The animals had free access to food and water and were allowed to acclimatize for at least one day before use. Mice were housed in an environment with controlled temperature (23 ± 3°C), humidity (55 ± 4%) and ventilation rate (10–12 exchanges per hour) under a 12 h light/dark cycle. Animal experiments were performed in accordance with the current ethical regulations for animal care and use at Kyung Hee University (KHUASP(SE)-18-003).
Mice were administered metoclopramide (MCP, 20 mg/kg) by intraperitoneal injection once every 2 days; this dopamine inhibitor was used to induce hyperprolactinemia for the experimental model. Mice were divided into 10 groups of 6 individuals: normal control, MCP (induced hyperprolactinemia group, 20 mg/kg), PF (positive control group, MCP + 100 mg/kg prefemin), HVE 100 (MCP + 100 mg/kg HVE), HVE 200 (MCP + 200 mg/kg HVE), CZE 100 (MCP + 100 mg/kg CZE), CZE 200 (MCP + 200 mg/kg CZE), Lomens-P0 50 (MCP + 50 mg/kg Lomens-P0), Lomens-P0 100 (MCP + 100 mg/kg Lomens-P0), Lomens-P0 200 (MCP + 200 mg/kg Lomens-P0). Each dose of the test substances was dissolved in 200 μL saline solution and administered by gavage, while mice in the normal control and MCP group received 200 μL of saline solution by gavage. All test groups were administered saline or the test substances for 21 days. The experimental animals were fasted for 12 h and anesthetized with CO2 before sacrifice. Blood samples were collected at the completion of the experimental treatments and serum was separated for measurement of prolactin, estrogen, progesterone, follicle-stimulating hormone (FSH), luteinizing hormone (LH), and PGE2 concentrations.
High-performance liquid chromatography (HPLC) analysis
To analyze the tricin concentration in Lomens-P0, a 1260 Infinity II diode array detector (DAD) (Agilent Technologies, Santa Clara, CA, USA) was used with the DAD set at 350 nm. The Shiseido MG II column (Shiseido, Tokyo, Japan; 250 mm × 4.6 mm, 5 μm) was used with the temperature set to 40°C. The gradient elution solution consisted of water with 1% acetic acid (A) and acetonitrile (B). The mobile phase was used under binary linear-gradient conditions as follows: 0–45 min, 85–10% A; 45–55 min, 10–85% A. The flow rate was 1.0 mL/min and the injection volume was 10 μL. Lomens-P0 (20 mg/mL) and tricin (150 μg/mL) were dissolved in a 50% methanol (MeOH) solution containing 10% DMSO (DMSO:MeOH:DW = 1:5:4) at the appropriate dilution. Peaks were identified by comparing their retention time and UV-vis spectra with tricin and were quantitated using the corresponding tricin curve. To analyze the chlorogenic acid concentration in Lomens-P0, a 1260 Infinity II (Agilent Technologies) was used with the DAD set at 330 nm. An INNO C18 column (Young Jin Biochrom Co., Ltd., Seongnam, Korea; 250 mm × 4.6 mm, 5 μm) was used with the temperature set to 40°C. The gradient elution solution consisted of DW with 0.5% phosphoric acid (A) and 0.5% phosphoric acid in acetonitrile (B). The mobile phase was used under binary linear-gradient conditions as follows: 0–33 min, 92–75% A; 35–40 min, 0–92% A. The flow rate was 1.2 mL/min, and the injection volume was 5 μL. Lomens-P0 (10 mg/mL) and chlorogenic acid (150 μg/mL) were dissolved in 50% MeOH at the appropriate dilution. Peaks were identified by comparing their retention time and UV-vis spectra with chlorogenic acid, and were quantitated using the corresponding standard curve.
Statistical analyses
Data were expressed as the means ± standard error of the mean. The statistical analyses were performed using the Sigma Plot 12.0 software program (Systat Software, Inc., San Jose, CA, USA). A one-way analysis of variance was used to identify significant differences between treatment group means (P < 0.05). When significant, specific differences between groups were identified by Tukey's multiple-comparison tests.
