

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Diabetes mellitus is a chronic metabolic disease characterized by abnormal insulin secretion or insulin resistance and associated hyperglycemia, in which blood glucose levels are elevated [12]. The global diabetes prevalence in 2019 was estimated to be 9.3% [3]. Over the past decade, studies on the cellular and molecular mechanisms of diabetes and its complications have revealed a close association between nutrient excess and derangements in mediators of immunity and inflammation [4]. High glucose levels have been shown to induce inflammatory cytokine secretion in both clinical and experimental settings [56].
Nuclear factor-κB (NF-κB) is a transcription factor involved in the expression of several genes, such as those regulating inflammatory and immune responses [78]. NF-κB has also been involved in the association between chronic diabetes complications, cardiovascular disease, and microvascular complications [9]. Among the toll-like receptors (TLRs), TLR2/4 has an important role in inflammation and diabetes [10]. In several inflammatory diseases, expression of TLRs and the downstream signal adapter, myeloid differentiation factor 88 (MyD88), are also increased, leading to NF-κB activation and induction of genes encoding inflammatory mediators such as cytokines and cyclooxygenase-2 (COX-2) [11].
An accumulation of evidence suggests that a chronic elevation of lipopolysaccharide (LPS) could have a role in the pathogenesis of insulin resistance in vivo. An elevated LPS concentration contributes to low-grade systemic inflammation, a central feature of obesity, insulin resistance, and type 2 diabetes [12]. TLRs can recognize various components of bacterial cell walls such as LPS, peptidoglycans, and lipopeptides. TLR4 binding to LPS activates NF-κB and leads to the production of inflammatory cytokines such as interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) [13]. Liang et al. [12] reported that metabolic endotoxemia could be involved in the pathogenesis of insulin resistance in obese and type 2 diabetes subjects and that targeting TLR4 might be beneficial in these individuals.
IL-6, a proinflammatory cytokine secreted during inflammation by monocytes and macrophages, activates lymphocytes to increase antibody production, and its expression is elevated in inflammatory lesions. TNF-α also has an important role in triggering inflammatory responses by activating macrophages and increasing the production of other proinflammatory cytokines [14].
Sirtuins (SIRTs) are members of the class III group of histone deacetylases (HDACs). Interest in these proteins is increasing because studies have shown that SIRTs are linked to age-related diseases, such as cancer, diabetes, and neurological diseases [15161718]. Among the SIRTs, SIRT3 is reported to act as an NAD+-dependent deacetylase, and SIRT6 acts as ADP-ribosyl-transferase [19]. SIRT3 and SIRT6 are important in glucose and lipid metabolism [20]. SIRT3 is associated with the generation of reactive oxygen species (ROS), and the predominant source of ROS is mitochondria. Excessive ROS levels may contribute to the development of type 2 diabetes [21]. A recent study has shown that SIRT6 could regulate the expression of NF-κB [22].
Hesperetin is an aglycone form of hesperidin, which is present in peels of several citrus fruits, and research on the health effects of hesperetin is progressing actively [23]. Hesperetin has been studied for its potential hypolipidemic [24] and anti-cancer properties [25]. Moreover, hesperetin exerts effects on diabetes-induced retinal oxidative stress, neuroinflammation, and apoptosis in rats [26]. However, the protective effects and mechanism of hesperetin action on the diabetic environment have not been fully described.
Thus, in this study, we investigated the effects and mechanism of action of hesperetin on diabetic inflammation in THP-1 cells. By using experimental conditions that mimic diabetic complications, we investigated whether hesperetin treatment can modulate inflammation through the TLR/MyD88/NF-κB signaling pathways and SIRT expression.
