

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Obesity is a significant risk factor for many diseases, including type 2 diabetes, hypertension, and cardiovascular disease, and thus increases the economic burden. The current scenario of obesity illustrates that obesity incidence is increasing more rapidly in newly industrialized and developing countries than in Western countries, owing to changes in dietary patterns and sedentary lifestyle. A recent report predicts that, by 2030, the prevalence of obesity in South Korea will be 62% among men and 37% among women [1]. In developing countries like India, the prevalence of obesity in urban area is around 13%–50% of the population whereas in rural areas it is 8%–32% [2]. Since obesity increase both the risk of diseases and contribute to the economic burden, maintaining a healthy body weight is of significant benefit. However, synthetic drugs used as anti-obesity medications often have undesired side effects. Therefore, natural compounds with potential anti-obesity effects that are efficient, safe, and have long-term benefits are of important research interest [3].
White adipose tissue is the primary organ that undergoes extensive remodeling in response to changes in the nutritional status of the body. Adipose tissue, primarily consisting of adipocytes, is also the residing site for several other cells, including monocytes, macrophages, and pluripotent stem cells which regulate the functioning of this tissue [4]. Obesity is characterized by an increase in adipose tissue mass, which occurs due to the expansion of adipocytes (hypertrophy) and the formation of new adipocytes from preadipocytes increasing their number (hyperplasia). Hyperplasia occurs mostly during childhood and adolescence, whereas hypertrophy occurs when the adipocyte cells accumulate fat as triglycerides (TGs). The formation of adipocytes from pluripotent stem cells occurs through a complex sequence of events known as adipogenesis. Briefly, adipogenesis comprises two major phases: 1) determination, wherein the pluripotent stem cells commit to becoming adipocytes, and 2) terminal differentiation, wherein the preadipocyte cells obtain the characteristic features of mature adipocytes [56]. The molecular regulation of the terminal differentiation phase is mediated by transcription factors such as peroxisome proliferator-activated receptor gamma (PPARγ), CCAAT/enhancer binding proteins (C/EBPs), and sterol regulatory element-binding protein (SREBP). PPARγ and C/EBPα stimulate the expression of genes such as adipocyte protein-2 (aP2), lipoprotein lipase (LPL), and adipocyte markers like leptin and adiponectin, that are involved in lipogenesis, lipolysis, and insulin sensitivity. SREBP1c initiates the expression of enzymes such as acetyl-CoA carboxylase (ACC) and fatty acid synthase (FAS) [78]. Targeting the transcription factors and their downstream molecules and identifying possible anti-obesity agents is of pharmacological significance [9].
Abeliophyllum distichum, known in Korea as “Miseon-namu,” is a 1–2 m tall growing shrub that belongs to the Oleaceae family and flowers in the early spring season. It is categorized as an endangered species and is endemic to Korea. Recently, the health benefits of A. distichum have been investigated. A. distichum flower extract is reported to have anti-inflammatory effects against lipopolysaccharide (LPS)-induced inflammation in Raw 264.7 cell lines [10]. The callus from A. distichum reduced reactive oxygen species-mediated cellular damage in skin cells [11]. Park et al. [12] showed that an extract containing the flower, leaf, and branch exerts anti-cancer activity against human colorectal cancer cells. In addition, the leaves of A. distichum have been shown to suppress postmenopausal osteoporosis in rats [13]. Our team recently showed that the leaf extract of A. distichum reduce body weight and ameliorate obesity-related parameters in mice fed with a high-fat diet [14]. In the present study, we investigated the inhibitory effect of A. distichum leaf extract (ALE) on lipid accumulation in mature adipocytes and elucidated the mechanism of action, focusing specifically on adipocytes.
