

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Osteoporosis is a systematic skeletal disorder characterized by a decrease in bone density and microstructural degradation. It is a multifactorial disease caused by aging, lack of sex hormones, drug use, alcoholism, nicotine, and medical conditions (such as metabolic syndrome) in which progressive bone loss occurs [1]. This loss increases bone fragility and fracture risk, and serious complications can occur. These symptoms are highly likely to degrade the quality of life. Drugs used to prevent and treat osteoporosis include selective estrogen receptor modulators, parathyroid hormone analogs, bisphosphonates, calcitonin, and hormone replacement therapy drugs [2]. Due to the nature of osteoporosis, long-term use of these drugs is essential. However, long-term administration of these drugs increases the risk of side effects such as some cancers (breast cancer, epithelial carcinoma, etc.), heart attack, blood clots, and stroke [34]. Therefore, there is an increasing interest in natural products with potential bone-protecting effects but without side effects, even after long-term administration.
Petasites japonicus Maxim (P. japonicus), also called butterbur, giant butterbur, great butterbur, or sweet-coltsfoot, is an herbaceous perennial plant in the Asteraceae family. It is mainly distributed in Korea, China, Japan, and Sakhalin and is widely used in soups or seasoned vegetable dishes [56]. In East Asia, it has traditionally been used as an anti-pyretic, antitussive, or wound healing agent. It is a species introduced in Europe and North America by Japanese immigrants, but its utilization rate is low. In Europe and America, P. japonicus has been mainly used as a botanical dietary supplement, used to treat migraines, allergy, bronchial asthma, and ulcers [78].
Recently, studies into various physiological actions of P. japonicus have been conducted, and its anti-tumor [910111213], anti-allergic [1415], anti-inflammatory [1617], neuroprotective [18], anti-asthma [19], and anti-obesity [818] effects have been reported. However, the relationship between P. japonicus and bone metabolism has not been previously described. In this study, to investigate the anti-osteoporotic effect of P. japonicus extracts, we measured their effect on osteoblast-specific markers and transcription factors in MC3T3-E1 murine osteoblastic cells.
MATERIALS AND METHODS
Materials
The materials used in this study were purchased from the indicated suppliers: α-minimal essential medium (α-MEM), fetal bovine serum (FBS), and penicillin/streptomycin from Thermo Fisher Scientific Inc. (Waltham, MA, USA); 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), ascorbic acid, β-glycerophosphate and 17-estradiol (E2) from Sigma-Aldrich Co. (St. Louis, MO, USA); TRACP & ALP assay and osteocalcin enzyme-linked immunosorbent assay (ELISA) kits from Takara Bio Inc. (Kusatsu, Japan); Sirius Red collagen detection kit from Chondrex Inc. (Redmond, WA, USA); Osteogenesis assay kit from Millipore (Burlington, MA, USA); Trizol from Invitrogen Life Technologies (Carlsbad, CA, USA); HyperScript™ RT master mix kit from GeneAll Biotechnology (Seoul, Korea); Rotor-Gene™ SYBR Green kit from Qiagen (Valencia, CA, USA).
Preparation of aqueous and ethanolic extracts of P. japonicus
Wild P. japonicus was collected in July 2020 from Chuncheon, Korea. The leaves and stems of P. japonicus were separated, washed, air-dried at 45°C for 24 h, and pulverized to form a powder. To prepare the aqueous extract of P. japonicus, 50 g of each powder were refluxed in 1 L of water at 100°C for 3 h. Each resulting supernatant was filtered through Whatman filter paper #2, after which the filtrates were evaporated in a rotary vacuum evaporator and subsequently freeze-dried. The resulting powders were used as the aqueous extract of P. japonicus leaves (AL) and the aqueous extract of P. japonicus stems (AS). To prepare the ethanolic extracts of P. japonicus, 50 g of each powder were refluxed in 1 L of 95% ethanol at 70°C for 3 h. The resulting supernatants were filtered, evaporated, and freeze-dried as described above. The resulting powders were used as the ethanolic extract of P. japonicus leaves (EL) and the ethanolic extract of P. japonicus stems (ES). The yields of AL, AS, EL, and ES were 19.1 g, 41.6 g, 20.9 g, and 23.9 g per 100 g of each dried powder, respectively.
