

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Chronic-allergic asthma is an airway inflammation that increases the number of eosinophils and other inflammatory cells and releases pro-inflammatory mediators, promoting allergic inflammatory cells. It causes submucosal bronchial layers to vasoconstriction, leading to breathing problems [1-3]. Ovalbumin-induced airway inflammation in an asthma mouse model results in increased eosinophil recruitment via T cell activation [4,5]. In addition, it results in massive secretion of Th2 cytokines regulated by transcription factors such as nuclear-activated T cell factor (NFAT), interleukin (IL)-13, -4, and -5, and tumor necrosis factor (TNF)-α in the lungs, blood serum, and bronchoalveolar lavage fluid (BALF) [1,2,6-8].
Allergic asthma with eosinophilic inflammation causes changes in IL-5 regulation, which facilitates the formation and differentiation of eosinophils from myeloid precursor cells [9,10], but also increase reactive oxygen species (ROS) levels, including nitric oxide [11,12]. Mepolizumab is a commonly used anti-IL-5 therapy for severe allergic asthma associated with eosinophilia; however, mepolizumab intensifies asthma by causing a prolonged decrease in eosinophils with dramatic lymphocyte development [13-15], limiting its use for severe-allergic asthma. Thus, effective and safe alternatives are required for the treatment of chronic allergic asthma.
Phytonutrients such as flavonoids, found in fruit and vegetables, red wine, tea, and regular foods [16] reduce the risk of asthma [17]. Additionally, following the consumption of fruit juice and red wine, flavonoids offered a protective effect due to antioxidant activity [18]. Furthermore, a recent study identified flavonoids as potential therapeutic candidates for asthma treatment by modulating immune mediators and binding DNA sequencing factors, thus targeting oxidizing molecules [19]. Besides, a report indicated that flavonol (fisetin) suppresses the production of cytokines (IL-5) in the human cell line [20].
The molecular mechanisms of naringenin have been identified in airway inflammatory disorders such as bronchial asthma, pulmonary and cystic fibrosis, chronic obstructive pulmonary disease, and lung cancer [21-23]. It was supported with substantial inhibition of eosinophilic airway inflammation, airway hyperreactivity, and production of Th2 cytokine from CD4 T cells in ovalbumin challenged mice treated with naringenin chalcone [24]. Furthermore, hesperetin and naringenin significantly reduced lung atelectasis, airway smooth muscle hypertrophy, and sub-epithelial fibrosis in the mouse model of asthma [25]. It was also found that both naringin and naringenin might relax the tracheal smooth muscle by opening a wide Ca2+-activated K+ conductance channel that mediates hyperpolarization of the plasma membrane. In addition, it decreased Ca2+ influx, suggesting a potentially productive therapeutic approach to naringin and naringenin for cough variant asthma [26]. Another study found that experimental mice with low IgE serum levels had considerably lower IL-13 and IL-4 in their bronchoalveolar lavage fluid, implying that naringenin plays a protective function in the asthma response [27,28].
Morin, a bioflavonoid similar to naringenin, demonstrated similar antitussive efficacy compared to codeine (an opioid antitussive) as a potential compound for inhibiting bronchial asthma. A study found that morin (30 and 100 mg/kg) significantly decreased serum and BALF (IL-4 and IL-13) levels in OVALBUMIN (OVA)-sensitized guinea pigs, as well as eosinophilic lung tissue and mastocyte infiltration, similarly to that of the corticosteroid budesonide [23]. In another study, morin administration significantly reduced goblet cell hyperplasia and collagen deposition/fibrosis. In addition, morin inhibited the OVA-induced rise in IgE, TNF-, IL-4, IL-13, metalloproteinase-9 matrix, and malondialdehyde it is a potent asthmatic agent that reduces airway inflammation in chronic asthma [29].
Apart from the pertinent data from numerous published studies on naringenin and morin, it is clear that no in vitro or in vivo studies on anti-IL-5 therapy as a safe and effective treatment for severe allergic asthma associated with eosinophilia have been conducted to date. However, it is a promising treatment for severe allergic asthma associated with eosinophilia.
