

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Identifying the causes of encephalitis can be challenging because various bacterial, viral, fungal, and parasitic infections can manifest with overlapping symptoms and signs. Pathogen-specific serological, PCR, and culture-based analyses are expensive, time-consuming, and lack standardization [1]. Recently, metagenomic next-generation sequencing (NGS) has been used to identify unusual or novel pathogens, including those causing meningoencephalitis. For example, Leptospira, a spirochaete, and Balamuthia mandrillaris, a free-living amoeba, have been identified in patients with meningoencephalitis by metagenomic NGS [1, 2]. Despite the several advantages of this approach, the detection of unusual protozoan infections by NGS within a reasonable timeframe is not always feasible for routine practice owing to the need for specialized equipment, and the time and expertise required for bioinformatic analyses. As an alternative, thymine–adenine (TA) cloning is more feasible and affordable than NGS. Here, we describe a patient with encephalitis caused by B. mandrillaris that was identified using the TA cloning method. To the best of our knowledge, this is the first report on B. mandrillaris gene detection in Korea. Cloning was performed using universal 18S rRNA gene primers that have been widely applied in eukaryotic metagenomic studies [3]. We initially identified all protozoal species in the brain sample, because B. mandrillaris infection was unknown in Korea at the time and was not initial-ly considered.
Go to : 
MATERIALS AND METHODS
Brain biopsy sample from a patient with encephalitis
A 50-year-old man without a history of neurological disease was referred to the Department of Neurosurgery, Eulji University Hospital, Daejeon, Korea, for evaluating an intracranial lesion visible on his brain magnetic resonance imaging (MRI) scans. The patient had a three-day history of mild speech disturbance and right-sided hemiparesis after a focal seizure. He did not chew tobacco, drink alcohol, or use drugs and showed no signs of infection, such as fever, leukocytosis, and meningeal irritation. A brain MRI revealed a mass in the cortex of the left parietal lobe (Fig. 1A, B). An open biopsy of the brain performed immediately upon arrival suggested amebic infection in the brain with inflammatory cell infiltration and necrotizing vasculitis. Numerous amebic trophozoites (20–25 μm) were identified by microscopy (Fig. 1C, D), and anti-Entamoeba histolytica and anti-Toxoplasma gondii IgM antibody titers were positive and negative, respectively. Laboratory findings were normal. An initial laboratory evaluation revealed a white blood cell count of 6.65×109/L, hemoglobin of 151 g/L, serum sodium level of 138 mmol/L, and serum glucose level of 7.05 mmol/L. A diagnosis of intracranial amebiasis was made based on the pathological findings, and the patient was treated with metronidazole, amphotericin B, and sulfadiazine/pyrimethamine. However, drowsiness and progressive hemiparesis seven days after the open biopsy indicated a deteriorated neurological status. Emergency MRI showed multiple new lesions on the opposite site of the brain and increased intracranial pressure, which was reduced by decompressive craniectomy.