Determination of total phenolic content
Each extract's total phenolic content was determined by applying the Folin-Cioalteau colorimetric method [20] with minor modifications and using gallic acid as the standard phenolic compound. In brief, 40 µL of the extracts (1 mg/mL) were mixed with 0.1 mL of Folin-Cioalteau reagent (Sigma-Aldrich) and 1.56 mL of distilled water. After 5 min, 0.3 mL of 2N sodium carbonate was added and mixed thoroughly. The reactive mixture was allowed to stand at room temperature for 1 h and the absorbance of the resulting reactive mixture was measured at 750 nm. Quantitative measurements were based on a standard calibration curve of gallic acid. The total phenolic content was expressed as milligrams of gallic acid equivalents per gram (mg [GAE]/g).
Determination of total flavonoid content
Each extract's total flavonoid content was determined by employing the aluminum chloride colorimetric method [21] with slight modifications and using quercetin as the standard flavonoid. Briefly, 30 µL of the extracts (1 mg/mL) were mixed with 30 µL of 10% aluminum chloride, 30 µL of 1 M potassium acetate, and 1.41 mL of 95% ethanol. The reactive mixture was kept at room temperature for 40 min and the absorbance of the resulting reactive mixture was measured at 415 nm. Quantitative measurements were based on a standard calibration curve of quercetin. The total flavonoid content was expressed as milligrams of quercetin equivalents per gram (mg [QUE]/g).
Cell culture and differentiation
MC3T3-E1 murine osteoblastic cells were acquired from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in α-MEM supplemented with 10% FBS, 100,00 U/L penicillin, and 100 mg/L streptomycin in an incubator at 37°C and a humidified atmosphere of 5% CO2. When cells were 90% confluent, they were further incubated in α-MEM supplemented with 10% FBS, 10 mmol/L β-glycerophosphate, and 50 mg/L ascorbic acid (osteoblast differentiation-inducing medium, ObDIM) to induce osteoblast differentiation. The medium was changed to fresh ObDIM every 3 days.
Cell viability assay
MC3T3-E1 cells were plated in 24-well plates at an initial density of 5 × 104 cells/well. After incubation for 24 h, cells were treated with various concentrations of each extract and incubated for 72 h. Cell viability was assessed by MTT assay as described previously [22].
Measurement of alkaline phosphatase (ALP) activity
To measure ALP activity levels, MC3T3-E1 cells were plated in 48-well plates at a density of 2 × 104 cells/well and incubated for 24 h. After incubation for 24 h, cells were incubated in ObDIM containing each extract at various concentrations. Following 5 days of treatment, ALP activity was measured using the TRACP & ALP assay kit and following the manufacturer's instructions.
Assessment of collagen synthesis
Collagen synthesis in osteoblasts was assessed by staining cells using Sirius Red, an anionic dye that strongly binds to collagen molecules. MC3T3-E1 cells were plated and treated with AL for 8 days as described above. The degree of collagen synthesis was estimated using Sirius Red collagen detection kit in accordance with the manufacturer's instructions.
Assessment of mineralization
Mineralization of osteoblasts was assessed by staining cells with Alizarin Red S, an anthraquinonoid derivative that binds to calcium. MC3T3-E1 cells were plated, treated with AL as described above, and incubated for 14 days to induce mineral deposition. The degree of calcium deposition in the cells was estimated using an osteogenesis assay kit according to the manufacturer's instructions.
Measurement of osteoblast-secreted osteocalcin levels
MC3T3-E1 cells were plated and treated with AL as described above. Following 14 days of treatment, cell culture media conditioned for 24 h were collected. The levels of osteoblast-secreted osteocalcin in the 24 h-conditioned media were measured using an osteocalcin ELISA kit according to the manufacturer's instructions.