RESULTS
Efficacy of inhibition prolactin secretion by the extracts
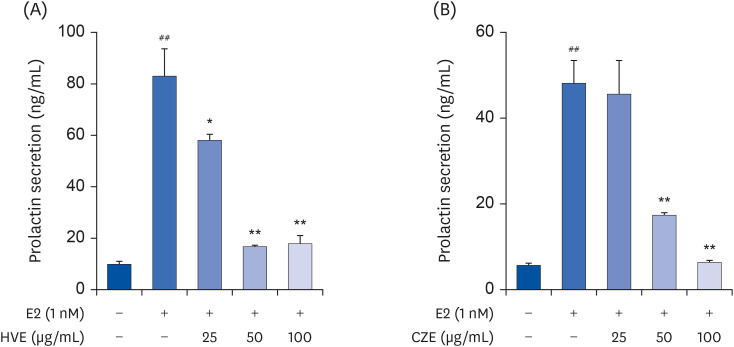
We conducted a study to test the hypothesis that inhibition of prolactin secretion could alleviate PMS symptoms. Therefore, we assessed the prolactin inhibition efficacy of HVE and CZE in GH3 rat pituitary cells. These extracts were found to inhibit prolactin secretion in a concentration-dependent manner in vitro, and by ELISA analysis, showed that both HVE and CZE induced a significant difference in inhibition of prolactin secretion at 100 μg/mL (Fig. 1). We mixed the HVE and CZE in various ratios to identify the ratios that best inhibited prolactin secretion. The results indicated prolactin secretion was effectively reduced when the extracts were mixed in a 1:1 ratio (data are not shown). We named the extract mixture ‘Lomens-P0’ and further experiments were using it.
Fig. 1
Inhibitory effect of HVE or CZE on PRL secretion in GH3 cells, the rat pituitary cell. (A) Effects of HVE on PRL secretion. (B) Effects of CZE on PRL secretion. Data are expressed as the mean ± standard error of mean (n = 3) and analyzed using a one-way analysis of variance.
HVE, H. vulgare L. extract; CZE, C. zawadskii var. latilobum extract; PRL, prolactin.
*P < 0.05 and **P < 0.01 vs. E2 control group; ##
P < 0.01 vs. normal control group.
Suppression of NO expression by the extracts
Prolactin is known to enhance immune functions in mammals [21]. We assessed NO expression as a marker of the initial inflammatory response. As shown in Fig. 2A, HVE, CZE, and Lomens-P0 were not cytotoxic to RAW 264.7 cells, and CZE suppressed the expression of NO to the greatest extent (Fig. 2B). Suppression of NO expression by CZE was significant from 50 μg/mL (P < 0.01). In the case of Lomens-P0, NO was significantly suppressed dose-dependently from 25 μg/mL (P < 0.05). On the other hand, HVE suppressed NO expression significantly only at 100 μg/mL (P < 0.05).
Fig. 2
Inhibitory effect of HVE, CZE or Lomens-P0 on NO production in RAW 264.7 cells. RAW 264.7 cells were incubated with various concentrations of HVE, CZE, or Lomens-P0 in the presence of LPS (1 μg/mL, 2 h) for 24 h. (A) Cytotoxic effects of HVE, CZE, or Lomens-P0. (B) Effects of HVE, CZE, or Lomens-P0 on the production of NO. Data were expressed as the mean ± standard error of mean (n = 3) and analyzed using a one-way analysis of variance.
HVE, H. vulgare L. extract; CZE, C. zawadskii var. latilobum extract; Lomens-P0, 1:1 mixture of H. vulgare L. extract and C. zawadskii var. latilobum extract; NO, nitric oxide; LPS, lipopolysaccharide.
*P < 0.05 and **P < 0.01 vs. LPS-treated group; ##P < 0.01 vs. normal control group.