Therefore, the current investigation was planned to study the in vitro and in vivo effects of naringenin and morin on down-regulation of signalling pathways involving the secretion of IL-5 and ROS in EL-4 cells induced by PMA+ionomycin through distinct pathways. The in vitro findings were further corroborated by determining asthma-dependent cytokines such as IL-13, IL-5, IL-4, and TNF-α for their suppressive actions on the inflammation caused airways due to allergic asthma by ovalbumin, followed by phosphorylation of mitogen-activated protein kinase and Akt, namely p38 and Erk1/2 in serum, BALF and lung tissue declining eosinophilic aggregation in the airways of the lung tissue caused by OVA-induced asthma in the mouse model.
METHODS
Chemicals
Citrus flavonoids (naringenin and morin, 98% purity), PMA (phorbol 12-myristate 13-acetate), dimethyl fumarate, ionomycin, rosiglitazone and retinoic acid were purchased from Sigma Aldrich Co., Beijing, China. NFAT inhibitor was obtained from Pure Chemicals Co. (Beijing, China). Anti-(Nrf2, -NFAT), anti-(β-actin, -PI3k, -Akt, -ERK, -p38, -JNK) and anti-phospho-(PI3k, -p38, -ERK, -JNK, and -Akt) were obtained Cell Signaling Technology (Beijing, China). Anti-(PPARγ, -lamin B1, and -HO-1) antibodies were obtained from Cell Signalling Technology. horse serum, Dulbecco’s modified Eagle’s medium and penicillin/streptomycin antibiotics were procured from Sigma Aldrich.
Cell culture
The BCRC 60179 cell line (EL-4 murine T-lymphoma) was acquired from the research centre (Beijing, China). Modified eagle’s medium (Dulbecco) was used to grow the cells supplemented by 10% horse serum. The subculture of confluent cells was carried at 1:4 with alteration of medium three times over a week with a supply of 5% CO2 under humid room temperature.
Cell viability evaluation
The cell viability tests were carried out following previous studies [30], where the cells were treated with naringenin and morin concentrations, followed by suspension in trypan blue solution to determine cell viability.
Experimental design
The Institution's Animal Ethics Committee (Beijing, China) approved all experimental procedures (RCC/BJ/04/2019_045). Before the experiments commenced, the BALB/c mice (either sex eight weeks old) were allowed to maintain at 20°C–21°C with a 12 h day/night cycle in the air-conditioned facility. After a week of acclimatization, four groups of mice were formed, each with ten mice. The control (without induction of allergic-asthma) mice (n = 10) were orally administered distilled water (0.5 ml).
A 3:1 ratio of ovalbumin (700 μg/ml) to alum (300 μg/ml) was prepared and adjusted to the final volume. Immunization was induced in mice (n = 30) by intraperitoneal administration of OVA (10 ml/kg) mixed with alum on day 0 to 15 days. The ovalbumin-sensitized mice were divided into three groups: allergic-asthma model, naringenin group, and morin group, each with ten mice. Ovalbumin sensitized mice with allergic asthma were given a vehicle (0.5 ml of phosphate-buffered saline without OVA) between 16–28 days together with 3% OVA (w/v) prepared in a phosphate buffer solution for 30 min via a nebulizer at a flow rate of 0.4 ml/min for eight days.
However, ovalbumin sensitised mice in treatment groups (n = 10) were given 0.5 ml flavonoids (naringenin and morin; prepared in phosphate-buffered saline; 100 mg/kg/day) orally from 16 to 28 days. Nevertheless, ovalbumin sensitised mice in treatment groups (n = 10) were given 0.5 ml flavonoids (naringenin and morin; prepared in phosphate-buffered saline; 100 mg/kg/day) orally from 16 to 28 days and were exposed to 3 percent OVA (w/v) inhalation prepared in a phosphate buffer solution for 30 min in a nebulizer chamber at a flow rate of 0.4 ml/min.
After treatments, mice were sacrificed with ketamine injections (2.5 mg/kg) on the 30th day after fasting overnight. Blood samples were collected, followed by lavaging from the lungs with ice-cold phosphate buffer (0.3 ml, three times). After centrifugation, the supernatant from BALF was preserved at −80°C until cytokine assays were performed. The sediment-containing pellets were used for cell counting and intracellular ROS assessment. Lung tissue slices were preserved in 10% formalin solution for histological analyses.
Western blot analysis and ELISA assay (enzyme-linked immunosorbent)
The production of IL-5 in PMA+ionomycin-treated EL-4 cells was measured quantitatively for 24 h with and without naringenin or morin (100 μM), as recommended by the ELISA kit (Sigma Aldrich). The nucleus and cytoplasmic proteins were isolated after treatment with and without naringenin or morin, as previously depicted [31,32].