Quantitative real-time reverse transcription-polymerase chain reaction (RT-PCR)
By following the manufacturer's instructions, total RNA was extracted with Trizol. The purity and content of total RNA were determined using a micro-volume spectrophotometer (BioSpec-nano, Shimadzu, Kyoto, Japan). Complementary DNA was synthesized from 2 µg of total RNA using a HyperScript™ RT master mix kit. Real-time PCR of complementary DNA was performed using a Rotor-Gene 3000 PCR cycler (Corbett Research, Mortlake, Australia) and a Rotor-Gene™ SYBR Green kit according to the manufacturer's instructions. The primer pair sequences used in this study are shown in Table 1. The amplification reaction was run to complete 40 cycles at 95°C for 20 s, 60°C for 20 s, and 72°C for 20 s. The results were analyzed with Rotor-Gene 6000 series System Software program, version 6 (Corbett Research). The relative expressions of the target genes were normalized to those of glyceraldehyde 3-phosphate dehydrogenase (GAPDH).

Statistical analyses
All results are presented as the mean and standard error of the mean. Statistical analyses were performed using the Student's t-test and analysis of variance. Differences between treatment groups were analyzed by using Duncan's multiple range test. Differences were considered significant at P < 0.05. Statistical analyses were undertaken using Statistical Analysis System for Windows version 9.4 (SAS Institute, Cary, NC, USA).
RESULTS
Characterization of the 4 extracts
To characterize each of the 4 extracts (AL, EL, AS, and ES), total phenolic compound and total flavonoid content levels in each extract were determined and are shown in Table 2. Of the 4 extract types, AL had the highest total phenolic compounds and total flavonoid content levels. AL contained 69.5 ± 0.2 mg (GAE)/g of total phenolic compounds and 27.1 ± 2.9 mg (QUE)/g of total flavonoids (Table 2).
Table 2
Total phenolic compounds and total flavonoids contents in P. japonicus extracts

| AL | EL | AS | ES | |
|---|---|---|---|---|
| Total phenolic compounds (mg [GAE]/g) | 69.5 ± 0.2 | 28.7 ± 0.4 | 12.4 ± 0.1 | 5.2 ± 0.3 |
| Total flavonoids (mg [QUE]/g) | 27.1 ± 2.9 | 14.7 ± 1.2 | 3.3 ± 0.5 | 3.1 ± 0.1 |
Data are shown as mean ± standard error of the mean.
AL, the aqueous extract of P. japonicus leaves; AS, the aqueous extract of P. japonicus stems; EL, the ethanolic extract of P. japonicus leaves; ES, the ethanolic extract of P. japonicus stems; GAE, gallic acid equivalents; QUE, quercetin equivalents.