Go to : 
MATERIALS AND METHODS
Reagents
Hesperetin, LPS, 3-(4, 5-dimethyl thiazol-2-yl)-2, 5-diphenyltetrazoliumbromide (MTT), and PMA were obtained from Sigma Aldrich (St. Louis, MO, USA). Dimethyl sulfoxide (DMSO) was obtained from Biosesang (Seongnam, Korea). Quantitative polymerase chain reaction (qPCR) primers were procured from Bioneer (Daejeon, Korea). The BCA protein assay kit was procured from Thermo Fisher Scientific (Waltham, MA, USA).
Cell culture and sample treatment
The human monocyte cell line THP1 was obtained from Korean Cell Line Bank (Seoul, Korea). THP1 cells were cultured in RPMI 1640 medium (Welgene, Daegu, Korea) supplemented at 37°C in 5% CO2. For monocyte to macrophage differentiation, THP1 cells (4 × 106 cells/mL) were seeded in a cell culture dish in RPMI 1640 medium with 1 μM PMA for 48 h. Differentiated THP1 cells were treated with hesperetin for 48 h in the absence or presence of LPS for 6 h under normoglycemic (NG, 5.5 mM/L glucose) or hyperglycemic (HG, 25 mM/L glucose) conditions. Next, medium was collected for measurement of cytokine release. Cells were washed in PBS and then harvested.
Cell viability assay
Cytotoxicity effects of hesperetin on cultured THP1 cells were measured by MTT assay. THP1 cells were seeded at 1 × 105 cells/well in a 24-well plate and treated with hesperetin for 48 h. The cells were then treated with LPS (100 ng/mL) for 6 h under NG or HG conditions. Then, MTT solution (100 μL; 1 mg/mL) was added, and the cells incubated for a further 3 h. After incubation, the supernatant was removed, and the precipitated formazan was solubilized in 150 μL of 100% DMSO. Absorbance was measured at 570 nm with an EZRead 400 Microplate reader (Biochrom, Cambridge, UK).
Western blot
Protein (20 μg) was mixed with buffer (100 mM Tris at pH 7.5, 2% sodium dodecyl sulfate, 2% glycerol, 1% 2-mercaptoethanol, and 0.01% bromophenol blue), incubated at 100°C for 5 min, and loaded on 10% polyacrylamide gels. Electrophoresis was performed using the Mini Protein Tetra Cell (Bio-Rad, CA, USA). Proteins were transferred to nitrocellulose membranes (Carlsbad, CA, USA), and the membranes blocked using blocking buffer (5% non-fat dried milk) for 2 h. The membranes were incubated with appropriate primary antibodies, then washed and incubated with the appropriate secondary antibody. Next, the membranes were visualized by applying the Western Blotting Luminol Reagent (Santa Cruz Biotechnology, Dallas, TX, USA) and performing autoradiography on a ChemiDoc XRS+ System (Bio-Rad). Equal loading of protein was normalized with β-actin and TATA-binding protein (TBP). The NFκB, COX-2, β-actin, and TBP were purchased from Santa Cruz Biotechnology. Acetyl-NFκB (Ac-NF-κB), IL-6, SIRT3, and SIRT6 were purchased from Cell Signaling Technology (Beverly, MA, USA). Signal quantification was performed using ImageJ software.
qPCR
Total RNA was isolated with TRIzol reagent (Ambion, Life Technologies, Carlsbad, CA, USA). The RNA concentration was measured using an absorbance ratio of 260 and 280 nm and a NanoDrop 2000 (Thermo Fisher Scientific). The cDNA was synthesized from total RNA using an Omniscript RT kit (QIAGEN, Hilden, Germany). The qPCR was performed with the CFX96 Touch Real-Time PCR system (Bio-Rad) and iQ SYBR Green Supermix (Bio-Rad) and the following primers (5′ to 3′, Forward [F], Reverse [R]); human TLR2 (F): GGGTCATCATCAGCCTCTCC, (R): GTGGATGTAATCGTTGTCACTGGA, human TLR4 (F): CAGAGTTGCTTTCAATGGCATC, (R): GGAGGTCCAAGAACTAATGTCAGA, human MyD88 (F): TGCTGGAGCTGGGACCCAGCATTGAGGAGGA, (R): TCAGACACACACAACTTCAGTCGATAG, human NFκB (F): GACAAGGTGCAGAAAGATGACAT, (R): TCATACGGTAACACAAGGCCT, human SIRT1 (F): CGGAAACAATACCTCCACCT, (R): CACCCCAGCTCCAGTTAGAA, human SIRT3 (F): ACATCGATGGGCTTGAGAGAGT, (R): CATGAGCTTCAACCAGCTTTGA, human SIRT6 (F): GTGCGCTCAGGCTTCCCCAG, (R): GGACAGGTCGGCGTTCCTGC, human β-actin (F): CACCCCGTGCTGCTGAC, (R): CCAGAGGCGTACAGGGATAG. Data were analyzed using the 2−ΔΔCT method and normalized using β-actin as a housekeeping gene.