Suppression of inflammatory marker expression by the extracts
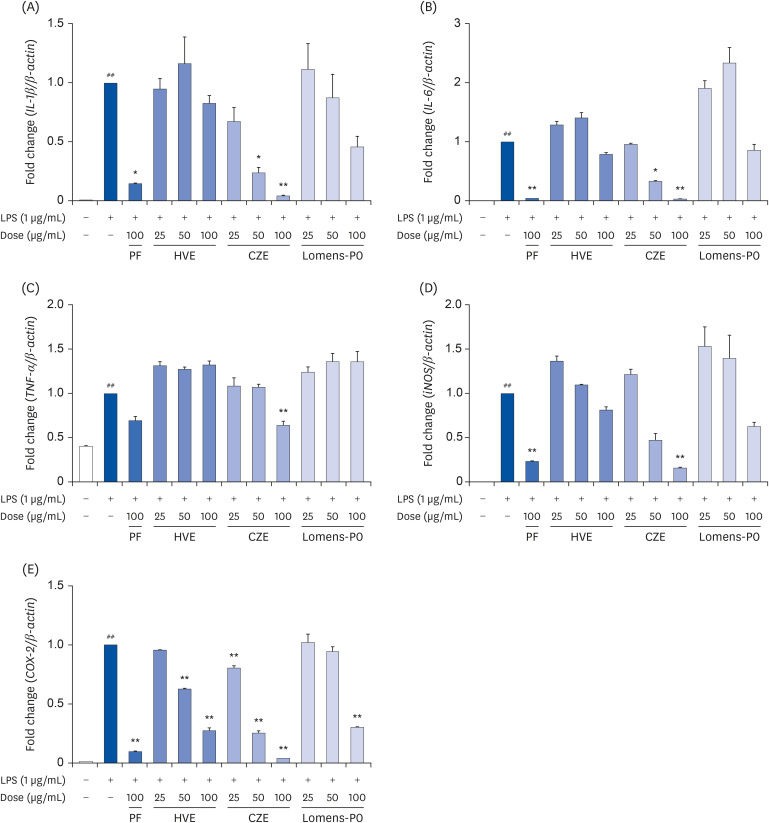
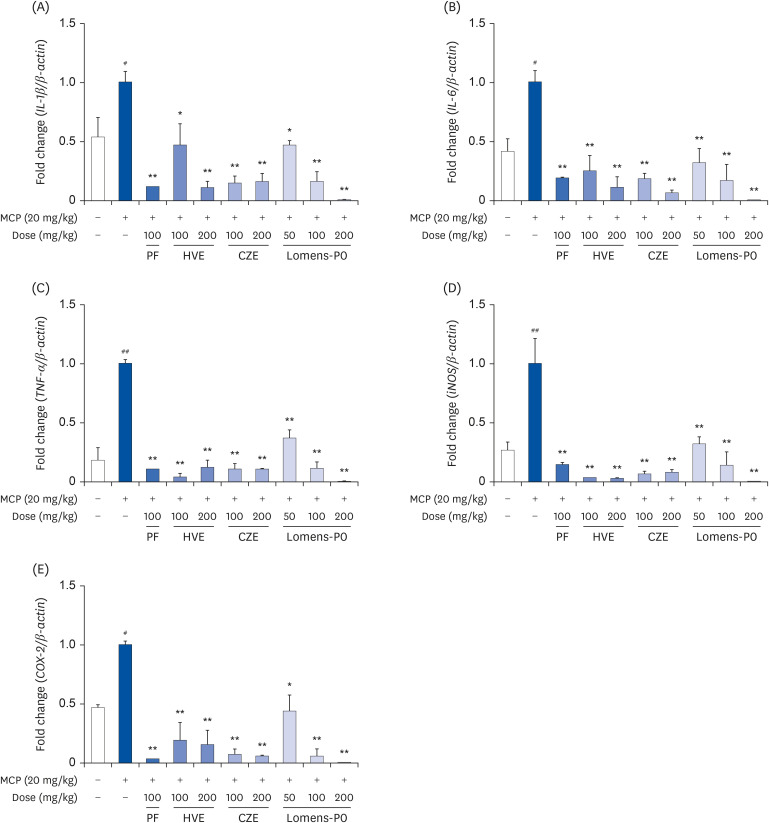
Also, prolactin is known to be involved in the regulation of various actions of immune response and inflammatory process in several physiological and pathological conditions [22]. Therefore, if HVE, CZE, and Lomens-P0 could regulate the prolactin secretion, it was expected that the inflammatory response induced by prolactin could also be controlled. We assessed mRNA expression for IL-1β, IL-6, tumor necrosis factor (TNF)-α, inducible nitric oxide synthase (iNOS), and cyclooxygenase (COX)-2 as inflammation-related markers. As shown in Fig. 3, the expression of all markers increased significantly after LPS stimulation in RAW 264.7 cells. The expression of IL-1β and iNOS tended to decrease in a concentration-dependent manner when the cells were incubated with CZE or Lomens-P0, but the effect was statistically significant only in the CZE group (Fig. 3A and D). In the case of IL-6, a significantly concentration-dependent decrease was obtained only in CZE (Fig. 3B). As shown in Fig. 3C, neither HVE nor Lomens-P0 affected TNF-α expression, but high concentrations of CZE (100 μg/mL) significantly inhibited TNF-α expression. All of the extract preparations inhibited COX-2 in a concentration-dependent manner (Fig. 3E).