MATERIALS AND METHODS
Cells and reagents
3T3-L1 preadipocyte cells were purchased from the Korean Cell Line Bank (Seoul, Korea). Dulbecco's Modified Eagle's Medium (DMEM) and penicillin-streptomycin were purchased from HyClone (Logan, UT, USA). Fetal bovine serum (FBS), bovine serum (BS), and phosphate-buffered saline (PBS) were purchased from Gibco (New York, NY, USA). Trypsin-EDTA solution, isobutylmethylxanthine (IBMX), dexamethasone (DEXA), and insulin were purchased from Sigma-Aldrich (St. Louis, MO, USA) and EZ-Cytox solution was purchased from DoGEN Bio (Seoul, Korea). TG quantification kit was purchased from Biovision (Milpitas, CA, USA) and Cell Proliferation ELISA Bromodeoxyuridine (BrdU) colorimetric kit from Roche (Mannheim, Germany). Trizol reagent was purchased from Invitrogen (Carlsbad, CA, USA). PrimeScript TM RT master mix for cDNA synthesis and SYBR green PCR master mix were purchased from Takara (Shiga, Japan) and Toyobo (Osaka, Japan), respectively.
Preparation of ALE and high-performance liquid chromatography (HPLC) analysis
A. distichum leaves were obtained from Woorinamoo Agricultural Union Corporation (Goesan, Korea). The collected leaves were then processed and extracted at Korea Prime Pharmacy Co., Ltd. (Gwangju, Korea). Two kilograms of the leaves were dried, crushed and extracted with 40 L of 70% ethanol, for 4 h at 75°C. After filtration using Whatman filter paper (No. 4, Whatman International Ltd., Kent, United Kingdom), the solvent was evaporated and lyophilized to obtain 1.02 kg of ALE extract which was stored at −20°C until use. The samples were analyzed by HPLC on a Shimadzu liquid chromatography system LC-20AD using a diode array detector (SPD-M20A) and YMC-Pack ODS-AM column (4.6 × 250 mm, 2.6 μm). Solvents A (0.1% formic acid in water) and solvent B (0.1% formic acid in acetonitrile) were used as mobile phase. The run time was 30 min with a flow rate of 1 mL/min, detected at 330 nm and the concentration was calculated comparing the sample peak area to that of standards. The results obtained is given in Supplementary Fig. 1.
Cytotoxicity assay
Cells were cultured in DMEM containing 10% BS and 100 U/mL penicillin-streptomycin at 37°C in a 5% CO2 incubator. The cytotoxic effect of the sample was assessed using the following method. After overnight adherence of the cells seeded in 96-well plates, they were treated with different concentrations of ALE for 24, 48, and 72 h. Culture medium containing EZ-Cytox solution was added to the cells at the aforementioned time points; the cells were then incubated for 2 h and the absorbance at 450 nm was measured using a microplate reader (SpectraMax M2, Molecular devices, San Jose, CA, USA).
Differentiation and treatment
Cells were differentiated by replacing BS media with induction media consisting 10% FBS, 0.5 mM IBMX, 1 μM DEXA and 10 μg/mL insulin in DMEM, two days following 100% cell confluence. The induction media was replaced after 48 h with complete media containing DMEM with 10% FBS and 10 μg/mL insulin. The cells were maintained in complete media for six days, replacing the media every other day. The cells were treated with selected concentrations of ALE along with the respective media, based on the experimental design. To analyze the effect of ALE in each stage of differentiation, the cells were treated with ALE at three different time periods: from the 1st to 8th day (full period, Fp), only on days 1 and 2 of the differentiation period (early period, Ep) and only on days 7 and 8 of the differentiation period (terminal period, Tp).
TG assay and Oil Red O staining
Following the removal of media, cells were washed twice with PBS. For TG quantification, the cells were homogenized with 5% Triton X and assayed using a commercially available kit. For lipid staining, the cells were washed with PBS, fixed with 10% formalin, stained with Oil Red O solution for 1 h and washed with distilled water. After drying, the pictures were taken at 100× magnification. For quantification, the stained cells were washed with isopropyl alcohol, and the absorbance of the extracted isopropanol was measured at 520 nm.
BrdU incorporation assay
Cells were seeded in 96-well plates and treated with induction media with or without ALE for 12 h and 24 h. Cell proliferation was measured by calculating the incorporation of BrdU into DNA using Cell Proliferation ELISA BrdU colorimetric kit, following the manufacture's protocol.