Immunofluorescence staining
The cells were seeded at 1 × 105 cells/well in a 24-well plate and subsequently treated with hesperetin with LPS (100 ng/mL) treated prior to immunofluorescence staining. The cell medium was removed, and the cells washed twice with PBS. The PBS was removed, and the cells were fixed with 4% paraformaldehyde (PFA) for 30 min. Next, the 4% PFA was removed and the cells washed in PBS. Cells were incubated with NF-κB p65 and Ac-NF-κB p65 antibodies. For detection of nucleic acid, 4′,6-diamidino-2-phenylindole (DAPI) solution was incubated at 37°C. The mounting solution was dropped on a slide glass, and signal quantification was evaluated using a microscope (Leica Microsystems, Wetzlar, Germany).
Enzyme-linked immunosorbent assay (ELISA)
The IL-6 and TNF-α ELISA kits were purchased from Abcam (Cambridge, MA, USA) and used to evaluate the effect of hesperetin on cytokine levels in LPS-activated, PMA-differentiated THP-1 macrophages under NG or HG conditions. The cells were seeded at 1 × 105 cells/well in 24-well plates and treated with hesperetin for 48 h followed by LPS (100 ng/mL) for 6 h before harvesting. Cytokine concentrations in the supernatants of THP-1 macrophage cell cultures were determined by using the ELISA kits according to the manufacturer's instructions.
Statistical analysis
Results are reported as means ± SD. Each experiment was performed at least three times. Statistical significance was assessed by applying one-way analysis of variance (ANOVA; SPSS version 25.0 software) followed by Dunnett's post hoc test. Statistical significance is expressed as #
P < 0.05 and ##P < 0.01 compared with normoglycemic (NG) results or *P < 0.05 and **P < 0.01 compared with the LPS + hyperglycemic + hesperetin 0 μM (H0) results.
Go to :
RESULTS
Cytotoxic range of hesperetin in LPS-treated THP1 macrophages under high glucose conditions
The concentration of LPS and/or glucose required to create an environment that mimics diabetic conditions at the cellular level was determined. Inflammation was confirmed by measuring the COX-2 expression level. Fig. 1A shows the results of an experiment to determine LPS concentration in the absence of glucose treatment. The COX-2 expression level increased in cells with LPS 100 ng/mL and LPS 500 ng/mL compared to that in untreated LPS. However, treatment with LPS 500 ng/mL resulted in a slight decrease in COX-2 expression compared to that with LPS 100 ng/mL. Thus, we used a concentration of LPS 100 ng/mL in this experiment. Fig. 1B shows the results of an experiment to determine the COX-2 expressions at various concentration of glucoses with a 100 ng/mL LPS concentration. MTT assay was performed to study the effect of hesperetin on cell viability after 48 h incubation (Fig. 1C). THP1 cells (1 × 105 cells/mL) were cultured in the presence of LPS (100 ng/mL) under NG or HG conditions. Cells were exposed to different concentrations of hesperetin (0–100 μM) for 48 h, and the results showed that hesperetin treatment and LPS stimulation did not affect cell viability in THP1 macrophages.