Fig. 3
Effects of HVE, CZE or Lomens-P0 on inflammatory markers and mRNA expression of RAW 264.7 cells. RAW 264.7 cells were incubated with various concentrations of HVE, CZE, and Lomens-P0 in the presence of LPS (1 μg/mL, 2 h) for 24 h. The changes in the expression of mRNA levels for (A) IL-1β, (B) IL-6, (C) TNF-α, (D) iNOS, and (E) COX-2 were determined by quantitative real-time PCR. All values were normalized to β-actin expression. Data were expressed as the mean ± standard error of mean (n = 3) and analyzed using a one-way analysis of variance.
PF, prefemin; HVE, H. vulgare L. extract; CZE, C. zawadskii var. latilobum extract; Lomens-P0, 1:1 mixture of H. vulgare L. extract and C. zawadskii var. latilobum extract; LPS, lipopolysaccharide; IL, interleukin; TNF, tumor necrosis factor; iNOS, inducible nitric oxide synthase; COX, cyclooxygenase; PCR, polymerase chain reaction.
*P < 0.05 and **P < 0.01 vs. LPS-treated group; ##P < 0.01 vs. normal control group.
Body weight change in hyperprolactinemia mice following treatment with the extracts
In vivo experiments were conducted to assess whether treatment with HVE, CZE, or Lomens-P0 could inhibit prolactin secretion and control inflammation and changes in hormone secretion in an animal model. Hyperprolactinemia was induced in the mice by intraperitoneal injection of MCP, which is a gastrointestinal pro-kinetic drug used to treat gastric reflux and gastroparesis and acts by inhibiting the DRD2. Inhibition of DRD2 induces hyperprolactinemia by increasing prolactin secretion [2324]. The body weight of MCP-treated mice administered the extracts was measured once every 2 days. There were no significant differences in body weights between the groups (Supplementary Fig. 1). This result indicates that neither the extracts nor MCP were highly toxic in the hyperprolactinemia mouse model.
Organ weight change in hyperprolactinemic mice following treatment with the extracts
Organ weights were examined to determine toxicity at the tissue level and were expressed relative to body weight. As shown in Table 2, the experimental results showed that the relative weights of the liver, kidney, spleen, and uterine tissues increased after MCP injection. In mice administered MCP, relative liver weights increased 11%, while relative kidney weights increased 22%, spleen weights increased 22%, and the weights of uterine tissues increased 37%. After administration of the extracts, there was no difference between the prefemin administered group and the hyperprolactinemia induction group; however, the relative liver and kidney tissue weights were significantly reduced in the 200 mg/kg HVE treated group (P < 0.05), and relative liver weight was reduced tend at 100 mg/kg CZE, and kidney, spleen, and uterine weight were reduced tend at both 100 and 200 mg/kg CZE. Meanwhile, we found that the relative liver, kidney, spleen, and uterine weights of the 200 mg/kg Lomens-P0 treated group were similar to those in the normal control group.
Table 2
Tissue weight of hyperprolactinemia mice induced by MCP
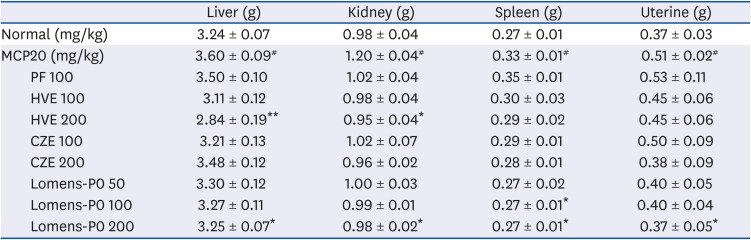
Data are expressed as mean ± standard error of mean (n = 6) and analyzed using one-way analysis of variance.