The lung tissues were homogenized in a phosphate buffer solution comprising RIPA lysis buffer (100 mM NaCl, 3 mM EDTA, 40 mM Tris, 0.3 percent SDS and 3 percent Triton X-100 adjusted pH to 7.5) with a blend of protease and phosphatase inhibitors. At 4ºC, the lysate was subjected for centrifugation at 15,000 rpm for 25 min. Protein concentration was assessed using a protein estimation kit (Sigma Aldrich), with BSA (bovine serum albumin) as the standard. In addition, the ELISA kits were employed to determine the levels of proteins such as IL-(13, -5, and -4) and TNF-α in the lung tissues and BALF as per the manufacturer's protocol.
The protein (40 μg) were prepared for western blot analysis onto SDS-PAGE (12 per cent) gels and then placed onto the nitrocellulose membranes. The primary antibodies were bound to the membranes and incubated at 4ºC for 12 h. The membranes with primary antibodies were washed four times and incubated for 2 h with conjugated antibodies (horseradish peroxidase), followed by chemiluminescence visualization. The following NFAT, Nrf2, PPARγ, HO-1, PI3k, Akt, ERK, JNK, p38, phospho-(PI3k, -Akt, -ERK, -JNK, -p38), lamin B1 and β-actin for the proteins isolated from PMA+ionomycin treated EL-4 cells; while Akt, phospho-(Akt and -p44/42), p44/42 MAPK (Erk1/2), MAPK (p-Erk1/2), phospho-p38 MAPK, p38 MAPK and β-actin were used as primary antibodies with 1:1,000 dilution (Cell Signaling Technology).
Quantification of intracellular ROS
EL-4 cells induced by PMA+ionomycin generated intracellular ROS in the presence and absence of naringenin or morin (100 μM) was determined using 2’,7’- dichlorodihydrofluorescein diacetate (DCHF-DA) as described by Cheng et al. [33]. Likewise, the intracellular ROS produced from the BALF cells was measured as previously described [33], where the cells obtained from the BALF were incubated for 30 min with DCHF-DA (100 μM). The cells were washed thrice using a phosphate buffer solution (pH 7.2), and DCF fluorescence was measured using a fluorometric microplate reader at excitation and emission wavelengths of 450 and 520 nm.
Serum alanine (ALT) and aspartate transaminases (AST) estimation
Serum enzymes such as ALT and AST were determined after treatment with naringenin or morin (100 mg/kg/day) for one week in control mice (without asthma), positive control mice (asthma induced), and treatment groups using commercial kits (Asian pure chemical, Beijing, China).
Inflammatory cell count in BALF
The inflammatory cells count was determined, as illustrated earlier in the BALF [34]. The cells from the sediment were collected from the BALF after centrifugation. The sediment was washed with phosphate buffer solution four times and resuspended in PBS (200 μl). Total cell counts were measured with a hemocytometer, and then a Diff-Quik staining reagent was employed to stabilize and stain. Different cells were counted and determined depending on their ability to stain and morphology.