Quantitative Reverse Transcription polymerase chain reaction (PCR)
Total RNA from cells was isolated using Trizol reagent and checked for purity (based on the 260:280 absorbance ratio) on a BioDrop Duo instrument (Biochrom, Holliston, MA, USA). Further, the cDNA synthesized using PrimeScriptTM RT master mix was used to perform PCR with SYBR green PCR master mix. Primer sequences used are given as Table 1.
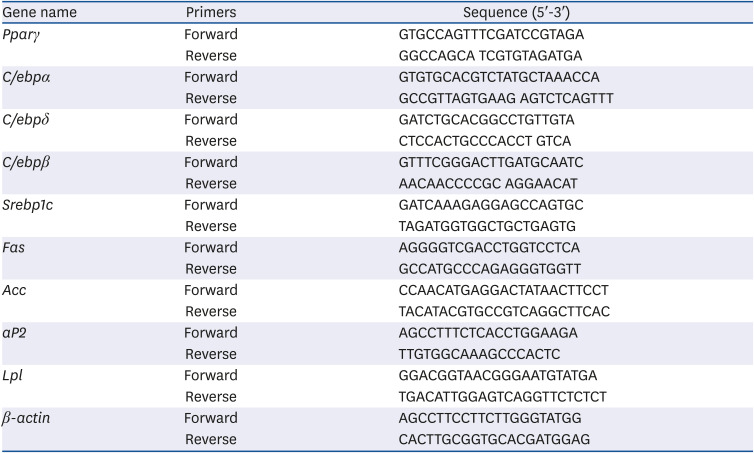
Table 1
List of primers used in for polymerase chain reaction
PPAR, peroxisome proliferator-activated receptor; C/EBP, CCAAT/enhancer binding protein; SREBP, sterol regulatory element-binding protein; FAS, fatty acid synthase; ACC, acetyl CoA carboxylase; aP2, adipocyte protein-2; LPL, lipoprotein lipase.
![]()
Western blot
The protein expression of PPARγ and β-actin (Cell Signaling Technology, Beverly, MA, USA) in cells treated for a full period was measured using western blot. The effect of ALE on MAPK signaling pathways was determined by checking the expression levels of proteins such as extracellular signal-regulated kinases (ERK), c-Jun amino-terminal kinase (JNK), and p38 for their total and phosphorylated forms. The method of protein extraction was as follows: after the lysis of cells using radioimmunoprecipitation assay (RIPA) buffer, it was centrifuged (4°C, 12,000 × g, 15 min), and the supernatant was collected. After protein quantification, the samples were matched to the same concentration, and heated at 95°C after mixing with a 5× protein buffer. The proteins were electrophoresed on a 10% sodium dodecyl sulfate-polyacrylamide gel and then transferred onto a polyvinylidene difluoride membrane (Bio-Rad Laboratories, Hercules, CA, USA) for blotting.
RESULTS
Effect of ALE on cell viability
As shown in Fig. 1, the cytotoxic assay revealed that ALE reduced the cell viability at high concentrations (400 and 500 µg/mL). At 24 h, the cell viability was lower than that of the control group by 18% and 26% for the 400 and 500 µg/mL concentrations, respectively. A reduction of 53% and 42% in cell viability was noticed with 400 µg/mL of ALE at 48 and 72 h, respectively. 500 µg/mL of ALE reduced cell viability by 68% and 66% at 48 and 72 h, respectively. However, up to 100 µg/mL ALE did not show any significant reduction in cell viability, whereas 200 µg/mL ALE showed a reduction in cell viability by 2%, 22%, and 6% at 24, 48, and 72 h, respectively. Considering the period of treatment and percentage of cell viability, concentrations up to 200 µg/mL were selected for further experiments.