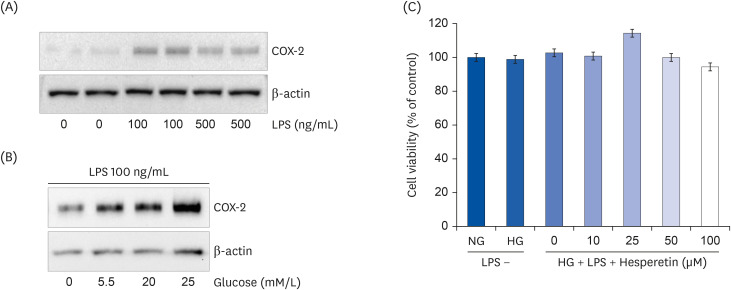 | Fig. 1Induction of COX-2 expression by LPS (A) or glucose (B) and the cytotoxic range of hesperetin (C) in LPS-treated THP-1 macrophages. COX-2 expression by glucose (0, 5.5, 20, and 25 mM/L) in LPS (100 ng/mL) was determined by western blotting. Effect of hesperetin on cell viability of THP-1 macrophages under LPS and hyperglycemic condition after 48 h was evaluated by performing 5-diphenyltetrazoliumbromide assays. The data are representative of at least three independent experiments.COX-2, cyclooxygenase-2; LPS, lipopolysaccharide; NG, normoglycemic 5.5 mM/L glucose; HG, hyperglycemic 25 mM/L glucose.
|
Hesperetin suppresses release of proinflammatory cytokines in LPS-treated THP1 macrophages under hyperglycemic conditions
To examine whether hesperetin could inhibit high glucose and LPS-induced inflammatory cytokine expressions in differentiated THP1 cells, the cells were treated with 10–50 μM hesperetin for 48 h, then stimulated using LPS 100 ng/mL for 6 h. Cell culture media were collected for measurement of cytokine release using ELISA (Fig. 2A and B) and western blotting (Fig. 2C). In the HG and LPS + HG conditions, TNF-α and IL-6 expressions were significantly increased compared to that under NG conditions and significantly decreased when the cells were treated with hesperetin.
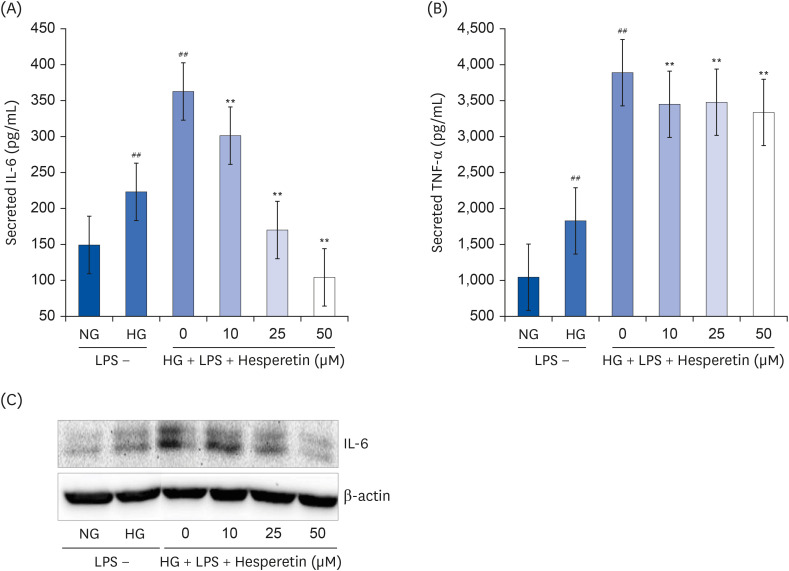 | Fig. 2Effect of hesperetin on cytokine release in LPS-treated THP-1 macrophages under hyperglycemic conditions. Cytokine levels in cell culture media were measured using enzyme-linked immunosorbent assay kits according to the manufacturer's instructions (A, B). Values were calculated based on the assay's standard curve. IL-6 levels were evaluated by western blot analysis (C). The data are representative of at least three independent experiments.IL-6, interleukin-6; NG, normoglycemic 5.5 mM/L glucose; HG, hyperglycemic 25 mM/L glucose; LPS, lipopolysaccharide; TNF-α, tumor necrosis factor-α; H0, LPS + hyperglycemic + hesperetin 0 μM.