RESULTS
Effects of naringenin and morin on EL-4 cells induced by PMA+ionomycin
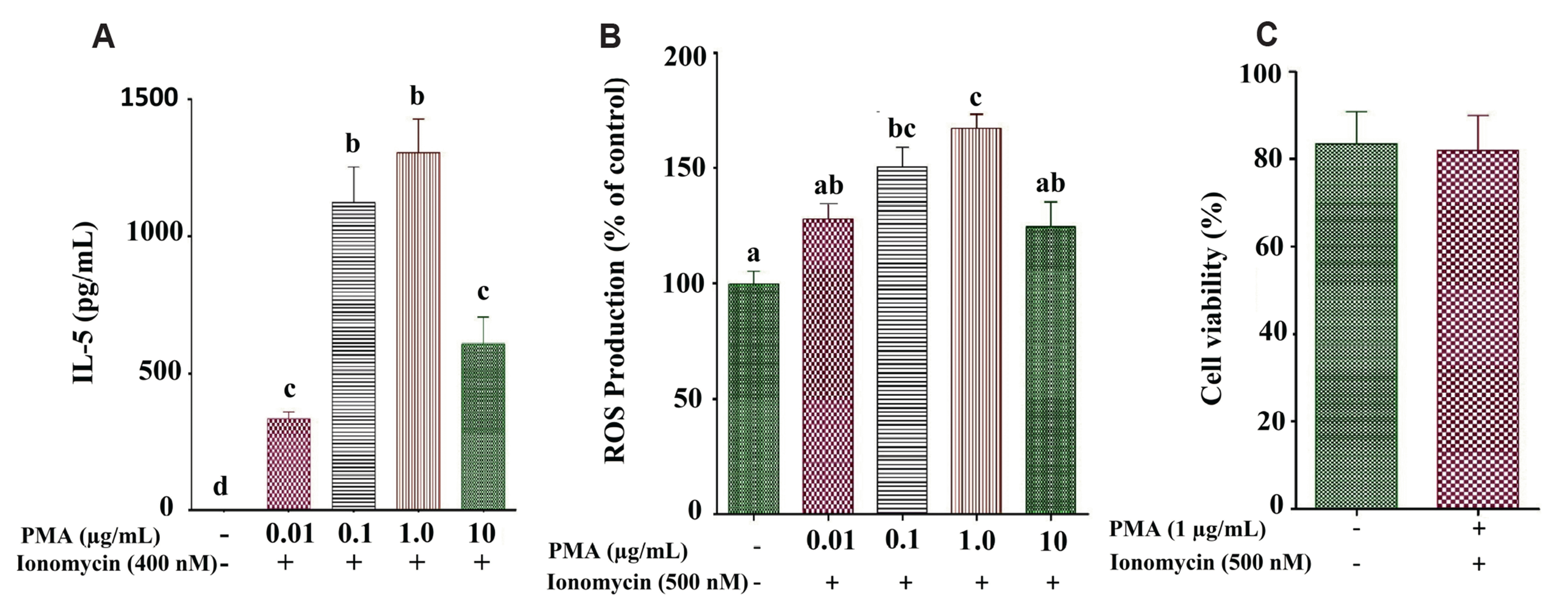
Regulation of PMA+ionomycin mediated IL-5 and ROS production in EL-4 cells: Several studies suggested that the increase in the production of Th2 cytokine is mediated by PMA+ionomycin in EL-4 cells. Our findings (Fig. 1A, B), following several studies, showed increased development of ROS and IL-5 with PMA (150 ng / ml) plus ionomycin (500 nM) in EL-4 cells treated for 48 h. Besides, 150 ng/ml PMA plus 500 nM ionomycin treated for 48 h was not impacted to cell viability compared with untreated cells (Fig. 1C)
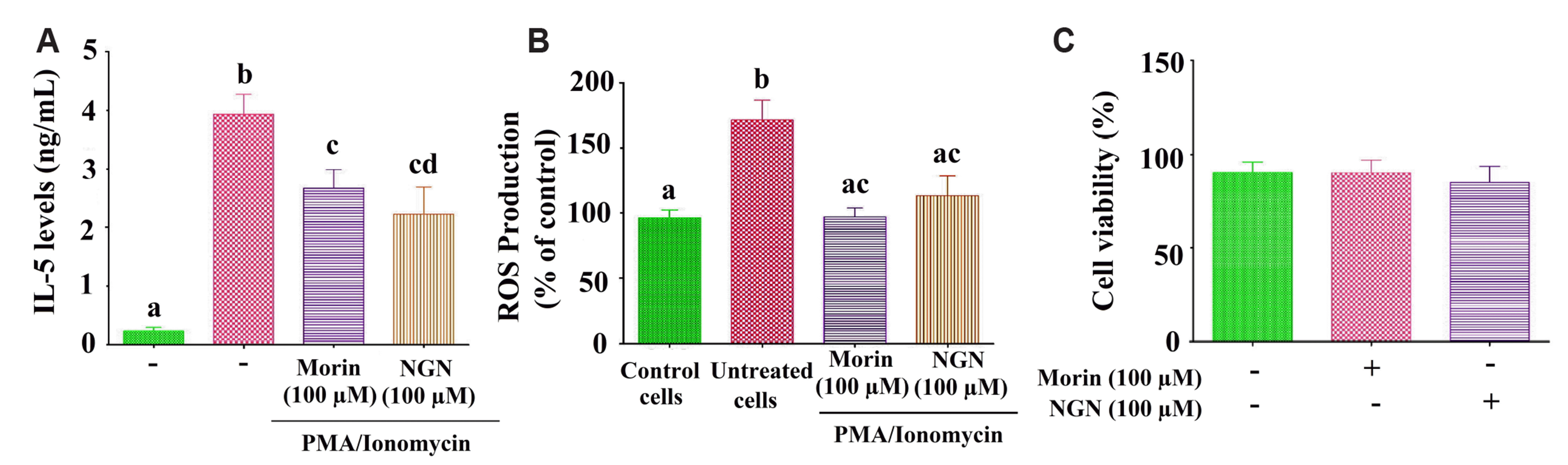
Naringenin or morin inhibited NFAT protein expression regulating the IL-5 secretion: Naringenin or morin exhibited improved anti-inflammatory and antioxidant activity; our present study, therefore, aimed to explore their suppressive actions (Fig. 2A, B) on ROS and IL-5 generation in PMA+ionomycin treated EL-4 cells (p < 0.05). Furthermore, no substantial cytotoxic changes were observed in treated naringenin or morin cells (100 μM; Fig. 2C) compared to untreated naringenin or morin cells in cell viability studies. The dose-dependent selection of doses (50 and 100 M) for the experiments using naringenin or morin was based on the inhibitory effect of NFAT (transcription factor) protein expression, which is responsible for regulating the IL-5 secretion in PMA+ionomycin treated EL-4 cells (Fig. 3).