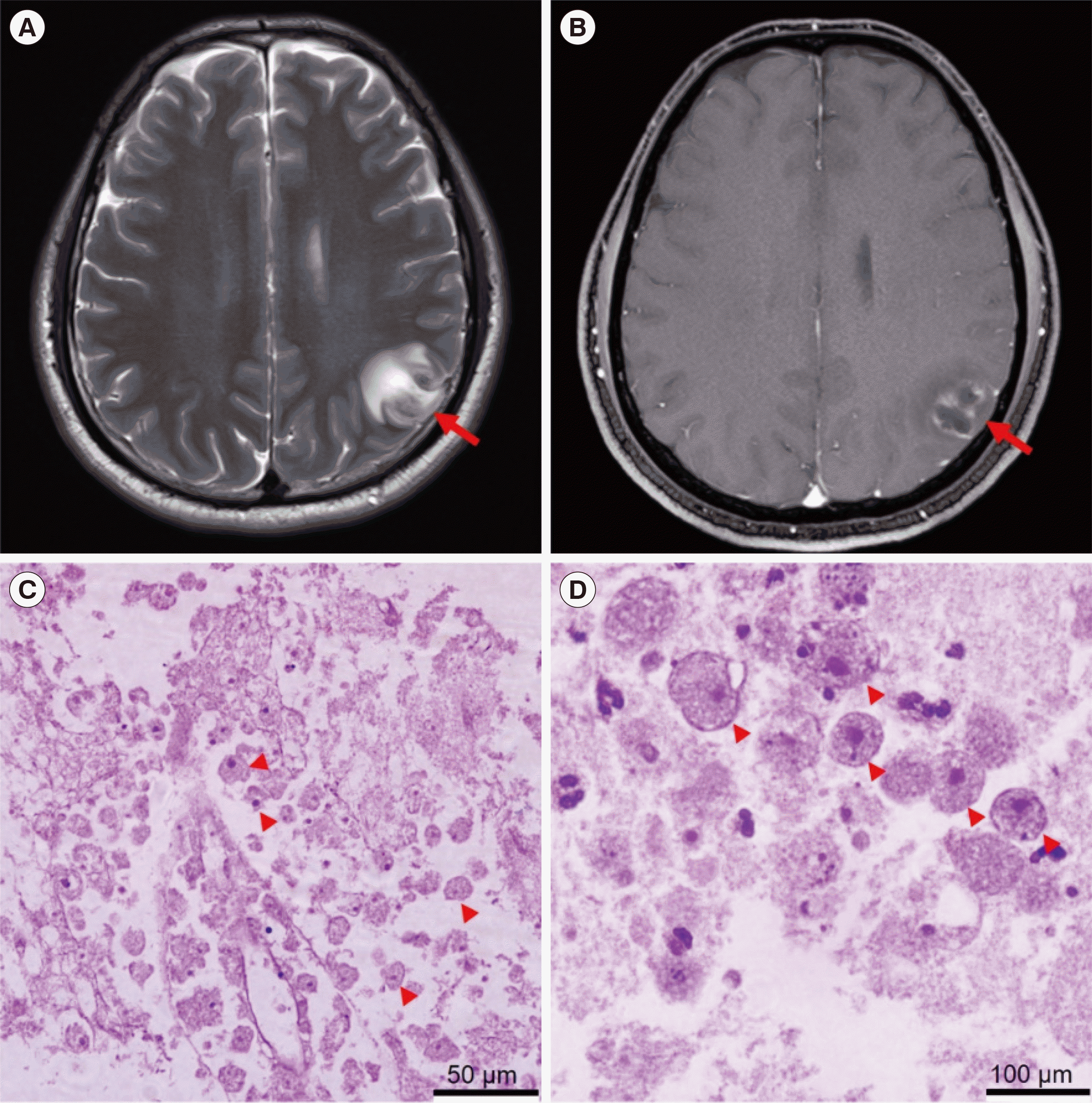 | Fig. 1Magnetic resonance image (MRI) and brain biopsy. T2-weighted MRI showing (A) a lesion (~21 mm×18 mm) with an irregular, marginated, heterogeneous dark signal and (B) marginal thin-rim enhancement of gadolinium at the left parietal cortex with surrounding edema (red arrows). (C, D) Hematoxylin and eosin staining of brain biopsy shows numerous amoebic trophozoites (20–25 μm) in the hemorrhagic necrosis background (arrowheads). Magnification: ×200 (C), ×1,000 (D).
|
The study protocol was approved by the Institutional Review Board of Yonsei University Sever-ance Hospital, Seoul, Korea (approval No. 4-2020-0201), and the patient provided written in-formed consent.