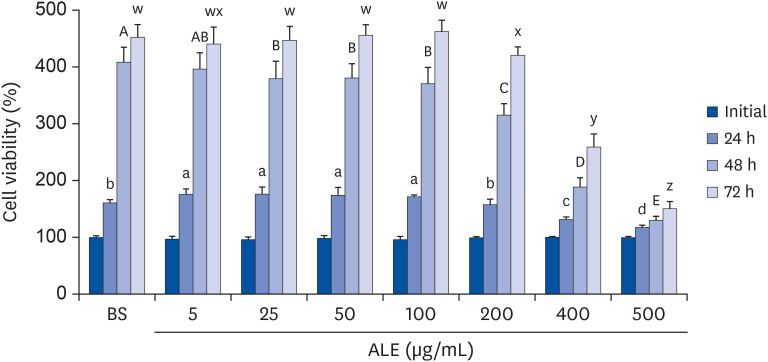
Fig. 1
Effect of ALE on cell viability. 3T3-L1 preadipocytes were treated with different concentrations of ALE for 24, 48, and 72 h, and cell viability was analyzed using MTT assay. Data expressed as mean ± SD (n = 8). Values with different superscripts; (a-d) for 24 h, (A-E) for 48 h, (w-z) for 72 h, indicate significant difference among groups by analysis of variance with Duncan's multiple range test at P < 0.05.
ALE, Abeliophyllum distichum leaf extract; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.
![]()
Effect of ALE on lipid accumulation in differentiated adipocytes
To verify the effect of ALE on lipid accumulation in differentiated adipocytes, we performed TG assay and Oil Red O staining after a full period of treatment. Oil Red O staining clearly revealed that 100 and 200 µg/mL concentrations of ALE reduced lipid accumulation in the differentiated cells (Fig. 2A and B). The lipid accumulation in ALE-treated cells were significantly decreased compared to that in the control group, in a dose-dependent manner. As shown in Fig. 2C, 50, 100, and 200 µg/mL concentrations of ALE decreased TG accumulation by 11%, 35% and 80%, respectively.
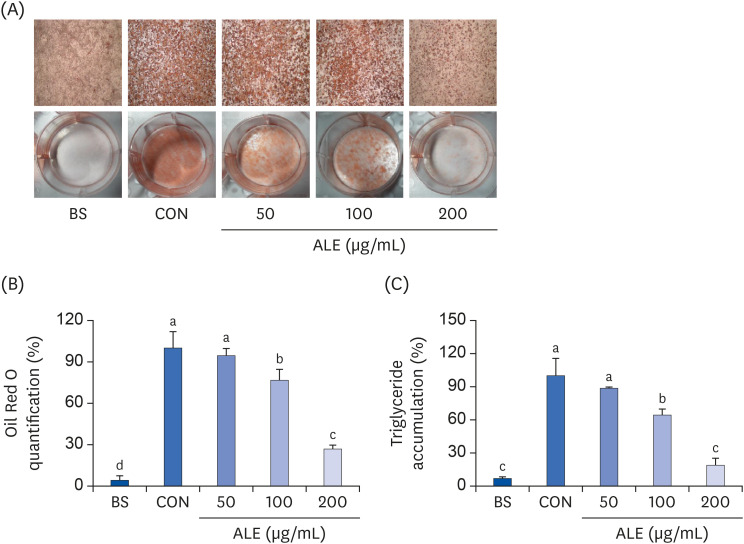
Fig. 2
Effect of ALE on adipocyte differentiation. 3T3-L1 preadipocytes were induced to differentiate with induction media in the presence or absence of ALE for 8 days and were analyzed for lipid accumulation. (A) Oil red O staining, (B) Oil Red O quantification, and (C) triglyceride quantification. Data expressed as mean ± SD (n = 6). Values with different superscripts indicate significant differences among groups by analysis of variance with Duncan's multiple range test at P < 0.05.
ALE, Abeliophyllum distichum leaf extract; BS, non-differentiated cells; CON, differentiated adipocytes.
![]()
Effect of ALE on lipid-metabolism related genes in differentiated adipocytes
Since 100 and 200 µg/mL concentrations of ALE significantly reduced lipid accumulation in differentiated adipocytes, we measured the mRNA expression of lipid metabolism-related genes in cells treated with these two concentrations. As shown in Fig. 3A, treatment with ALE significantly reduced the expression levels of Pparγ, Fas, and Acc irrespective of the doses. C/ebpα was reduced in a dose-dependent manner. ALE showed a tendency to decrease Srebp-1c gene expression. ALE treatment also significantly decreased the expression levels of Lpl and aP2 compared to that of the control group. In addition, ALE reduced the protein expression of PPARγ (Fig. 3B).