MCP20, 20 mg/kg metoclopramide; PF, prefemin; HVE, H. vulgare L. extract; CZE, C. zawadskii var. latilobum extract; Lomens-P0, 1:1 mixture of H. vulgare L. extract and C. zawadskii var. latilobum extract.
*P < 0.05 and **P < 0.01 vs. vehicle control (MCP20); #
P < 0.05 vs. normal control.
Effects of the extracts on sex hormones secretion in the hyperprolactinemia mouse model
We induced hyperprolactinemia in mice and then examined the changes in hormone secretion. In our experiments, the progesterone/estrogen ratio of all experimental groups did not increase to a level close to the normal control, but it increased significantly compared to hyperprolactinemia in groups of 100 mg/kg HVE and 200 mg/kg Lomens-P0 (Fig. 4A). We judged that this result was due to inhibition of prolactin secretion. The suppression of prolactin secretion was more dramatic in the Lomens-P0 group than in the HVE and CZE groups, and hormonal secretion resulting from this was also confirmed significantly (Fig. 4B). As shown in Fig. 4C and D, LH secretion inhibited by MCP showed significant changes at 200 mg/kg CZE and 100 and 200 mg/kg Lomens-P0, and FSH secretion was dramatically increased in all groups provided Lomens-P0 (P < 0.01) and was similar to the normal control group level. Meanwhile, as shown in the results of Fig. 4E, PGE2 secretion was inhibited by all of the extract preparations, and the prefemin used as a positive control was found to inhibit PGE2 secretion to a similar degree.
Fig. 4
Effects of HVE, CZE or Lomens-P0 on sex hormones and inflammatory mediators in the serum of hyperprolactinemic mice. The hyperprolactinemia mouse model was prepared by intraperitoneal injection of MCP (20 mg/kg). Untreated animals served as normal controls. (A) Serum progesterone/estrogen ratio, (B) Serum PRL, (C) Serum LH, (D) Serum FSH, and (E) Serum PGE2. Data were expressed as the mean ± standard error of mean (n = 6) and analyzed using a one-way analysis of variance.
PF, prefemin; HVE, H. vulgare L. extract; CZE, C. zawadskii var. latilobum extract; Lomens-P0, 1:1 mixture of H. vulgare L. extract and C. zawadskii var. latilobum extract; MCP, metoclopramide; PRL, prolactin; LH, luteinizing hormone; FSH, follicle-stimulating hormone; PGE2, prostaglandin E2.
*P < 0.05 and **P < 0.01 vs. MCP-treated group; #P < 0.05 and ##P < 0.01 vs. normal control group.

Effects of extracts on uterine inflammation in a hyperprolactinemic mouse model
The in vivo experiments were conducted to determine whether the extracts could inhibit prolactin secretion in animal models and also control inflammation. Mouse uterine tissue was collected and inflammatory marker expression was determined at the mRNA level. As shown in Fig. 5, all of the extract preparations inhibited the expression of mRNA for the inflammatory markers in a dose-dependent manner, especially high-dose Lomens-P0. These results indicate that treatment with Lomens-P0 could suppress hormonal imbalances, inflammatory responses in the uterus, and pain caused by hyperprolactinemia.
Fig. 5
Effects of HVE, CZE or Lomens-P0 on uterine inflammation in hyperprolactinemic mice. The hyperprolactinemia mouse model was prepared by intraperitoneal injection of MCP (20 mg/kg). Untreated animals served as controls. Changes in the expression of mRNA for (A) IL-1β, (B) IL-6, (C) TNF-α, (D) iNOS, and (E) COX-2 were determined by quantitative real-time PCR. All values were normalized to β-actin expression. Data were expressed as the mean ± standard error of mean (n = 6) and analyzed using a one-way analysis of variance.