Effect of each extract on the viability of MC3T3-E1 cells
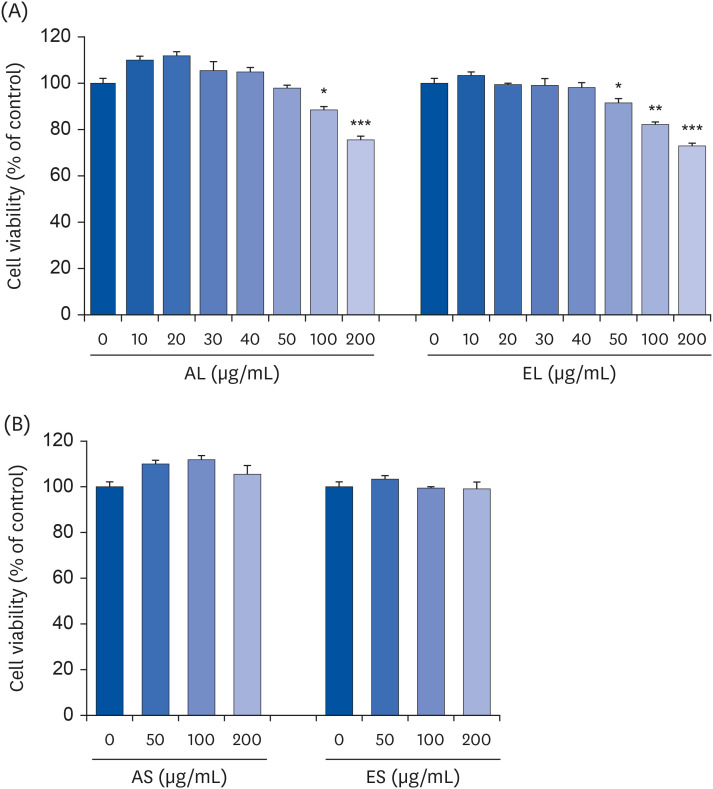
In order to determine the concentration of each extract that could not cause cytotoxicity in MC3T3-E1 cells, cells were incubated for 72 h in culture media containing 0–200 µg/mL of each extract, followed by MTT assay. The viability of MC3T3-E1 cells was significantly decreased when the concentrations of AL and EL were above 100 and 50 µg/mL, respectively (Fig. 1A). On the other hand, AS and ES did not affect cell viability in the 0–200 µg/mL concentration range (Fig. 1B). To exclude possible cytotoxic effects, we used AL and EL at 0–40 µg/mL concentrations and AS and ES at 0–200 µg/mL concentrations in subsequent experiments.
Fig. 1
Effects of four P. japonicus stem and leaf extracts on the viability of MC3T3-E1 cells.
MC3T3-E1 cells were plated at a density of 5 × 104 cells/well in 24-well plates with α-MEM supplemented with 10% FBS. Twenty-four h after plating, the cells were incubated for 72 h in media containing 0–200 µg/mL of each extract. Cell viability was estimated by using the MTT assay. Each bar represents the mean ± standard error of the mean (n = 3).
α-MEM, α-minimal essential medium; FBS, fetal bovine serum; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; AL, the aqueous extract of P. japonicus leaves; ES, the ethanolic extract of P. japonicus leaves; AS, the aqueous extract of P. japonicus stems; ES, the ethanolic extract of P. japonicus stems.
*P < 0.05, **P < 0.01, and ***P < 0.001 indicate significant differences from the 0 µg/mL group.
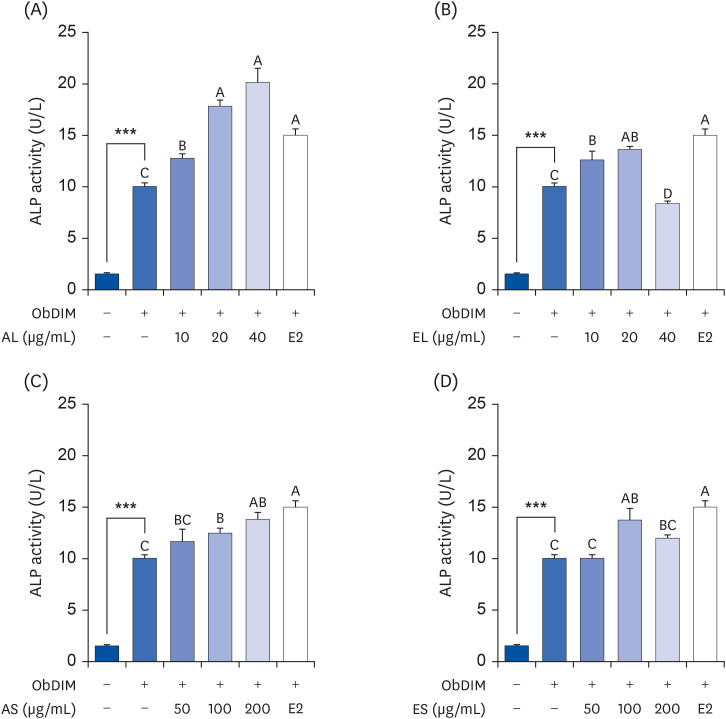
AL is the most potent among P. japonicus extracts in increasing ALP activity in osteoblasts
First, we estimated ALP activity, an early-stage marker of the osteoblast differentiation process, in extract-treated MC3T3-E1 cells, to select the extract that was most effective in promoting osteoblast differentiation among the 4 extract types. As shown in Fig. 2, ALP activity was remarkably increased by ObDIM. Treatment of cells with 100 nM E2, a positive control, significantly increased the ObDIM-induced ALP activity. ObDIM-induced ALP activity levels were increased by all extracts when treated at varying concentrations, except when EL at a concentration of 40 µg/mL was treated. Of the 4 extract types, AL most effectively increased ObDIM-induced ALP activity. At 40 µg/mL, AL markedly increased ObDIM-induced ALP activity by 101%, compared to the non-AL-treated control (Fig. 2).