Fig. 3
Effect of ALE on adipocyte differentiation markers. 3T3-L1 preadipocytes were induced to differentiate with induction media in the presence or absence of ALE for 8 days and were analyzed for lipid accumulation. (A) mRNA expression levels of adipogenic and lipogenic genes and (B) Protein expression levels of PPARγ. Data expressed as mean ± SD (n = 6). Values with different superscripts indicate significant differences among groups by analysis of variance with Duncan's multiple range test at P < 0.05.
ALE, Abeliophyllum distichum leaf extract; BS, non-differentiated cells; CON, differentiated adipocytes; Ppar, peroxisome proliferator-activated receptor; C/ebp, CCAAT/enhancer binding protein; Srebp, sterol regulatory element-binding protein; Fas, fatty acid synthase; Acc, acetyl CoA carboxylase; aP2, adipocyte protein-2; Lpl- lipoprotein lipase.
![]()
Effect of ALE on different stages of adipocyte differentiation
To observe the inhibitory effect exerted by ALE on different stages of adipocyte differentiation, we examined lipid accumulation using Oil Red O staining and the expression levels of adipogenesis markers after treatment with 100 µg/mL (Supplementary Fig. 2) or 200 µg/mL (Fig. 4) of ALE during the early, terminal and full periods of the 8-day differentiation period. It was clearly evident from Oil Red O staining that treatment with ALE at an early period reduced lipid accumulation significantly compared to that of the terminal period (Fig. 4A). The cells treated with ALE for the full period showed approximately 43% reduction in lipid accumulation compared to that in the control cells (Fig. 4B). Interestingly, the cells treated only during the early period showed a 29% reduction in lipid accumulation. In contrast, lipid accumulation in cells treated only in the terminal period decreased by 3% compared to that in the control cells, indicating that the inhibitory effect of ALE on adipocyte differentiation is initiated in the early phase of differentiation. As shown in Fig. 4C, the mRNA expression of Pparγ, C/ebpα, and Fas was downregulated only in the early period and full period treatments but not in the terminal period treatment. This suggests that the inhibitory effect of ALE on adipocyte differentiation occurs mainly during the early adipogenic phase.
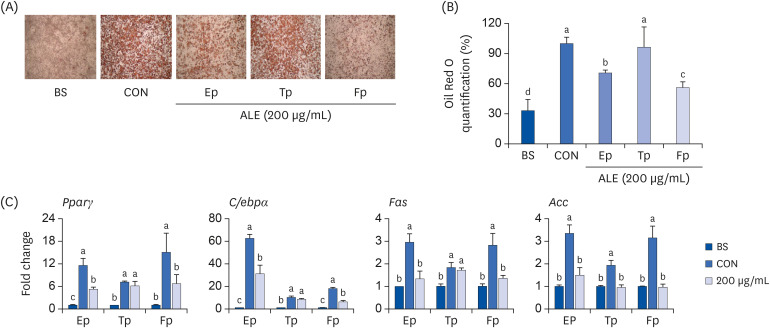
Fig. 4
Effect of ALE on adipocyte differentiation in cells treated at different time points. 3T3-L1 preadipocytes were induced to differentiate with induction media in the presence or absence of ALE for 8 days at different time points and were analyzed for lipid accumulation. (A) Oil red O staining, (B) Oil Red O quantification, and (C) Expression levels of genes associated with adipogenesis. Data expressed as mean ± SD (n = 6). Values with different superscripts indicate significant differences among groups by analysis of variance with Duncan's multiple range test at P < 0.05.
ALE, Abeliophyllum distichum leaf extract; BS, non-differentiated cells; CON, differentiated adipocytes; Ep, early period; Tp, terminal period; Fp, full period; Ppar, peroxisome proliferator-activated receptor; C/ebp, CCAAT/enhancer binding protein; Fas, fatty acid synthase; Acc, acetyl CoA carboxylase.