Naringenin or morin induced heme oxygenase-1 (HO-1) expression in EL-4 cells through Nrf2 involving the signalling pathway PI3K/AKT or ERK/JNK: Fig. 4 showed increased cytosolic Nrf2 and HO-1 protein expression levels after 6-hour treatment with naringenin incubated in PMA+ionomycin-induced EL-4 cells. Besides, Heme oxygenase-1 expression mediated through related kinases were showed to augment the phosphorylation of PI3K/Akt, ERK and c-JNK in EL-4 cells after treating with naringenin (Fig. 4). Our study findings follow the report showing upregulation of HO-1 expression in human alveolar epithelial cells treated with rosiglitazone through modulation of the transcription factor, Nrf2 or peroxisome proliferator-activated receptor (PPARg).
Our findings further noticed that increased expression of nuclear PPARg and HO-1 protein expression in PMA+ionomycin treated EL-4 cells after 6 h morin (100 μM) treatment (Fig. 4). Furthermore, phosphorylation of related ERK kinases was increased after treatment with morin (100 M) for 5 h. Such results were consistent with the previously reported HO-1 study regulated by transcription factors, namely Nrf2 or PPARg.
Effects of naringenin and morin on ovalbumin-induced allergic asthma in a mouse model
Total and different inflammatory BALF cells: The treatment of naringenin or morin demonstrated a dramatic (p < 0.05) decline in the number of inflammatory total cells and eosinophils in the BALF relative to the asthma-induced group (Table 1) as compared to the control group, and the findings suggested that OVA was capable of causing inflammation of the airways in asthma-induced mice.