Fig. 2
Effect of four P. japonicus stem and leaf extracts on ALP activity in MC3T3-E1 cells.
MC3T3-E1 cells were plated at 2 × 104 cells/well in 48-well plates and incubated for 24 h. After incubation for 24 h, cells were incubated for 5 days in ObDIM containing various concentrations of each extract and E2. ALP activity was measured. Each bar represents the mean ± standard error of the mean (n = 4).
ALP, alkaline phosphatase; ObDIM, osteoblast differentiation-inducing medium; AL, the aqueous extract of P. japonicus leaves; ES, the ethanolic extract of P. japonicus leaves; AS, the aqueous extract of P. japonicus stems; ES, the ethanolic extract of P. japonicus stems; E2, 100 nM 17-estradiol.
***P < 0.001 significantly different from the ObDIM-untreated group. Means without a common letter differ significantly at P < 0.05.
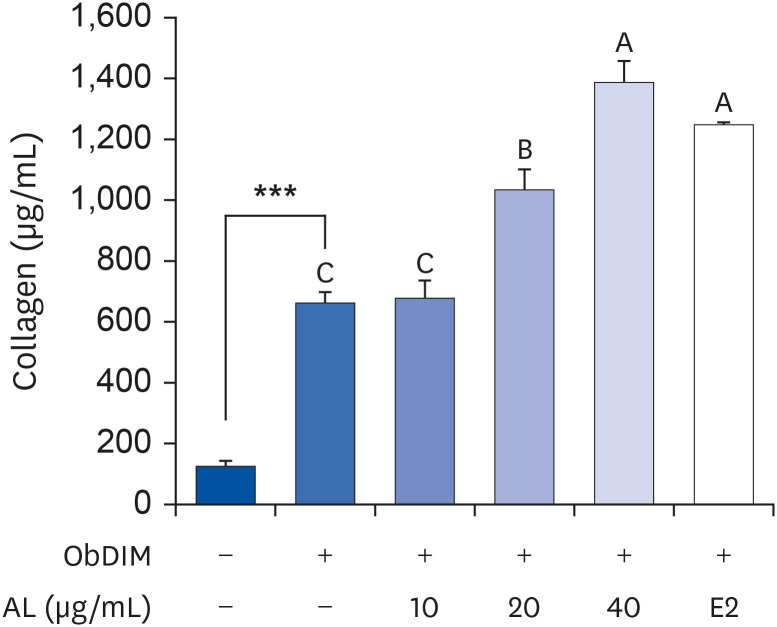
AL promotes collagen synthesis in osteoblasts
To further investigate the effect of AL on the early-stage osteoblast differentiation process, collagen synthesis was estimated by Sirius Red staining. Similar to the ALP activity results, collagen synthesis was dramatically increased by ObDIM. Treatment of cells with 20 or 40 µg/mL AL significantly increased ObDIM-induced collagen synthesis. At 40 µg/mL, AL increased ObDIM-induced collagen synthesis by 109% compared to the non-AL-treated control (Fig. 3).