![]()
Effect of ALE on mitotic clonal expansion (MCE)
Considering the effect of ALE on lipid accumulation in the early phase, the mRNA expression levels of the transcription factors that regulate the initial adipogenic stages, C/ebpβ and C/ebpδ, were measured after treating the cells with ALE for 24 and 48 h. As shown in Fig. 5A, only 200 µg/mL of ALE significantly downregulated the expression of C/ebpβ at 24 h of treatment. In contrast, the C/ebpδ levels decreased after 48 h of treatment by only 200 µg/mL concentration of ALE, though 100 µg/mL showed a tendency to reduce C/ebpδ after 48 h of treatment. The BrdU assay is used to analyze the cells in the Synthesis phase of the mitosis [15]. Treatment with ALE at 200 µg/mL only significantly reduced the number of cells entering MCE, indicated by the lower levels of BrdU labeling, in the ALE-treated group compared to that of the control cells (Fig. 5B). At 12 h, only 200 µg/mL concentrations of ALE significantly decreased BrdU labeling, whereas at 24 h, this effect was dose-dependent.
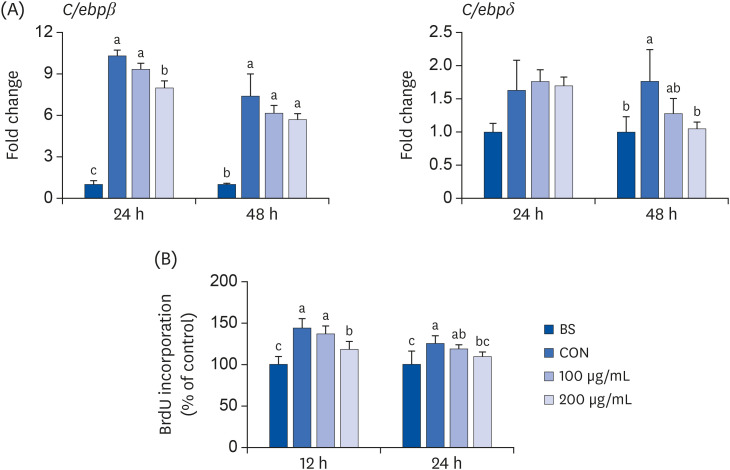
Fig. 5
Effect of ALE on the MCE. 3T3-L1 preadipocytes were induced to differentiate with induction media in the presence or absence of ALE for 24 and 48 h. (A) Expression levels of genes associated with MCE and (B) BrdU incorporation. Data expressed as mean ± SE (n = 6). Values with different superscripts indicate significant differences among groups by analysis of variance with Duncan's multiple range test at P < 0.05.
ALE, Abeliophyllum distichum leaf extract; BS, non-differentiated cells; CON, differentiated adipocytes; MCE, mitotic clonal expansion; BrdU, Bromodeoxyuridine; C/EBP, CCAAT/enhancer binding protein.
![]()
Effect of ALE on mitogen-activated protein kinase (MAPK) signaling pathway associated with adipogenesis
MAPK signaling pathway, consisting of three groups of molecules-ERK 1/2, JNK, and p38, modulates the expression of adipogenic transcription factors [16]. The phosphorylation of ERK 1/2, JNK, and p38 were significantly increased in the differentiated cells compared to the non-differentiated cells (Fig. 6). Treatment with ALE reduced the phosphorylation levels of ERK 1/2 and p38 in a dose-dependent manner. Also, ALE suppressed the phosphorylation of JNK, irrespective of the doses. Taken together, the results suggested that ALE reduced adipogenesis by suppressing the activation of MAPK signaling pathways, thereby inhibiting the activation of its downstream targets and the early phase of adipogenesis.