DNA extraction
A paraffin block of the sample was prepared immediately after the biopsy (February 21, 2020 at Eulji University Hospital). Two weeks after the biopsy, the paraffin block was transferred to the laboratory at Yonsei University College of Medicine, and DNA extraction, TA cloning, and PCR were performed on the same day. DNA was extracted from paraffin-embedded brain biopsy samples using the Exgene Formalin-Fixed, Paraffin-Embedded Tissue DNA Kit (GeneAll, Seoul, Korea) in accordance with the manufacturer’s instructions and stored at 4°C until analyses.
TA cloning
We amplified protozoan 18S rRNA gene sequences by PCR using two sets of primers targeting the V4-5 and V9 regions of 18S rRNA, which are typically used in eukaryotic metagenomics as they amplify the 18S rRNA of nearly all types of eukaryotic species [3]. Thus, this procedure can discriminate the 18S rRNA gene sequences of patient and protozoan DNA. The 18S rRNA gene V4-5 and V9 regions of any eukaryotic species in the sample were cloned using the TOPcloner TA Kit (Enzynomics, Daejeon, Korea), in accordance with the manufacturer’s instructions with the following primers: V4-5 region, V4-1 forward 5´-GCGGTAATTCCAGCTC-3´ and V4-4 reverse 5´-GCCMTTCCGTCAA-TTCC-3´; V9 region, 1391f 5´-GTACACACCGCCCGTC-3´ and EukBr 5´-TGATCCTTCTGCAGGTTCACCTAC-3´ [3, 4]. The reaction conditions were as follows: initial denaturation for 5 minutes at 95°C; 34 amplification cycles comprising denaturation for 30 seconds at 95°C, annealing for 30 seconds at 55°C, and extension for 45 seconds at 72°C; and final extension for 5 minutes at 72°C. The following day, recombinant colonies were cultured overnight in Luria–Bertani broth with ampicillin, and the recombinant plasmids from the recombinant clones were extracted using the Exprep Plasmid SV Mini Kit (GeneAll, Seoul, Korea). Forty-one plasmids recombinant with 18S rRNA V4-5 regions and 63 plasmids recombinant with 18S rRNA V9 re-gions were sequenced at Bionics (Seoul, Korea) using the M13F(-20) primer. Homology was de-termined using sequences from the National Center for Biotechnology Information (NCBI) Basic Local Alignment Search Tool (BLAST, https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Phylogenetic analysis
The DNA sequences were compared with nucleotide sequences available in GenBank (NCBI). A phylogenetic tree was constructed using Molecular Evolutionary Genetics Analysis version X (MEGA-X) software (The Pennsylvania State University, PA, USA) [5].
Conventional PCR for specific protozoal infection
Conventional PCR was performed to detect the protozoa Entamoeba histolytica, Toxoplasma gondii, Naegleria fowleri, and Acanthamoeba spp. based on previous studies [6-9]. N. fowleri-specific PCR was performed using the following primer set to amplify the amplicon of predicted size 183 bp: Nae3-F (5´-CAAACACCGTTATGACAGGG-3´) and Nae3-R (5´-CTGGTTTCCCTCACCTTACG-3´) [6]. Entamoeba histolytica-specific nested PCR was per-formed using the following primer sets to amplify the amplicon of predicted size 270 bp: Fw1 (5´-GCAGAAAAAAATAATAATAAC-3´) and Rv1 (5´-TTCATTTGTTTTACTTTCA-3´) for the first PCR and Fw2 (5´-CGCCCGCCGCGCGGCCGCGGCCGGCCGGGGGCACGCGGCGGCAGAAAAAAA-TAATAATAAC-3´) and Rv2 (5´-TTCATTTGTTTTACTTTCA-3´) for the subsequent PCR [7]. Toxoplasma gondii-specific PCR was performed using the following primer set to amplify the amplicon of predicted size 469 bp: Tg1 (5´-AAAAATGTGGGAATGAAAGAG-3´) and Tg2 (5´-ACGAATCAACGGAACTGTAAT-3´) [8]. Acanthamoeba-specific PCR was performed using the following primer set to amplify the amplicon of predicted size 463 bp: JDP1 (5´-GGCCCAGATCGTTTACCGTGAA-3´) and JDP2 (5´-TCTCACAAGCTGCTAGGGGAGTCA-3´) [9]. To confirm the TA cloning results, B. mandrillaris-specific PCR was performed using the following primer sets to generate products of pre-dicted sizes 1075, 230, and 170 bp, respectively: 5′Balspec16S (5´-CGCATGTATGAAGAAGACCA-3´) and 3´Balspec16S (5´-TTACCTATATAATTGTCGATACCA-3´); 5´Balspec 16S (5´-CGCATGTATGAAAGAAGACCA-3´) and Balspec 16Sr 610 (5´-CCCCTTTTTAACTCTAGTCATATAGT-3´); and BalaF1451 (5´-TAACCTGCTAAATAGTCATGCCAAT-3´) and BalaR1621 (5´-CAAACTTCCCTCGGCTAATCA-3´) [10-12].
