

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Extraintestinal pathogenic Escherichia coli (ExPEC) causes various infections, including urinary tract infection (UTI), sepsis, and neonatal meningitis [1, 2]. The mechanisms underlying ExPEC transmission and the selection of resistant clones are poorly understood.
Among multidrug-resistant ExPEC strains, the most frequent sequence type (ST) is ST131, which is globally disseminated and resistant to multiple antibiotics [3, 4]. Recently, ST1193, which is resistant to fluoroquinolones, has spread rapidly [4]. ExPEC strains have virulence factors (VFs), including adhesion molecules, iron acquisition systems, invasion proteins, and toxins, which facilitate infection by allowing bacterial cells to migrate into and multiply within the host. Several ExPEC VF genes, including pap, vat, kpsMII, ibeA, and clbB/N, are significantly associated with the dissemination of ExPEC isolates [5, 6]. The yersiniabactin siderophore receptor encoded by fyuA is involved in the efficient uptake of iron from the bloodstream and invasion of the bloodstream from the urinary tract [7]. Pneumonia-specific E. coli isolates carry higher proportions of the VF genes sfa/foc, papGIII, hlyC, cnf1, and iroN than bacteremia isolates [8]. The gene cnf1 encodes cytotoxic necrotizing factor 1, a toxin associated with sepsis severity, whereas the presence of fyuA is associated with mortality. Further, the presence of P fimbriae is associated with improved bacterial survival [9, 10]. The presence of hek/hra may predict clinical outcomes, as one study showed an association between the presence of these genes and mortality in newborns [11].
ExPEC strains involved in UTIs are believed to express a diverse repertoire of VFs to colonize and infect the urinary tract in an ascending manner [1]. However, the role of VFs in the pathogenesis and clinical outcomes of E. coli bacteremia remains to be investigated. We hypothesized that some VFs of ExPEC are involved in bacterial movement into the bloodstream in UTI patients, resulting in urosepsis. Therefore, we compared the repertoire of STs and VF genes between isolates from urine and blood from UTI patients using whole-genome sequencing (WGS) analysis.
Go to : 
MATERIALS AND METHODS
Patients and definitions
We conducted a prospective study in the 855-bed Hanyang University Hospital, Seoul, Korea. We consecutively recruited hospitalized UTI patients with E. coli isolates from blood and/or urine (N=40) and with E. coli isolates from urine alone (N=40), between March 2019 and May 2020. Patients with UTI were defined as those who met all of the following criteria: (i) fever (body temperature ≥37.8°C), (ii) pyuria (≥5–9 white blood cells per high-power field), (iii) clinical symptoms or signs relevant to UTI as judged by an infectious disease specialist, and (iv) absence of other medical conditions that can cause fever or pyuria. We collected clinical data, including demographic data (age and sex), time lag between fever or UTI symptoms (dysuria, frequency, urgency, or nocturia) and bacterial culture sampling, history of antibiotic use prior to bacterial culture sampling, and existence of underlying urinary tract abnormalities. Antibiotic regimens were considered concordant, if they included at least one antibiotic active against the causative organisms based on in vitro susceptibility testing. The clinical factors that may have influenced the culture results of blood and urine specimens, are listed in Table 1. No significant differences were found in these clinical factors. This study was approved by the Hanyang University Hospital Institutional Review Board, with waiver of informed consent (IRB N 201905054).
Table 1
Clinical factors possibly influencing the culture results of blood and urine specimens
![]()
Bacterial culture and antimicrobial susceptibility testing
Blood culture bottles were incubated in the BACT/ALERT VIRTUO System (bioMérieux, Marcy l’Étoile, France). Urine specimens were inoculated on blood agar and MacConkey agar plates, and the plates were incubated at 35°C for 24 hours. Bacterial species were identified by matrix-assisted laser desorption/ionization (MALDI) time-of-flight mass spectrometry using the MALDI Biotyper system and related software (version 2.3, Bruker Daltonics, Bremen, Germany). Antimicrobial susceptibility was tested using the MicroScan Walkaway system (Beckman Coulter, Brea, CA, USA), and the results were interpreted according to the Clinical and Laboratory Standards Institute (CLSI) guidelines, with intermediate isolates designated as resistant [12]. All isolates were maintained in 20% skim milk at –80°C.