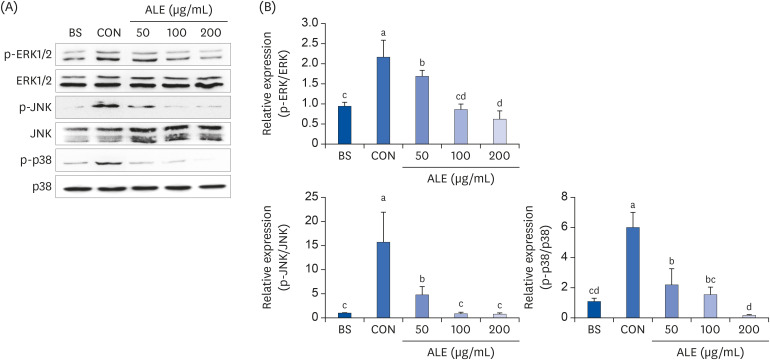
Fig. 6
Effect of ALE on MAPK signaling pathways. 3T3-L1 preadipocytes were induced to differentiate with induction media in the presence or absence of ALE for 2 h. (A) Western blot and (B) Quantification of protein levels. Data expressed as mean ± SD (n = 6). Values with different superscripts indicate significant differences among groups by analysis of variance with Duncan's multiple range test at P < 0.05.
ALE, Abeliophyllum distichum leaf extract; BS, non-differentiated cells; CON, differentiated adipocytes; MAPK, mitogen-activated protein kinase; ERK, extracellular signal-regulated kinases; JNK, c-Jun amino-terminal kinase.
![]()
DISCUSSION
Until a few decades ago, adipose tissue was considered only as a tissue mass that stores energy and provides insulation and protection to internal structures. However, the regulatory role of adipose tissue in maintaining body homeostasis became an important focus after the discovery of leptin in 1994 [17]. Adipocytes help maintain body homeostasis by secreting proteins that regulate blood pressure, immune function, angiogenesis, and energy balance. The increasing rate of obesity has made adipose tissue a significant point of interest, as it responds rapidly to the energy requirements of the body [5]. Obesity is characterized by an increase in adipose tissue mass due to adipogenesis and the regulation of the adipogenic process is a significant target for the control and/or prevention of obesity [15]. Anti-obesity agents of plant origin are gaining attention as they have fewer or no side effects than synthetic drugs [3]. A. distichum is a flowering shrub, endemic to Korea. It contains several bioactive compounds such as rutin, caffeic acid, ferulic acid and quercetin [18]. The HPLC analysis of the ALE used in the present study showed the presence of verbascoside as a major compound, followed by rutin (Supplementary Table 1). Verbascoside, also known as acteoside, is reported to improve lipid metabolism in HepG2 cells with oleic acid-induced lipid accumulation [19]. In the present study, we showed that ALE containing a high concentration of verbascoside inhibits adipocyte differentiation and prevents lipid accumulation in 3T3-L1 cells.
A few previous studies have shown that A. distichum leaves exhibit beneficial properties, including osteoprotective and anti-cancer effects [1318]. Our team has recently shown that the leaf extract of A. distichum has anti-obesity effects [14]. In the present study, we aimed to investigate the effect of ALE on the differentiation of adipocytes and elucidate the mechanism of action through which ALE may suppress lipid accumulation in adipocytes.
Upon stimulation by inducers such as insulin preadipocytes undergo changes to accommodate lipid transport and synthesis, become sensitive to insulin signaling, and secrete adipokines [5]. In the present study, treatment with ALE for a full period of 3T3-L1 preadipocyte differentiation reduced lipid accumulation in the mature adipocytes as evident from the TG assay and Oil Red O imaging. Furthermore, ALE treatment decreased the mRNA expression levels of Pparγ, C/ebpα, Lpl, aP2, Acc, and Fas compared to that of the control cells. PPARγ is known as the master regulator of adipogenesis, as it is inevitable and adequate for this process. In the absence of PPARγ, adipogenesis fails to occur and adipocytes cannot be maintained in the differentiated state [5]. PPARγ transcriptionally activates the expression of lipogenic genes such as aP2, FAS and ACC [9]. C/EBPα is involved in the induction of insulin receptor and insulin receptor substrate 1 (IRS-1), which is crucial for insulin sensitivity. C/EBPα and PPARγ are known to increase their expression levels via a positive feedback loop that contributes to the differentiation process [20]. The results of the present study suggest that ALE inhibits adipogenesis by lowering the major transcription factors and their downstream targets genes involved in lipogenesis.