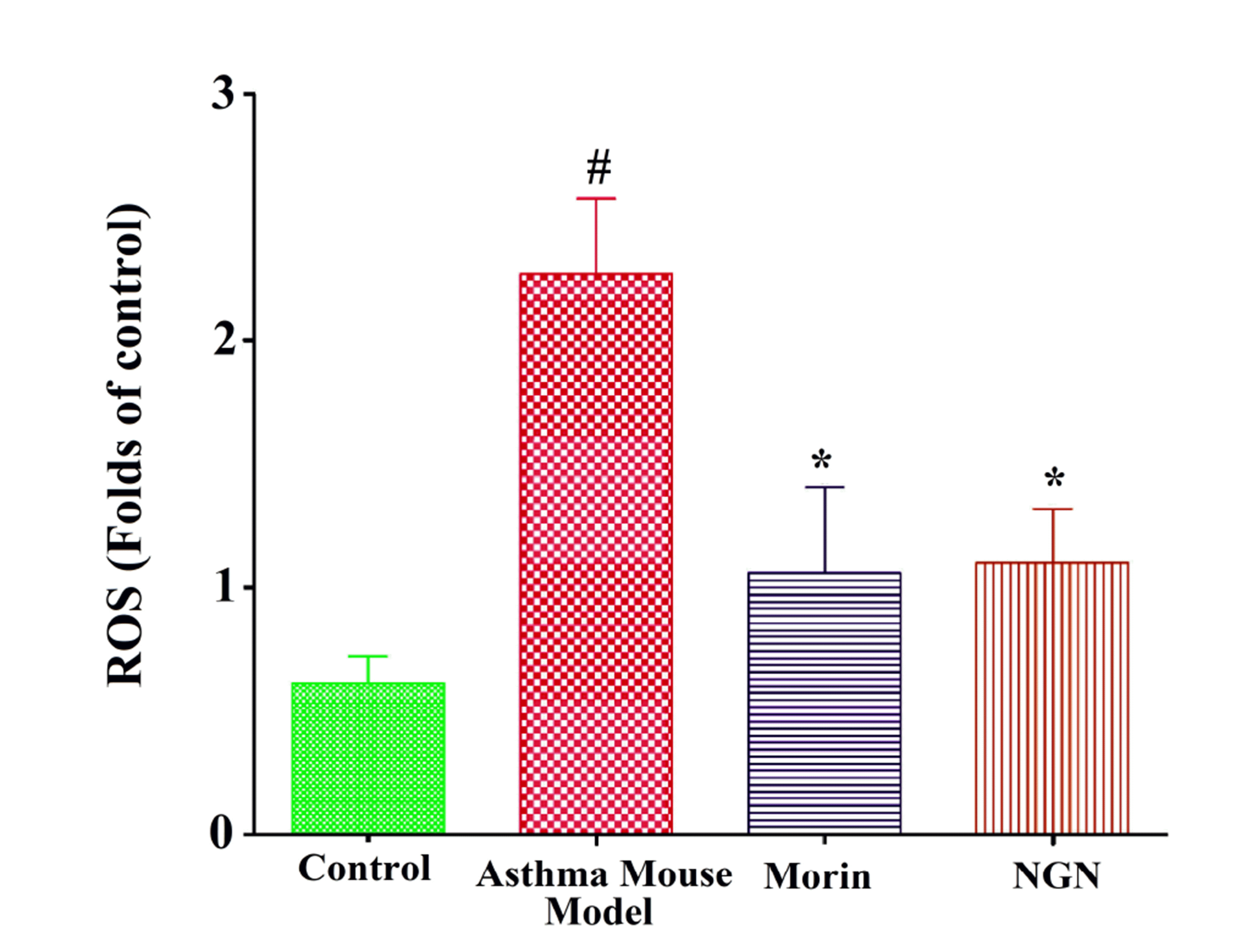
Intracellular ROS generation in BALF: The effect of naringenin or morin in the ovalbumin-induced asthma mice showed decreased ROS generation compared to control mice (Fig. 5), indicating ROS mechanism in bronchial airway inflammation as a significant benefit in allergic asthma.
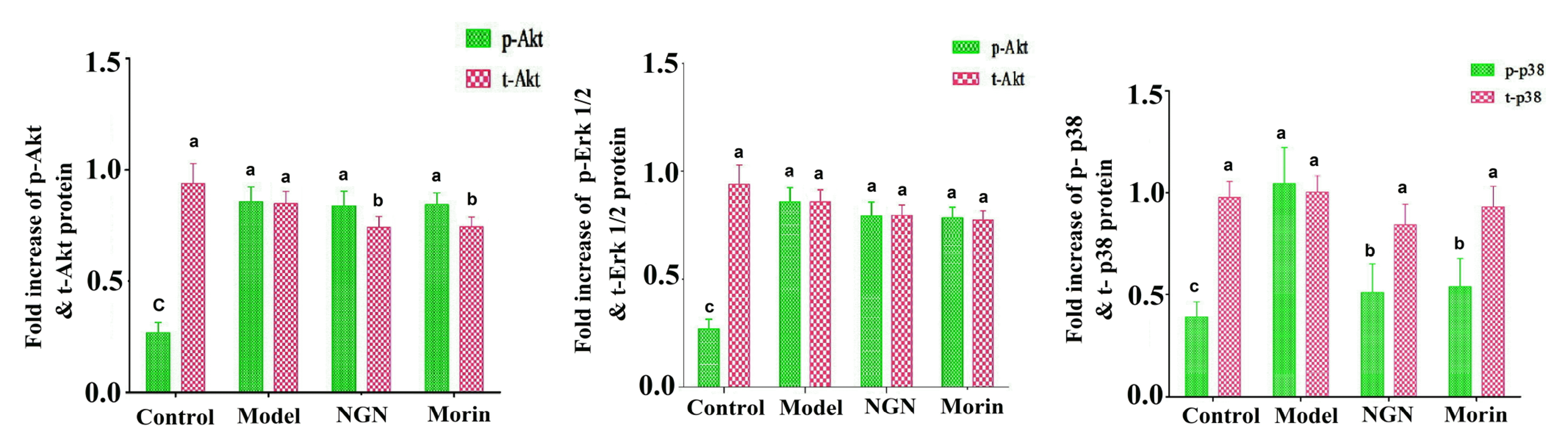
Akt and MAPK phosphorylation pathways in lungs: Naringenin or morin's inhibitory effects on the secretion of IL-5 in PMA+ionomycin treated EL-4 cells were further corroborated with naringenin or morin's inhibitory effects on cytokine production IL-(4, -13, and -5) and TNF-α through the Akt phosphorylation and MAPK signalling pathways in OVA-sensitized mice, emphasizing their role in allergic asthma (Fig. 6).