WGS analysis
Bacterial isolates cultured on MacConkey agar plates were sent to Macrogen (Seoul, Korea) for WGS that was performed using the Illumina system (Illumina, Inc., San Diego, CA, USA). De novo assembly and assembly validation were performed for all 80 isolates. WGS data were analyzed using the Center for Genomic Epidemiology website (http://www.genomicepidemiology.org/), and multilocus ST (MLST), serotype, fimH type, antimicrobial resistance gene repertoire, and VF repertoire were determined. The presence of several virulence traits was determined using the VF analyzer pipeline and VF database (http://www.mgc.ac.cn/VFs/) [13]. Virulence scores were calculated as the sum of VFs present in each isolate [5].
Statistical analysis
The chi-square test (Fisher’s exact test) was used to compare clinical factors, ST, antimicrobial resistance rates between blood and urine groups, and the Kruskal–Wallis test was used to compare virulence scores according to ST. Data was analyzed using SPSS 26 (SAS Institute Inc., Cary, NC, USA), and P<0.05 was considered statistically significant.
Go to :
RESULTS
Molecular epidemiological characteristics
The MLST results revealed a diverse population comprising 18 STs (Table 2). The most frequent STs were ST131 (N=19, 24%), ST95 (N=16, 20%), ST69 (N=11, 14%), and ST1193 (N=9, 11%). We detected five ST73 isolates and two ST127 isolates. There were no significant differences in ST frequencies between the blood and urine specimens. ST12 was more frequently found in urine specimens than in blood specimens (P=0.04), but only four isolates were obtained.
Table 2
Number of isolates (%) according to MLST in blood and urine specimens
| ST-fimH-serotype | Blood (N=40) | Urine (N=40) | P |
|---|---|---|---|
| ST131 | 8 (20) | 11 (28) | 0.431 |
| ST131-fimH30-O25:H4 | 6 (15) | 8 (0.556) | 0.556 |
| ST131-fimH41-O16:H5 | 2 (5) | 2 (1.000) | 1.000 |
| ST131-fimH99-O25:H4 | 0 (0) | 1 (0.314) | 0.314 |
| ST95 | 9 (23) | 7 (18) | 0.576 |
| ST95-fimH27-O2:H4 | 6 (15) | 3 (0.288) | 0.288 |
| ST95-fimH27-O2:H7 | 1 (3) | 1 (1.000) | 1.000 |
| ST95-fimH41 | 2 (5) | 2 (1.000) | 1.000 |
| ST95-fimH54 | 0 (0) | 1 (0.314) | 0.314 |
| ST69-fimH27 | 8 (20) | 3 (8) | 0.105 |
| ST1193 | 4 (10) | 5 (13) | 0.723 |
| ST1193-fimH64-O75:H5 | 3 (8) | 4 (0.692) | 0.692 |
| ST1193-fimH64-O18:H5 | 1 (3) | 1 (1.000) | 1.000 |
| ST73 | 3 (8) | 2 (5) | 0.644 |
| ST12 | 0 (0) | 4 (10) | 0.040 |
| ST38 | 1 (3) | 2 (5) | 0.556 |
| ST127 | 0 (0) | 2 (5) | 0.152 |
| ST357 | 1 (3) | 1 (3) | 1.000 |
| Others* | 6 (15) | 3 (8) | 0.288 |
![]()
FimH typing revealed that fimH27 isolates were the most frequent (N=22, 28%), followed by fimH30 isolates (N=14, 18%). We also determined fimH-OH serotype clonal groups among the ST isolates. Among the 19 ST131 isolates, the ST131-fimH30-O25:H4 group was predominant (N=14, 74%). Among the 16 ST95 isolates, ST95-fimH27 isolates were the most frequent (N=11, 69%). All nine ST1193 isolates were fimH64, whereas this type was not found among non-ST1193 isolates.
Antimicrobial resistance patterns and antimicrobial resistance genes
The ampicillin and ampicillin/sulbactam resistance rates were 72.5% and 40.0%, respectively, for blood isolates and 75.0% and 45.0%, respectively, for urine isolates. In addition, 20.0% of blood isolates and 27.5% of urine isolates were extended spectrum β-lactamase (ESBL) producers. We found no significant differences in antimicrobial resistance rates between blood and urine isolates (Table 3). However, ST131 and ST1193 isolates showed higher resistance rates than the other isolates. The cefotaxime, ceftazidime, aztreonam, and cefepime resistance rates of ST131 isolates (78.9%) were higher than those of non-ST131 isolates (0%–12.0%). Among the 19 ST131 isolates, 15 carried ESBL-associated genes including CTX-M-15, CTX-M-27, CTX-M-55, and CTX-M-14. CTX-M-14 genes were detected in ST95, ST38, and ST457, and the DHA-1 gene was detected in one ST12 isolate (Table 4). Of the 80 E. coli isolates, 27 (33.8%) were resistant to fluoroquinolone. Among them, the most prevalent clonal group was ST131-fimH30, followed by ST1193-fimH64. All ST1193 isolates were resistant to ciprofloxacin. Quinolone resistance analysis revealed quinolone resistance-determining region (QRDR) point variants in gyrA, parC, and parE. All 27 fluoroquinolone-resistant isolates carried the variations S83L and D87N or D87Y in gyrA;S80I and/or E84V in parC were detected in 24/27 (89%) isolates, and I529L, L416F, and S458A in parE were detected in 23/27 (78%) isolates.