##P < 0.01 compared with NG; **P < 0.01 compared with H0.
|
Hesperetin regulates TLR-mediated signaling pathway-related TLR2, TLR4, and MyD88 gene expressions in LPS-stimulated THP1 macrophages under hyperglycemic conditions
qPCR was performed to confirm the mRNA expressions of TLR2/4 and MyD88 in THP-1 macrophages exposed to high glucose and LPS-induced inflammatory conditions that mimic complex diabetic diseases. The inhibitory effect of hesperetin on the activation of TLR2/4 and MyD88 was also investigated. The mRNA level expressions of TLR2/4 and MyD88 increased under HG and LPS + HG conditions compared to that under NG conditions. However, the expressions of these molecules were significantly decreased following hesperetin treatment (Fig. 3). Modulating TLRs may be a useful strategy in preventing diabetic complications, given the crucial role of inflammation in microvascular and associated complications [27]. Our results suggested that hesperetin regulates inflammatory responses within the TLR2, TLR4, and MyD88 pathways.
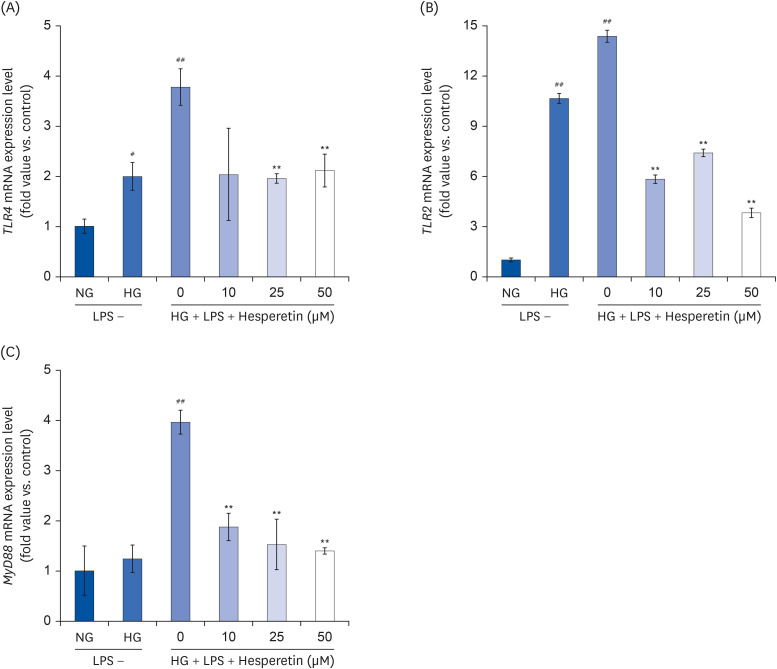 | Fig. 3Effect of hesperetin on TLR2, TLR4, and MyD88 gene expressions in LPS-treated THP-1 macrophage under hyperglycemic conditions. Cells were treated with hesperetin (0, 10, 25, and 50 μM) for 48 h, and mRNA expressions of TLR2, TLR4, and MyD88 were evaluated by quantitative polymerase chain reaction (A, B, C). The data are representative of at least three independent experiments. Data were analyzed using the 2−ΔΔCT method.NG, normoglycemic 5.5 mM/L glucose; HG, hyperglycemic 25 mM/L glucose; LPS, lipopolysaccharide; H0, LPS + hyperglycemic + hesperetin 0 μM.