AST and ALT levels: The results of naringenin or morin demonstrated no substantial modifications in serum levels of AST and ALT between the treated and ovalbumin-induced asthma mice, thereby confirming the absence of hepatic toxicity in mice (Table 2).
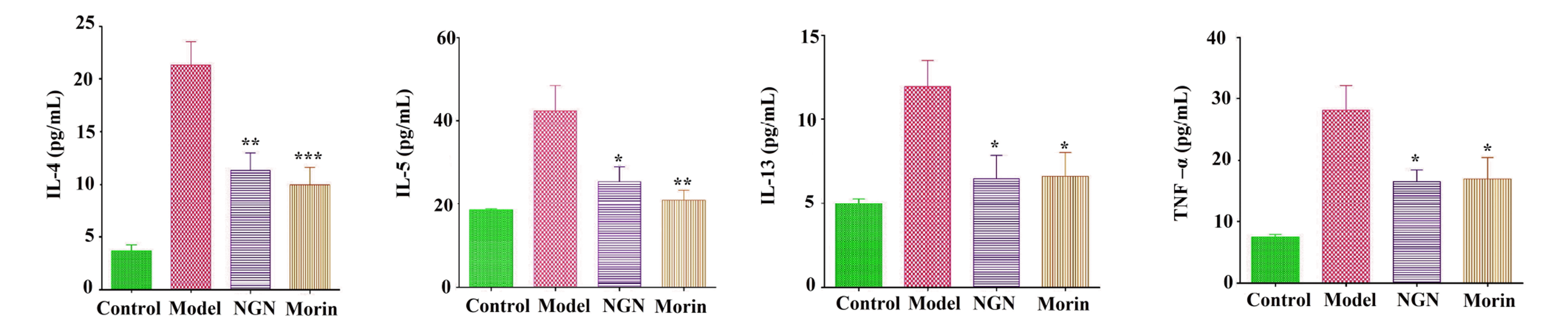
BALF cytokine levels and lungs: Naringenin or morin treatment decreased significantly Th2 cytokines levels in lung tissues and BALF in OVA-induced asthma mice compared with the untreated asthma group. Besides, TNF-α levels in lung tissues and BALF with naringenin or morin treatment for ovalbumin-induced asthma were dramatically reduced relative to the untreated asthma group, thus corroborating the findings of production of Th2 cytokine (IL-5) after treating with naringenin or morin in PMA+ionomycin treated EL-4 cells (Fig. 7).
Histopathological changes in the lung: The findings showed asthma-induced group showed dramatic (p < 0.05) elevation in inflammatory cell infiltration relative to control mice (without induction of asthma). Nonetheless, treatment with naringenin or morin significantly reduced penetration and aggregation of inflammatory cells in the lung tissue relative to the group induced with asthma (p < 0.05; Fig. 8).