Go to :
RESULTS
Four days after the brain biopsy, the protozoan species in the brain lesion was identified as B. mandrillaris by TA cloning. However, the patient had already succumbed to brain death and died four weeks later.
The pathogen-specific PCR results ruled out E. histolytica, N. fowleri, Acanthamoeba spp., and T. gondii infections based on the absence of positive bands (Fig. 2A). Simultaneous TA cloning was performed using general eukaryotic primers to detect unexpected protozoan infections because we did not consider B. mandrillaris infection.
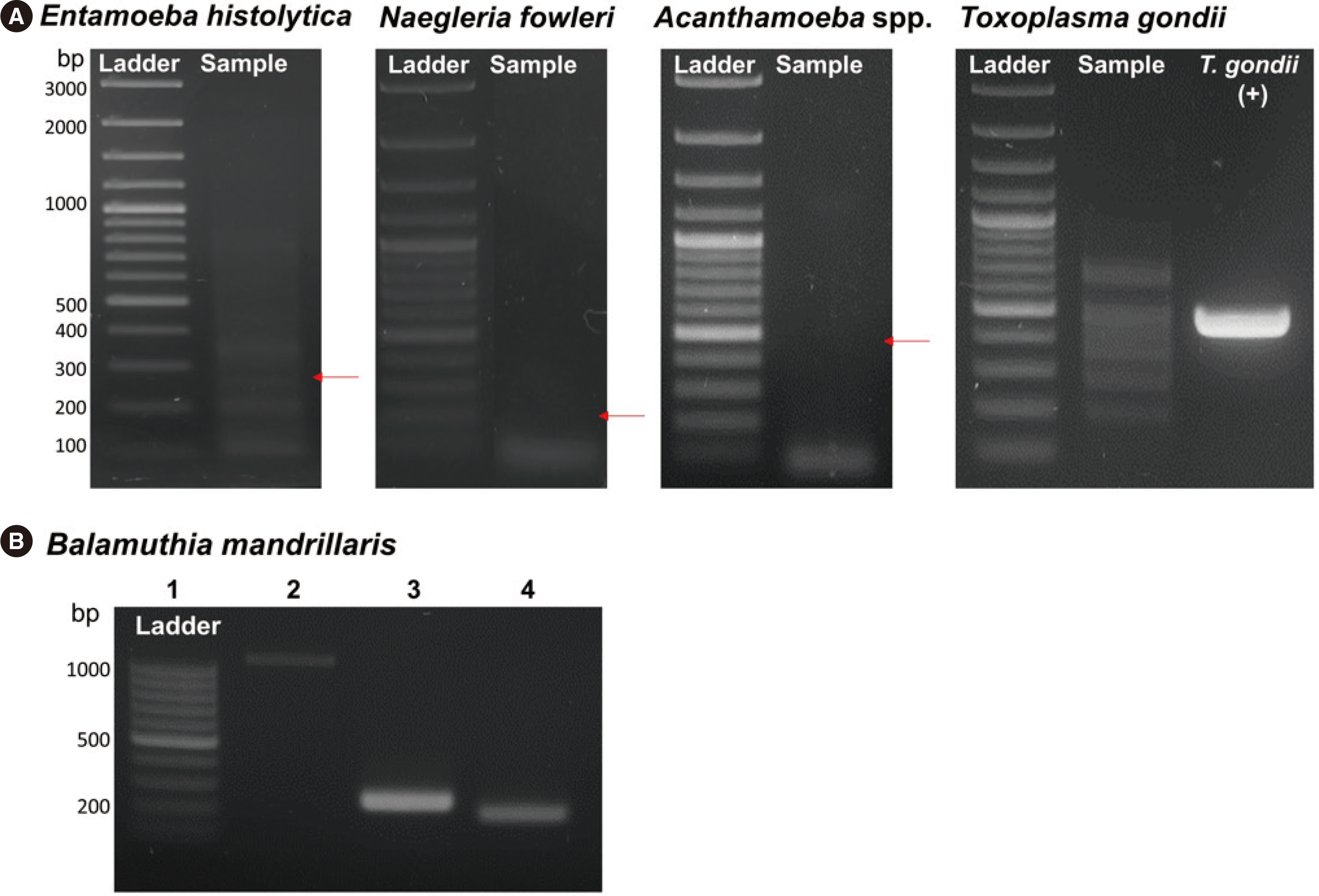 | Fig. 2Pathogen-specific PCR targeting (A) Entamoeba histolytica, Naegleria fowleri, Acanthamoeba spp., Toxoplasma gondii, and (B) Balamuthia mandrillaris. Red arrows show approximate band sizes of 270, 183, and 463 bp for E. histolytica, N. fowleri, and Acanthamoeba spp., respectively. Toxoplasma gondii DNA (469 bp) was used as the positive control. Balamuthia mandrillaris-specific PCR using the patient DNA was performed with three sets of primers. Lane 1: 100 bp ladder. Lanes 2, 3, and 4: 5′Balspec 16S and 3′Balspec16S, 5′Balspec 16S and Balspec 16Sr 610, and BalaF1451 and BalaR1621 primers, respectively.
|