PF, prefemin; HVE, H. vulgare L. extract; CZE, C. zawadskii var. latilobum extract; Lomens-P0, 1:1 mixture of H. vulgare L. extract and C. zawadskii var. latilobum extract; MCP, metoclopramide; IL, interleukin; TNF, tumor necrosis factor; iNOS, inducible nitric oxide synthase; COX, cyclooxygenase; PCR, polymerase chain reaction.
*P < 0.05 and **P < 0.01 vs. MCP-treated group; #P < 0.05 and ##P < 0.01 vs. normal control group.
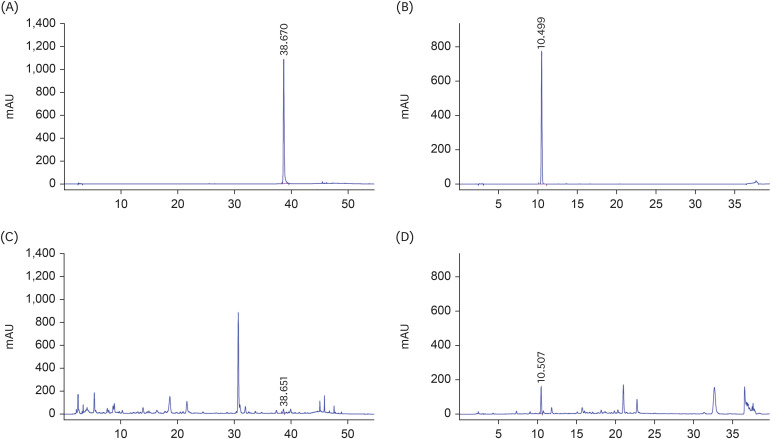
Identification of the active components in Lomens-P0 by HPLC analysis
Tricin, an active component of H. vulgare, was identified in the Lomens-P0 mixture by HPLC analysis. Fig. 6A is the standard chromatogram of tricin, and Fig. 6C is the result of the Lomens-P0 analysis. Tricin was detected after 38.6 min at a concentration of 0.18 ± 0.005 mg/g in Lomens-P0. Chlorogenic acid, which is considered an active constituent of C. zawadskii, was also assessed in Lomens-P0 by HPLC analysis, the result of which are shown in Fig. 6B and D. Chlorogenic acid was detected after 10.5 min at a concentration of 3.43 ± 0.06 mg/g in Lomens-P0. We conducted additional tests to select active constituents to help alleviate PMS symptoms.
Fig. 6
HPLC analyses. (A) Chromatogram of the tricin standard; (B) Chromatogram of the chlorogenic acid standard; (C) Chromatogram of tricin in Lomens-P0; and (D) Chromatogram of chlorogenic acid in Lomens-P0. All experiments were performed in triplicate.
HPLC, high-performance liquid chromatography; Lomens-P0, 1:1 mixture of H. vulgare L. extract and C. zawadskii var. latilobum extract.
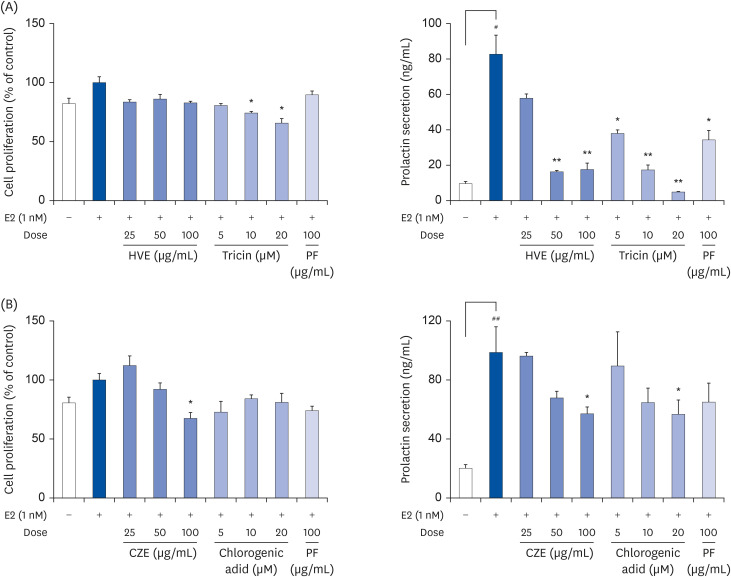
Critical components influencing prolactin secretion
Prolactin secretion was measured after incubating GH3 cells with the active constituents of Lomens-P0 identified through HPLC analysis. As shown in Fig. 7, all constituents inhibited prolactin secretion in a concentration-dependent manner. Tricin was detected only in HVE and chlorogenic acid was detected only in CZE. Thus, these constituents were selected as indicator components and are considered the active ingredients in HVE and CZE.