#P < 0.05, ##P < 0.01 compared with NG; **P < 0.01 compared with H0.
|
Hesperetin reduces NFκB protein and mRNA expressions in LPS-treated THP1 macrophage under hyperglycemic conditions
Western blotting, qPCR, and immunofluorescence assay were performed to examine the expression of NFκB in THP1 cells stimulated with LPS and cultured under hyperglycemic conditions. The potential inhibitory effect of hesperetin on the activation of NFκB was also investigated. NFκB protein expression in the nucleus increased in the HG and LPS + HG conditions and decreased when cells were treated with hesperetin (Fig. 4A, B, and C). Consistent with protein expression results, the mRNA level of NFκB increased in HG and LPS + HG conditions compared to that under the NG condition but decreased significantly in response to treatment with hesperetin (Fig. 4D). Using immunofluorescence, the expression of NFκB and the active form of Ac-NF-κB were shown to be increased under HG and LPS + HG conditions and decreased in cells treated with 50 μM hesperetin (Fig. 4E and F). These results indicated that hesperetin treatment could suppress NFκB expression by blocking NFκB translocation to the nucleus.
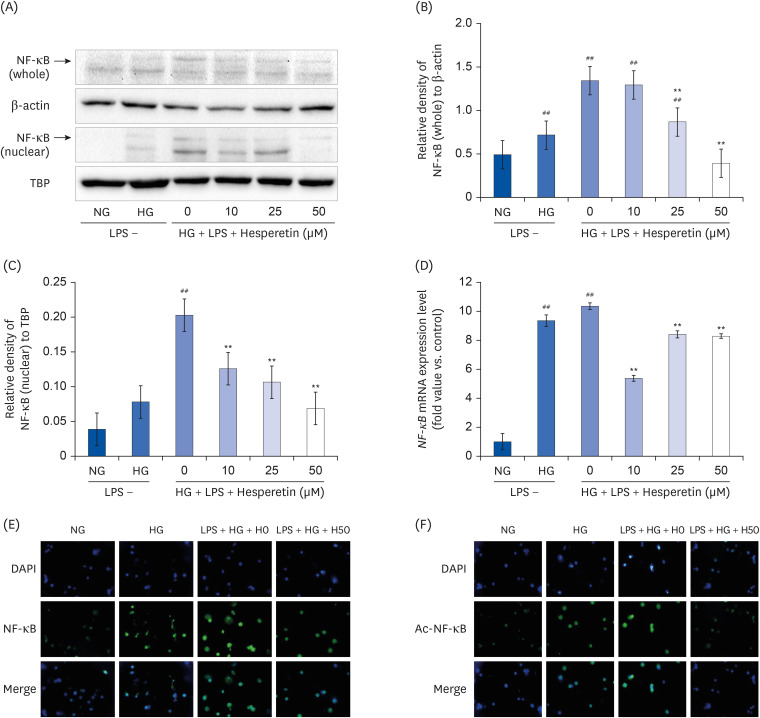 | Fig. 4Effect of hesperetin on NF-κB protein and mRNA expressions in LPS-treated THP-1 macrophages under hyperglycemic conditions. NF-κB levels were evaluated by western blot analysis of cell lysates (A). The percentage of NF-κB (whole)/β-actin (B) and the percentage of NF-κB (nuclear)/TBP (C) were revealed by image J. Cells (1 × 106 cells/mL) were treated with hesperetin for 48 h, and NF-κB mRNA levels were evaluated by qPCR (D). Immunofluorescence images show downregulation of NF-ĸB (E) and Ac-NF-ĸB (F) in LPS-treated THP-1 macrophages under hyperglycemic conditions following hesperetin treatment. Differentiated THP-1 cells were cultured in the presence of LPS (100 ng/mL) under normoglycemic or hyperglycemic conditions in the absence or presence of hesperetin (50 μM) for 48 h (400 × Magnification). Signal quantification was performed using ImageJ software. The data are representative of at least three independent experiments. Data were analyzed using the 2−ΔΔCT method.NF-κB, nuclear factor-κB; TBP, TATA-binding protein; NG, normoglycemic 5.5 mM/L glucose; HG, hyperglycemic 25 mM/L glucose; LPS, lipopolysaccharide; H0, LPS + hyperglycemic + hesperetin 0 μM; DAPI, 4′,6-diamidino-2-phenylindole.