Table 3
Antimicrobial resistance rates (%) according to ST and specimen type
![]()
Table 4
Antimicrobial resistance mechanisms of ESBL-producing and fluoroquinolone-resistant isolates according to ST-fimH-serotype
![]()
Prevalence of VFs
Reference strains were uropathogenic E. coli (UPEC) strains (E. coli CFT073, E. coli O25b:H4-ST131, and E. coli VR50), adherent and invasive E. coli strains (E. coli UM146 and E. coli O83:H1 strain NRG 857C), neonatal meningitis E. coli (NMEC) strains (E. coli O7:K1 strain CE10 and E. coli O45:K1:H7), and an enterohemorrhagic E. coli strain (E. coli O157:H7 strain EDL933). In total, 56 VFs were examined and categorized as adhesion molecules, autotransporter systems, invasion proteins, or toxins (Table 5). All isolates carried E. coli common pilus, hemorrhagic E. coli pilus, and EaeH. The frequencies of individual VFs according to ST ranged from 91%–100% for type I fimbriae and 64%–100% for P fimbriae. F1C or S fimbriae were not detected in prevalent isolates, such as ST131, ST95, ST69, and ST1193. Among VFs associated with autotransporter systems, the temperature-sensitive hemagglutinin gene (tsh) was detected in all ST131 and ST1193 isolates, 6% of ST95 isolates, and 0% of ST69 isolates. Among invasion VFs, ibeA, associated with neonatal meningitis, was detected in only four (5%) isolates, including ST357, ST372, and ST998 isolates. Iron uptake systems, such as the sitABCD pump and chuA system, were detected in nearly all isolates (98%) (Table 5). Yersiniabactin, aerobactin, and salmochelin siderophores, which are indirect iron uptake systems, were detected in 99%, 63%, and 29% of the isolates, respectively.
Table 5
Proportions (%) of VFs according to ST and specimen type
![]()
In ST131 and ST1193 isolates, yersiniabactin and aerobactin siderophores were more frequent than salmochelin siderophores. In contrast, salmochelin siderophores were more frequent (68%) in other isolates than ST131 (5%) and ST1193 (0%) isolates. Several toxin genes were detected in UTI-associated isolates. The hlyE gene was detected in all isolates. The usp and senB genes were predominant in 78% and 55%, respectively, of the isolates. The alpha-hemolysin genes, hlyA, hlyB, and hlyD, were detected in 35% of the isolates. These toxin genes were more frequent in the ST131 group (53%) than in the non-ST131 group (30%), but the difference was not significant (P=0.07). The cnf1 gene was more frequent in the ST131 group (53%) than in the non-ST131 groups (20%) (P<0.05).