The homology analysis using NCBI BLAST showed that the 18S V4-5 and V9 regions of the protozoan pathogen in the sample exhibited 99.2% and 100% nucleotide identity with those of B. mandrillaris KF874819 and KT175741 in GenBank, respectively (Table 1).
Table 1
Molecular identification (nucleotide identity) of the protozoa from the patient and previously published protozoan DNA sequences that required differential diagnoses
![]()
The DNA of the patient and B. mandrillaris was detected at different rates when cloning the 18S rRNA V4-5 or V9 variable region. This is likely related to the variation in the GC content and DNA sequence lengths in the two genes (Table 2).
Table 2
Number of recombinant colonies containing 18S rRNA genes (V4-5 and V9 regions) of Balamuthia mandrillaris and Homo sapiens and their GC content and DNA length
![]()
B. mandrillaris infection was reconfirmed using B. mandrillaris-specific PCR with three primer sets. The bands resolved at the expected points in gel electrophoresis and their DNA sequences were identical to published B. mandrillaris nucleotide sequences (Fig. 2B).
The phylogenetic tree of 18S rRNA V4-5 and V9 sequences showed that the sequence of the sample was distant from those of other protozoan candidates, including E. histolytica, N. fowleri, Acanthamoeba castellanii, and T. gondii (Fig. 3).