Fig. 7
Inhibition of PRL secretion by active components identified in HVE or CZE. GH3 cells were treated with various concentrations of the active compounds. PRL secretion and cell viability were assessed using an ELISA assay. (A) Effects of HVE and tricin on cytotoxicity and PRL secretion, (B) Effects of CZE and chlorogenic acid on cytotoxicity and PRL secretion. Data were expressed as the mean ± standard error of mean (n = 3) and analyzed using a one-way analysis of variance.
PF, prefemin; HVE, H. vulgare L. extract; CZE, C. zawadskii var. latilobum extract; ELISA, enzyme-linked immunosorbent assay; PRL, prolactin.
*P < 0.05 and **P < 0.01 vs. MCP-treated group; #P < 0.05 and ##P < 0.01 vs. normal control group.
DISCUSSION
Severe PMS is a common disease among women of reproductive age and affects both the social life and work productivity of affected women. However, the biologic mechanisms underlying symptoms in PMS remain to be determined [25]. Currently, for relief of severe PMS symptoms, ovulation suppression or serotonin reuptake inhibitors are recommended; however, side effects of pharmacological treatment may occur, and many women prefer natural approaches. For these women, alternative medicines could be an option [26]. And the fruit of Vitex agnus castus is one commonly used alternative medicine. The mechanism of action of V. agnus castus is related to modulation of prolactin secretion by dopamine, without directly affecting LH and FSH [27]. Prefemin, used as a positive control in this study, was developed and marketed as a PMS palliative drug using V. agnus castus.
Prolactin hypersecretion is one of various causes of PMS. Hyperprolactinemia induces hormone imbalances, resulting in an inflammatory response throughout the body [28]. Inflammation in the context of PMS leads to unspecified pain in the back and abdomen, bloating, and weight gain due to appetite enhancement [29]. Therefore, we hypothesized that suppression of excessive prolactin secretion can suppress PMS symptoms. In previous studies, we confirmed that HVE, an extract of H. vulgare, and CZE, an extract of C. zawadskii, inhibited prolactin hypersecretion. In the present study, we evaluated the efficacy of Lomens-P0, a mixture of these extracts, to inhibit prolactin secretion. We confirmed the inhibitory effects of HVE, CZE, and Lomens-P0 on prolactin hypersecretion in GH3 cells (Fig. 1) and cytotoxic and anti-inflammatory effects in RAW 264.7 cells (Figs. 2 and 3). NO is a signaling molecule that plays an important role in the pathogenesis of inflammation. It exerts an anti-inflammatory effect under normal physiological conditions but is considered a pro-inflammatory mediator causing inflammation when overproduced under abnormal conditions [30]. After inducing abnormal physiological conditions with LPS, we applied the extracts and confirmed their effects on pro-inflammatory mediators. Gene expression of iNOS and COX-2, which induce NO, and expression of pro-inflammatory cytokines including IL-1β, IL-6, and TNF-α showed significant differences mainly in CZE. Lomens-P0 was confirmed to inhibit COX-2 expression (P < 0.01). We conducted in vivo experiments based on the results showing that the extracts affect the release of prolactin and the expression of inflammatory substances at the in vitro level.