DISCUSSION
Chronic allergic asthma is an airway inflammation characterized by eosinophilia [1,4], inflammatory cytokine levels like IL-5 production, and oxidative stress in cells [35-37]. Beneficial effects of citrus flavanoids are widely reported with several anti-inflammatory, antioxidant, antimicrobial and anti-endotoxemic activities in the prevention of hepatic ischemia, cardiac hypertrophy, liver steatosis and suppression of dengue virus replication [38-44]. However, the effect of naringenin and morin on inflammatory responses to asthma by inhibiting IL-5 and ROS production in PMA+ionomycin treated EL-4 cells and corroborated naringenin or morin effects OVA-induced asthma is not yet reported.
NFAT, a transcription factor, is involved in mediating cytokine secretion by Th2 responses [7,45,46]. A previous study showed the secretion of IL-5 modulating NFAT transcription factor in asthma patients [47]. Our results consistently showed the production of IL-5 in PMA+ionomycin treated EL-4 cells involving the regulation of NFAT (Fig. 1). Downregulation of the expression of the nuclear NFAT protein was significantly inhibited when treated with naringenin or morin (100 μM), as shown in Fig. 3.
It has been shown that suppression of HO-1 expression appears to be a novel therapeutic modulation of inflammation through the Nrf2 and MAPK signalling pathways [48]. Convincingly, our findings showed that PMA+ionomycin treated in EL-4 cells upregulated production of HO-1 with the treatment of naringenin and morin. Interestingly, naringenin showed increased HO-1 expressions in EL-4 cells through the transcriptional Nrf2, associated with P13K/Akt or Erk/JNK signalling pathways (Fig. 4). Our findings showed that HO-1 production was upregulated by naringenin and morin through similar signalling pathways. However, both of them (naringenin or morin) showed dramatic downregulation of the production of IL-5 through the NFAT transcription factor (Figs. 3, 4, and 7), thus suggesting that increased expression of HO-1 might suppress inflammation caused in asthma by inhibiting IL-5 production.
Naringenin and morin have anti-inflammatory and antioxidant effects on OVA-induced asthma models. However, the present study provided evidence of these effects using ovalbumin-induced airway inflammation in a mouse model of asthma to corroborate the effects of naringenin or morin on PMA+ionomycin induced EL-4 cell outcomes showing a substantial reduction in airway inflammation characteristics which include inflammatory responses, cytokine production along with suppression of ROS production.
Additionally, chronic allergic asthma involves elevation in BALF eosinophils and subsequent lung infiltration [4]. Our in vivo findings showed that naringenin or morin decreased eosinophilic penetration in the lungs and decreased their BALF numbers, respectively, suggesting that they possess anti-inflammatory effects in allergic asthma.
It is reported in several studies that suppression of Akt and p38 phosphorylation is the significant pathway involved in immune responses that helps in the regulation of genes related transcriptional functions of pro-inflammatory cytokines such as IL-(3, -4, -5, -13) and TNF-α [49]. Therefore, in our study, we demonstrated that naringenin or morin suppressed the phosphorylation of Akt/p38 pathways, thereby significantly inhibiting the Th2 cytokines production IL-(5, -4, and -13) and TNF-α in the BALF and lungs, respectively, which were raised during inflammation of airway in an allergy-induced asthma mouse model. One of the allergic reaction pathways is an elevation in the secretion of IL-5 in airway inflammation that causes eosinophil infiltration into the lungs [50]. Consequently, allergic asthma can be known as Th2-mediated immune disease. Therefore, these findings indicate that naringenin or morin play a vital role in suppressing eosinophils recruitment into the lungs by inhibiting Th2-medicated cytokine pathways.
Our research showed that naringenin or morin activated the Heme oxygenase-1 and Nrf2 expression through the signalling pathways involving the phosphorylation of P13K/Akt or ErK/JNK. Such processes, therefore, contributed a significant role in suppressing NFAT expression and thus inhibiting IL-5 production in the PMA+ionomycin induced EL-4 cells. Collectively, downregulation of NFAT by Nrf2 activation had contributed to the inhibitory outcome of naringenin or morin on the generation of ROS and IL-5 by PMA+ionomycin treatment EL-4 cells thus confirming the stimulation of HO-1 expression could suppress asthmatic inflammatory responses by inhibiting IL-5 secretion and ROS formation. Our findings conclude that naringenin or morin can protect anti-inflammatory and antioxidant activity against allergic inflammation in asthma. Therefore, naringenin or morin could be developed as therapeutic agents or functional foods that help in allergic asthma prevention and treatment.
 XML Download
XML Download