To understand the mechanism of action of ALE, differentiating cells were treated at different time periods. Based on Oil Red O staining and gene expression analysis, it was evident that treatment with ALE in the early period (days 1 and 2) had a more significant inhibitory effect compared to treatment in the terminal period (days 7 and 8). This indicated that ALE might inhibit the early phase of adipogenesis. Growth-arrested preadipocytes, when differentiated using media containing FBS, insulin, DEXA, and IBMX, reenter the cell cycle and undergo two rounds of division, termed as MCE. In the first 24 h after induction, MCE is followed by the expression of a series of transcription factors responsible for early adipogenic events. Transcription factors such as PPARγ and C/EBPs (β, δ, and α) are responsible for the activation of genes responsible for the synthesis and accumulation of fat in mature adipocytes [6]. From the results of the present study, it was evident that the cells treated with ALE in the early period alone showed significantly lower levels of lipid compared to those treated in the terminal period alone, suggesting that ALE probably exerts its inhibitory effect mainly in the early adipogenic phase. Furthermore, ALE reduced BrdU incorporation in differentiating cells in the first 24 h of induction. ALE also decreased the mRNA expression levels of early adipogenic markers such as C/EBPβ and C/EBPδ. C/EBPβ is expressed immediately after the induction of differentiation and is reported to be a prerequisite for MCE. It is also involved in the activation of the transcription factors PPARγ and C/EBPα, which initiate the activation of genes involved in lipogenesis, and thus induce the adipocyte phenotype [21]. C/EBPδ is expressed in cells that are ready for differentiation, and increases the expression of a protein, factor of adipocyte differentiation 49 (fad49), and C/EBPβ, which are involved in MCE [22]. The fad49 expression is limited to differentiating cells and is transiently expressed. The expression of fad49 increases quickly after the differentiation is induced, reaches the peak at 3 h, and then decreases. Hishida et al. [23] have shown that knockdown of fad49 gene negatively affects the early stage of adipocyte differentiation by altering the levels of SREBP-1c, C/EBPβ and C/EBPδ. In the current study, the mRNA expression of C/EBPβ and C/EBPδ in ALE-treated cells decreased at 24 and 48 h, respectively. Altogether, these data indicate that ALE inhibits adipocyte differentiation by decreasing the number of cells undergoing MCE, thus lowering the expression of early adipogenic markers. In growth-arrested fibroblast-like cells, such as 3T3-L1 cells MAPK signaling molecules are known to be upregulated during differentiation [24]. ERK 1/2 is known to modulate cell proliferation and needed for the activation of early adipogenesis factors [16]. JNK activation in the adipocyte is supposed to be associated with adipocyte insulin resistance [25]. Activation of p38 stimulates the induction of PPARγ [26]. Treatment with ALE reduced the expression of phosphorylated MAPKs; ERK, JNK, and p38 indicating that the inhibitory effect of ALE on adipocyte differentiation could be attributed to the suppression of MAPK activation.
A. distichum is used in Korean traditional medicine to relieve pain [10]. Different plant parts including, leaf, flower, and branch, have recently been explored for possible health effects, including anti-inflammatory, anti-cancer, and osteoprotective effects [121327]. In the present study, we showed that the leaf extract of A. distichum significantly reduces lipid accumulation in mature adipocytes. This inhibitory effect was mediated through the suppression of the MAPK pathway resulting in downregulation of the MCE process and decreased expression of adipogenic transcription factors, resulting in the reduced the expression of lipogenesis markers. Additionally, we reported that ALE exerts anti-obesity effects evident from reduced body weight gain, decreased abdominal fat volume, improved serum lipid profile, and hepatic injury markers in high-fat diet-induced obesity mice model. Also, ALE prevented ectopic lipid accumulation and reduced lipogenesis in adipose tissue via activation of the AMPK pathway [14]. In the current study, we have shown that ALE exhibits anti-adipogenic and lipid-lowering effects on differentiating adipocytes by suppressing the MAPK signaling pathway. Taken together, our in vivo and in vitro results provide clear scientific evidence for the action of A. distichum in improving the adipose tissue metabolism and support the potential of A. distichum as a nutritional supplement with anti-obesity properties. Further studies may focus on the effect of the ALE on humans.
 XML Download
XML Download