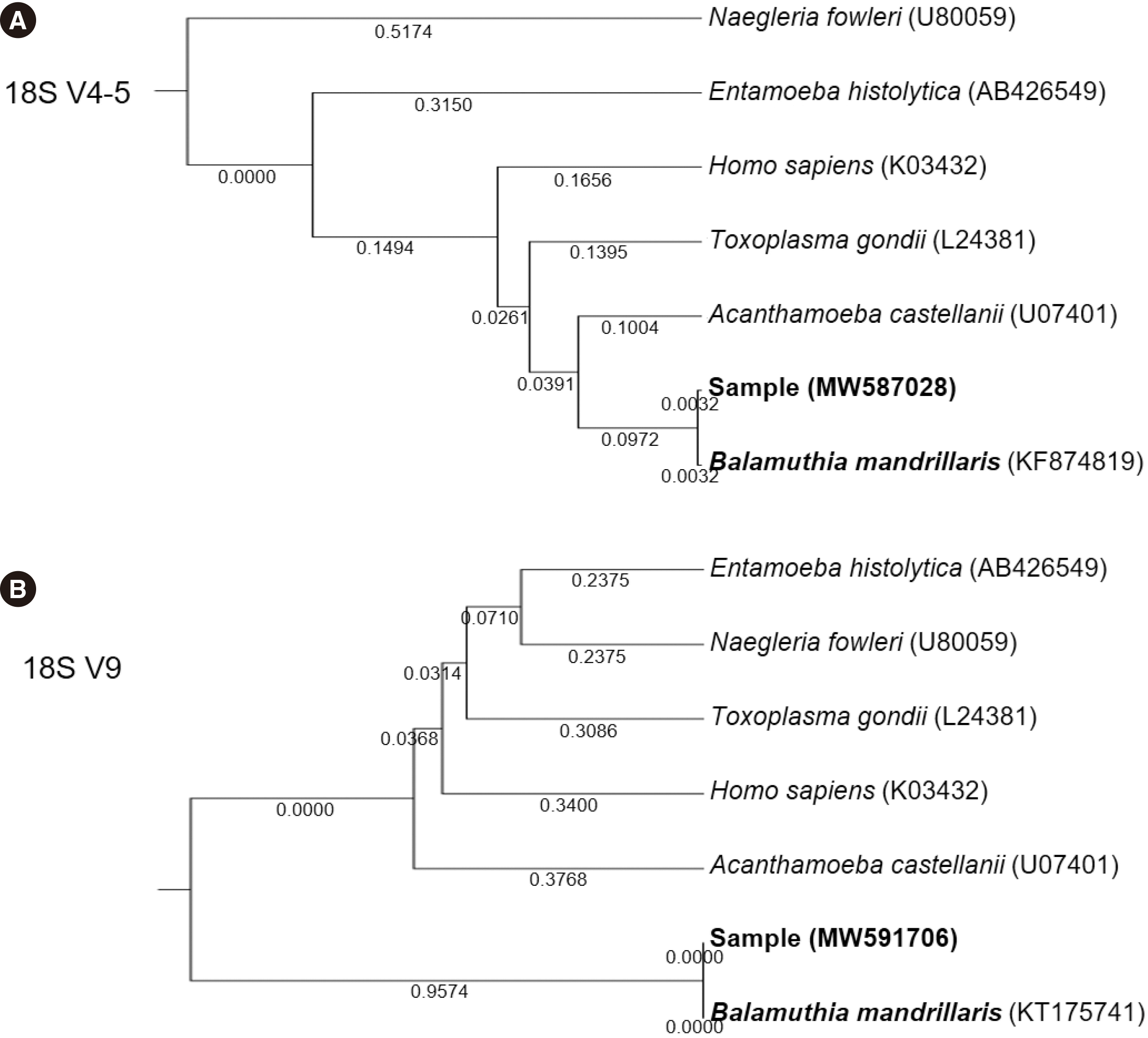 | Fig. 3Phylogenetic trees of the sequenced sample based on the (A) V4-5 region and (B) V9 region of the 18S rRNA gene. The total branch lengths of phylogenetic trees for the V4-5 and V9 regions were 0.8465 and 2.2305, respectively, computed using the maximum composite likelihood method and measured in base substitutions per site. Sequences of 18S rRNA genes of reference organisms were obtained from GenBank (National Center for Biotechnology Information).