##P < 0.01 compared with NG; **P < 0.01 compared with H0.
|
Hesperetin increases SIRT3 and SIRT6 expressions in LPS-treated THP1 macrophages under hyperglycemic conditions
We analyzed the effect of hesperetin treatment on the expression levels of SIRT3 and SIRT6 in THP-1 macrophages cultured under diabetes-mimicking conditions by performing western blot and qPCR. As shown in Fig. 5A, western blotting showed that SIRT3 protein expression increased under the NG condition but decreased under the HG and LPS + HG conditions. Conversely, SIRT3 protein expression increased following treatment with hesperetin. Consistent with this, SIRT3 mRNA levels increased significantly with hesperetin 25 and 50 μM treatment (Fig. 5B); similarly, SIRT6 mRNA levels increased (Fig. 5C), but there was no significance to the changes.
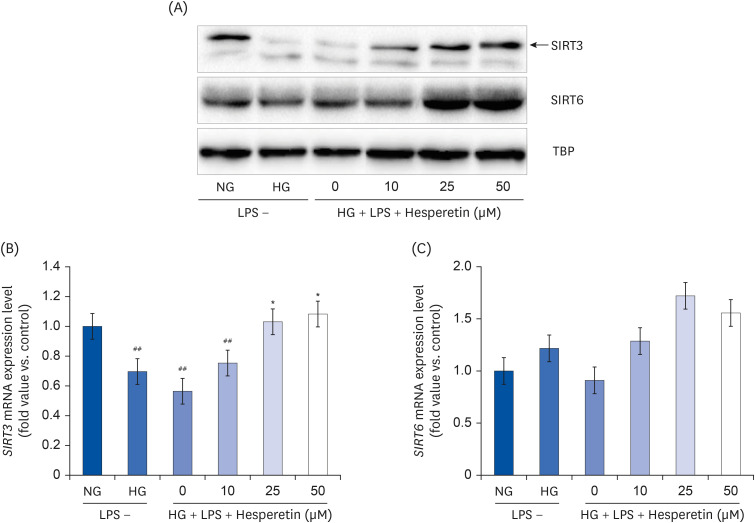 | Fig. 5Modulation of SIRT3 and SIRT6 protein expressions by hesperetin in LPS-treated THP-1 macrophages under hyperglycemic conditions. SIRT3 and SIRT6 protein levels were evaluated by western blot analysis and normalized to TBP (A). SIRT3 and SIRT6 mRNA levels were evaluated by quantitative polymerase chain reaction (B, C). The data are representative of at least three independent experiments. Data were analyzed using the 2−ΔΔCT method.NG, normoglycemic 5.5 mM/L glucose; HG, hyperglycemic 25 mM/L glucose; LPS, lipopolysaccharide; H0, LPS + hyperglycemic + hesperetin 0 μM; TBP, TATA-binding protein.
##P < 0.01 compared with NG; *P < 0.01 compared with H0.
|
Go to :
DISCUSSION
High blood glucose levels in type 2 diabetic patients are reported to increase inflammatory cytokines and ROS expressions via the polyol pathway, nicotinamide adenine dinucleotide phosphate oxidase, advanced glycation end products pathway, and the mitochondrial electron transport system [2829]. Increased Inflammatory cytokine and ROS levels contribute to malignant circulation, which contributes to an increased incidence of complications and reduced insulin secretion [303132].
Several natural components present in common foods can inhibit the expression of inflammatory cytokines. For instance, curcumin can regulate high glucose-induced proinflammatory cytokine production in monocytes [33]. The present study investigated hesperetin, a flavanone glycoside in citrus fruits. The study aimed to determine whether hesperetin could be a therapeutic agent against inflammation, particularly inflammatory responses that contribute to diabetes and diabetic complications. Increased secretion of inflammatory cytokines is known to reduce insulin sensitivity and induce diabetes, and abnormal expression levels are accompanied by systemic low-level inflammatory conditions, which are ultimately responsible for the onset of diabetic complications [3435].