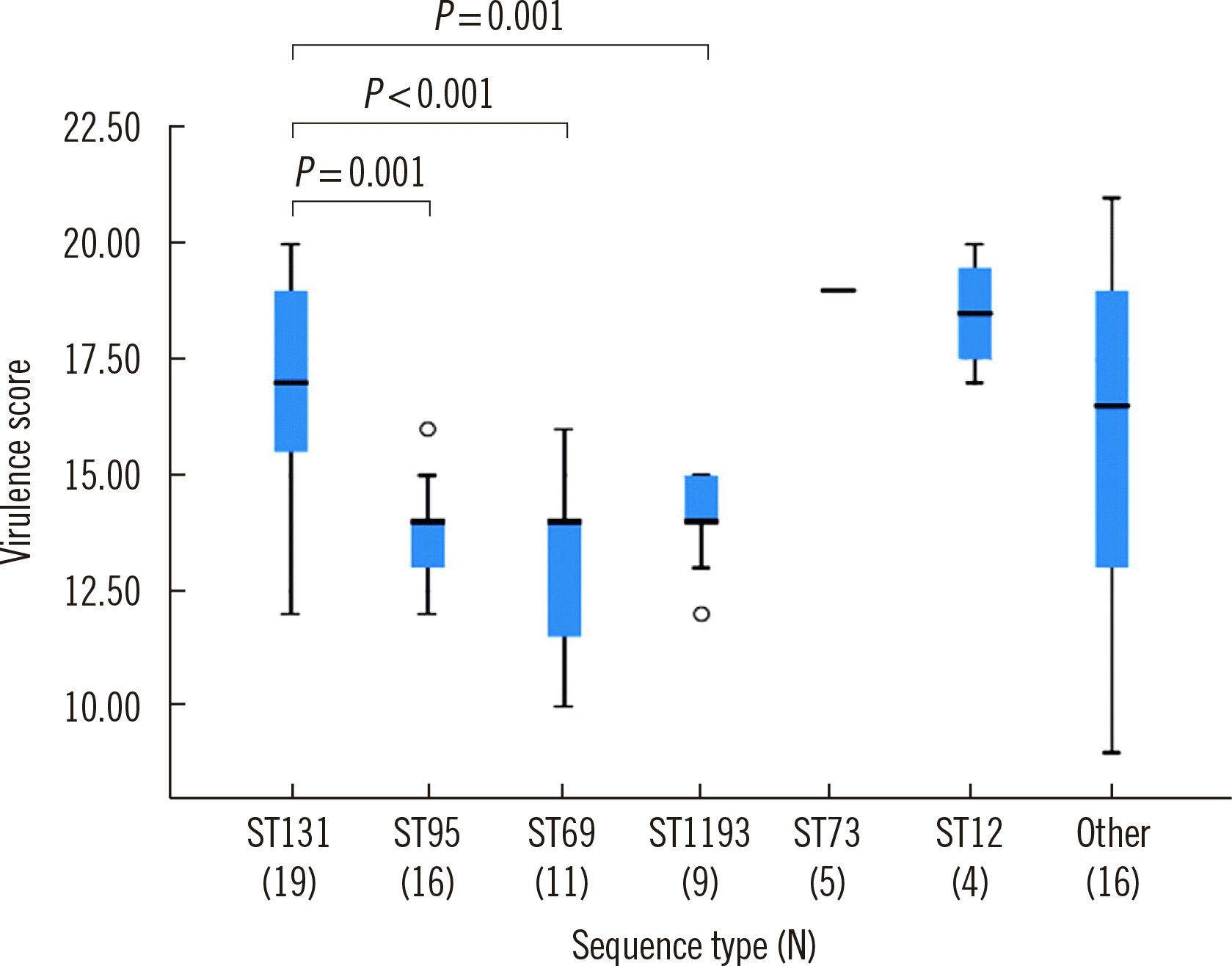
The median virulence score for all 80 isolates was 15 (range, 0–21). Virulence scores differed significantly among ST groups (Fig. 1). ST12 and ST73 isolates had high virulence scores, ranging from 17–20.
Go to :
DISCUSSION
We hypothesized that we could define specific VFs of E. coli linked to bacterial movement from the urinary tract to the circulation. However, virulence properties as well as molecular epidemiological traits and antimicrobial resistance patterns showed no significant differences between blood and urine isolates from our cohort of UTI patients, suggesting that specific VFs of E. coli do not contribute to bacterial dissemination from the urinary tract to the circulation. However, specific isolates, such as ST131, carried more VF genes and exhibited higher antimicrobial resistance than the other isolates evaluated.
We classified isolates according to ST-fimH-OH serotype clonal groups using WGS. The ST131-fimH30-O25:H4 type was the most predominant and was highly resistant to several antibiotics, including β-lactams and fluoroquinolones. Among the 14 ST131-fimH30-O25:H4 isolates, 12 produced CTX-M type EBSLs and carried variations in gyrA, parC, and parE. The next three most prevalent ST-fimH isolates were ST95-fimH27, ST69-fimH27, and ST1193-fimH64. ST131 is the predominant E. coli lineage among multidrug-resistant ExPEC isolates worldwide [14]. Most ST131 isolates are fluoroquinolone-resistant, many are co-resistant to aminoglycosides and cotrimoxazole, and a minority produce ESBLs, such as CTX-M enzymes [5, 14]. In our study, 78.9% of the ST131 isolates produced CTX-M ESBLs and were resistant to fluoroquinolones, and 52.6% of these were co-resistant to aminoglycosides and cotrimoxazole. After the rapid expansion of ST131 E. coli strains, fluoroquinolone-resistant ST1193 isolates emerged [4]. ST1193 strains, derived from ST14, are commonly resistant to fluoroquinolones. fimH64 is a highly specific marker for ST1193, a clonal group that may be entirely fluoroquinolone-resistant [4]. Consistent herewith, all ST1193-fimH64 isolates in our study were resistant to fluoroquinolones.