Prolactin plays an important role in metabolism, immune system regulation, and pancreatic development. It stimulates liver growth and regeneration by promoting liver cell proliferation and angiogenesis in adults [31] and is eliminated by the liver and kidney [32]. Prolactin has a physiologically active function as a hormone and cytokine, managing maturation of CD4− CD8− thymocytes to CD4+ CD8+ T cells through IL-2 receptor expression, as well as a role in inducing innate and adaptive immune responses. Thus, hyperprolactinemia has been reported in several autoimmune patients [15]. In our animal experiment, both MCP and extract were harmless as noted by change in body weight during the experiment. However, there was a change in organ weight due to MCP. As shown in the results of Table 2, the weights of the liver, spleen, kidney, and uterus all increased compared to the normal control group. The increase in liver and kidney weight in the MCP group in this experiment was a result of prolactin-induced liver cell growth or excessive prolactin-eliminating load. It was concluded that the increase in weight of the spleen was due to promotion of the immune response. A previous study confirmed that inducing hyperprolactinemia using MCP did not cause a significant change in uterine weight [33]. However, in our study, the weight of the uterus was significantly increased. This may be due to an inflammatory response and tissue swelling due to an increase in prolactin. Or, it could be a uterine weight change caused by a steroid hormone [3435]. In our study, it is thought that the weight of the uterus may have been increased by excessively secreted estrogen (as indicated by the reduced progesterone/estrogen ratio). Further research is needed to validate these findings.
As with the results of cell experiments, the extracts had a significant effect on reduction of serum prolactin secretion and regulation of sex hormones including progesterone, estrogen, LH, and FSH in hyperprolactinemic mice (P < 0.05). In addition, the secretion of PGE2 was reduced. PGE2 has physiological and pathological functions in various cells and tissues [36]. In particular, PGE2 is well known to regulate reproductive processes including ovulation, fertilization, implantation, luteolysis, and uterine contractility, and impaired PG metabolism can induce PMS symptoms (excessive uterine contraction and abdominal pain) [3738]. Therefore, appropriate regulation of PGE2 secretion is an important factor in relieving PMS symptoms. In our study, secretion of PGE2 was decreased significantly by high concentrations of HVE, CZE, and Lomens-P0. The extracts also influenced the expression of mediators related to inflammation in animal experiments. MCP injection significantly increased the gene expression of IL-1β, IL-6, TNF-α, iNOS, and COX-2 at the mRNA level, and significant decreases were found in all groups that administered HVE, CZE, and Lomens-P0 (P < 0.01). These effects were more pronounced when providing Lomens-P0, attributed to tricin in HVE and chlorogenic acid in CZE.
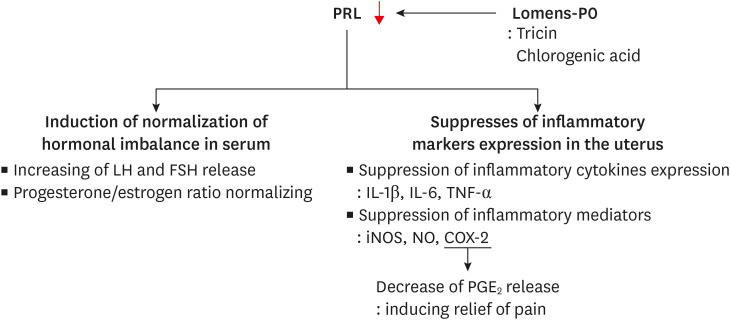
In this study, we measured biomarkers of sex hormone imbalance and inflammatory mediators increased by hyperprolactinemia, which is the posited cause of PMS. When Lomens-P0 was given to hyperprolactinemic mice, serum prolactin and sex hormone levels were similar to those of normal controls, while the gene expression of inflammation-related biomarkers was significantly reduced (Fig. 8). Though it is necessary to validate the results in humans, this study showed the potential of developing materials of Lomens-P0 for PMS symptom relief.
Fig. 8
Summary of this paper. Lomens-P0 normalizes the hormone imbalance and suppresses the expression of inflammatory cytokines and mediators involved in PMS symptoms. These results are due to the prolactin secretion inhibitory effect of Lomens-P0 (especially tricin and chlorogenic acid).
Lomens-P0, 1:1 mixture of H. vulgare L. extract and C. zawadskii var. latilobum extract; PMS, premenstrual syndrome; PRL, prolactin; LH, luteinizing hormone; FSH, follicle-stimulating hormone; IL, interleukin; TNF, tumor necrosis factor; iNOS, inducible nitric oxide synthase; COX, cyclooxygenase; NO, nitric oxide; PGE2, prostaglandin E2.
 XML Download
XML Download