Several studies have suggested that inflammatory markers, such as TNF-α and IL-6, are associated with diabetes and glucose disorders [3637]. According to Xie et al. [38] blueberries can reduce proinflammatory cytokine TNF-a and IL-6 production in mouse macrophages. Also, Guha et al. [39] reported that a high glucose level can upregulate the expression of TNF-α in THP-1 cells. In support of those studies, herein, we show that the expression of TNF-α and IL-6 is significantly decreased by hesperetin treatment compared to that under hyperglycemic and LPS + hyperglycemic conditions. Thus, hesperetin is considered to have anti-inflammatory properties effective against diabetes.
Increased TLR4 and NF-κB levels have been reported in patients with inflammatory chronic kidney disease, non-alcoholic fatty liver disease, and diabetes and can lead to increases in other proinflammatory cytokines [4041]. In addition, an increase in TLR2 in an LPS-induced inflammatory environment leads to enhanced NF-κB activation and COX-2 expression [42]. According to Ghanim et al. [43] in normal-weight men and women, when taken with a high-fat/high-carbohydrate meal, orange juice can prevent meal-induced oxidative and inflammatory stresses, including increased endotoxin levels and TLR expression. Previous studies have also reported that phytochemicals such as resveratrol [41] and curcumin [27] are effective against diabetes by inhibiting TLR signaling. Our results have shown that hesperetin treatment can significantly reduce TLR2, TLR4, and MyD88 mRNA levels in LPS-treated THP-1 macrophages under hyperglycemic conditions, suggesting that hesperetin inhibits TLR activation in LPS-induced inflammation.
The expression of inflammatory cytokines is necessarily accompanied by NF-κB activity. When NF-κB is activated by LPS and a high glucose level, it migrates from the cytoplasm to the nucleus and induces gene expression [44]. This results in the expression of proinflammatory cytokines that lead to diabetic complications. According to Yang et al. [45] epigallocatechin gallate inhibits inflammatory agents such as LPS and proinflammatory cytokines such as TNF-α. Another study showed that resveratrol inhibits the expressions of COX-2 and inducible nitric oxide synthase by inhibiting NF-κB activation induced by proinflammatory stimulants such as LPS and H2O2 [4647]. Our data showed that NF-κB expression and mRNA levels increased in THP-1 macrophages treated with LPS and/or maintained under hyperglycemic conditions and decreased following treatment with hesperetin. Furthermore, as a result of immunofluorescence staining, the expression of Ac-NF-κB was reduced by treatment with hesperetin. Thus, hesperetin inhibits the activation of NF-κB, which is mediated by several proinflammatory stimuli in diabetic complications.
The activity of SIRTs in metabolic syndrome is presumed to be related to aging, nutrition, and type 2 diabetes [48]. SIRT3 and SIRT6 are associated with dysregulated metabolism and other complications observed in diabetes. [48]. Elliott et al. [49] reported a decrease in fasting and postprandial blood glucose levels and decreased postprandial insulin levels in type 2 diabetes patients treated with resveratrol. Our data showed that SIRT3/6 protein and mRNA expressions in THP-1 macrophages maintained in a diabetes-mimicking environment increased following treatment with hesperetin. Therefore, hesperetin may produce effective therapeutic outcomes in diabetes by regulating SIRT 3/6 expression.
In the present study, we observed that treatment with hesperetin modulates inflammatory cytokine release, NF-κB acetylation, and SIRT3/6 expression via the TLR/MyD88/NF-κB signaling pathways. Thus, we propose that hesperetin may be a potential therapeutic agent for preventing diabetes and diabetic complications.
Go to :
 XML Download
XML Download