|
Go to :
DISCUSSION
Naegleria fowleri, Acanthamoeba spp., and B. mandrillaris are established etiological agents of encephalitis. N. fowleri causes primary amebic meningoencephalitis with a fulminant disease course, and Acanthamoeba spp. and B. mandrillaris cause granulomatous amebic encephalitis [13, 14]. Among approximately 150 patients with Balamuthia amebic encephalitis reported worldwide, only 10 have survived [15]. B. mandrillaris enters the host via skin wounds or inhalation and spreads through the bloodstream to the central nervous system, resulting in fever, seizures, cranial nerve dysfunction, motor deficits, increased intracranial pressure, and altered consciousness [16, 17]. The survival rate of patients with B. mandrillaris encephalitis can be <2% or a maximum of approximately 6.7% due to its rarity, difficulty in diagnosis, and lack of effective treatment protocols [16, 18]. The US Center for Disease Control and Prevention recommends pentamidine, sulfadiazine, flucytosine, fluconazole, clarithromycin, azithromycin, and miltefosine to treat B. mandrillaris encephalitis [19-23].
To the best of our knowledge, this is the first case of B. mandrillaris encephalitis in Korea identified using a molecular (TA cloning) approach. Although a 71-year-old man in Korea was diagnosed as having B. mandrillaris encephalitis in 2019 based on microscopy findings [24], including the detection of trophozoites and cysts of B. mandrillaris as well as areas of granuloma in host tissues, this protozoan is morphologically indistinguishable from Acanthamoeba spp. [13, 15, 25]. Balamuthia culture requires feeder cells and several weeks for cell proliferation. A diagnosis should be confirmed by PCR-based detection of B. mandrillaris DNA in tissues [26]. However, this technique is not widely available given the extreme rarity of B. mandrillaris encephalitis [1].
In the present case, pathogen-specific PCR was negative for E. histolytica, N. fowleri, Acanthamoeba spp., and T. gondii. T. gondii was the only positive control because protozoal and infected tissue samples were not available in our laboratory. This is likely a common situation in other research institutions, as it is difficult to maintain positive samples for all possible infectious diseases.
We applied TA cloning in this case, because it can provide results faster than metagenomic NGS and the protozoan cells in the brain tissues detected by microscopy were enough to enable confirmation using TA cloning by separating amplified protozoan and patient 18S rRNA genes. With this approach, we identified B. mandrillaris genes in the brain biopsy sample four days after receiving the paraffin-embedded sample. Theoretically, TA cloning can generate results within 48 hours; there-fore, it should be among the diagnostic choices available to detect infectious agents of unknown etiology in clinical practice.
Cloning with the primer amplifying the 18S rRNA V9 region resulted in 35 colonies with the B. mandrillaris gene and eight colonies with the patient gene. In contrast, cloning with the 18S V4-5 primer resulted in two and 32 colonies with B. mandrillaris and human genes, respectively (Table 2). This difference is probably due to the length and the GC content of the PCR products (Table 2). The PCR product of the 18S V4-5 region of B. mandrillaris was 11% longer than that of the human gene. Furthermore, the GC content of B. mandrillaris PCR products increased from 40.8% for the V9 primer to 46.1% for the V4-5 primer, which might have reduced PCR efficiency. Various primers can be used to amplify different 18S rRNA regions during TA cloning to increase the probability of protozoa detection.
This study is limited by the fact that we identified B. mandrillaris by TA cloning only in a brain lesion. As brain tissue biopsies can cause neurological disorders, diagnosing B. mandrillaris infection in samples other than brain tissues such as the blood and cerebrospinal fluid might be more helpful for patients undergoing clinical treatment.
In conclusion, to the best of our knowledge, this is the first study to describe the molecular identification of B. mandrillaris encephalitis in Korea using TA cloning. This method should be considered for detecting infectious agents of unknown etiology in routine clinical practice.
Go to :
 XML Download
XML Download