Adhesins, including fimbriae and afimbrial adhesins, play a significant role in host cell colonization [7, 15]. We detected type I fimbriae, E. coli common pilus, hemorrhagic E. coli pilus, and EaeH in all isolates. P fimbriae were detected in 80% of the isolates. ExPEC can cause sepsis and infections under very low iron availability, as this pathotype has developed many strategies for obtaining iron [16]. The sitABCD system is a membrane pump system, and the ChuA transporter enables Fe uptake directly from extracellular heme. In addition, ExPEC have siderophores, such as salmochelin, yersiniabactin, and aerobactin, which are small molecules with high affinity for Fe ions that indirectly uptake Fe [17]. Yersiniabactin contributes to the pathogenicity of UPEC during urinary tract colonization [18, 19]. In our study, 98% isolates had yersiniabactin, the sitABCD system, and the chuA transporter. Aerobactin receptors are substantially more efficient at capturing Fe than enterobactin receptors [7]. In this study, 74% and 89% of the ST131 and ST1193 isolates, respectively, had the gene encoding aerobactin. An ExPEC salmochelin marker gene, iroN, was detected in 5% of the ST131 isolates, but in none of the ST1193 isolates.
Some of the most frequently detected toxin genes in ExPEC are hlyA, hlyD, hlyF, cdtB, tsh, sat, pic, vat, and astA [7]. hlyF, cdtB, and tsh have been detected in NMEC strains [20]. While all isolates carried hlyE in this study, alpha-hemolysins encoded by hlyABCD were detected in only 53% of ST131 and 27% of ST69 isolates. The cnf1 gene was detected in 53% of the ST131 isolates. The presence of these genes in ST131 isolates suggests that they exhibit high virulence and toxicity compared with non-ST131 isolates. The IbeA protein recognizes surface receptors on brain capillary endothelial cells, allowing the pathogen to invade the nervous system and cause neonatal meningitis [21–23]. Therefore, ibeA is a representative VF gene in NMEC strains. In our study, three of each of ST357, ST372, and ST998 isolates (5%) carried ibeA. Major UTI pathogens, such as ST131 and ST1193, may not carry ibeA and therefore, do not cause meningitis in neonates.
ST131 isolates carried more VF genes, including tsh and espC, than non-ST131 isolates. The VF genes in ST131 isolates may play a role in pathogenesis. In a previous study, tsh was present in >50% of avian pathogenic E. coli isolates, 4.5% of UPEC isolates, and 11.5% of NMEC isolates [15]. In this study, all ST131 and ST1193 isolates among UTI-associated E. coli isolates carried tsh. Although the function of the temperature-sensitive hemagglutinin encoded by tsh in human infections is unclear, in Bacteroides fragilis, this protein contributes to abscess formation in intra-abdominal infections [24]. ST127 isolates can cause bacteremia in adults and are highly virulent in experimental models of invasive E. coli infection. Therefore, ST127 isolates should be monitored because of their greater pathogenic potential than more widely recognized clones, including ST73, ST95, and ST131 [25]. Two ST127 isolates were recovered from urine in this study and had high virulence scores of 20 and 17.
ST131 isolates reportedly had a higher virulence potential than other E. coli types in a murine sepsis model [5]. Similarly, in this study, ST131 isolates had higher virulence scores than ST1193, ST95, and ST69 isolates (P<0.05). Shah, et al. [26] suggested a significant association between VFs and antimicrobial resistance in UPEC. Consistent herewith, we found that ST131 isolates of UTI patients had several VFs and a high resistance rate to several antibiotics.
Our study had some limitations, mainly related to the relatively small specimen size. Because of the limited number of E. coli bacteremia isolates reported, larger studies are needed to accurately determine the VF composition of these isolates in relation to their clonal characteristics. Further studies are needed to clarify the association between virulence and antibiotic resistance in urosepsis-related E. coli strains. Second, WGS analysis and web-based assessment are subject to experimental error. Third, the association between VFs and disease severity could not be clarified in this study.
In conclusion, E. coli bacteremia isolates from UTI patients were analyzed for a broad range of VFs, molecular backgrounds, and antibiotic resistance genes using WGS. We found no STs and VFs associated with bacteremia in WGS data of E. coli isolated from UTI patients. ST131, a pandemic multidrug-resistant clone present in both blood and urine specimens, was the most frequent ST and carried more VF genes, especially tsh and espC, than non-ST131 isolates.
Go to :
 XML Download
XML Download