Fig. 3
Effect of P. japonicus leaf extract on collagen synthesis in MC3T3-E1 cells.
MC3T3-E1 cells were plated at 2 × 104 cells/well in 48-well plates and incubated for 24 h. After incubation for 24 h, cells were incubated for 8 days in ObDIM containing various concentrations of AL and E2. Collagen synthesis was measured by Sirius Red staining. Each bar represents the mean ± standard error of the mean (n = 4).
ObDIM, osteoblast differentiation-inducing medium; AL, the aqueous extract of P. japonicus leaves; E2, 100 nM 17-estradiol.
***P < 0.001 significantly different from ObDIM-untreated group. Means without a common letter differ significantly at P < 0.05.
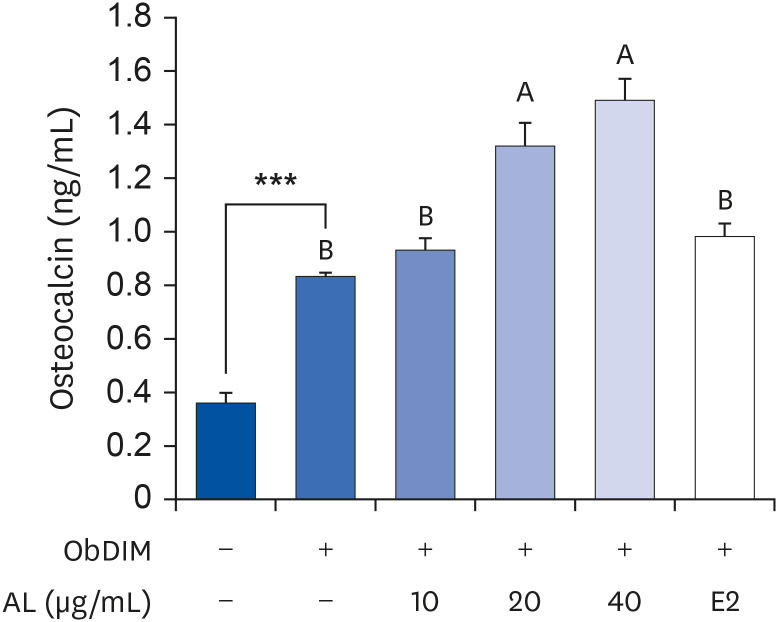
AL encourages mineralization and osteocalcin production in osteoblasts
Next, we measured the degree of mineralization and the production of osteocalcin to investigate the effect of AL on the late-stage osteoblast differentiation process. As depicted in Figs. 4 and 5, ObDIM markedly increased mineralization and osteocalcin production in MC3T3-E1 cells. Treatment of cells with AL significantly increased ObDIM-induced mineralization. However, ObDIM-induced mineralization showed no significant difference among the AL-treated groups at different concentrations (Fig. 4). When concentrations of 20 or 40 µg/mL AL were treated, ObDIM-induced osteocalcin production was significantly increased compared to the non-AL-treated control. Treatment of cells with 10 µg/mL AL showed no significant difference in osteocalcin production compared to the non-AL-treated control (Fig. 5).
Fig. 4
Effect of P. japonicus leaf extract on mineralization in MC3T3-E1 cells.
MC3T3-E1 cells were plated at 2 × 104 cells/well in 48-well plates and incubated for 24 h. After incubation for 24 h, cells were incubated for 14 days in ObDIM containing various concentrations of AL and E2. The degree of mineralization was measured by Alizarin Red S staining. Each bar represents the mean ± standard error of the mean (n = 4).
ObDIM, osteoblast differentiation-inducing medium; AL, the aqueous extract of P. japonicus leaves; E2, 100 nM 17-estradiol.
***P < 0.001 significantly different from ObDIM-untreated group. Means without a common letter differ significantly at P < 0.05.
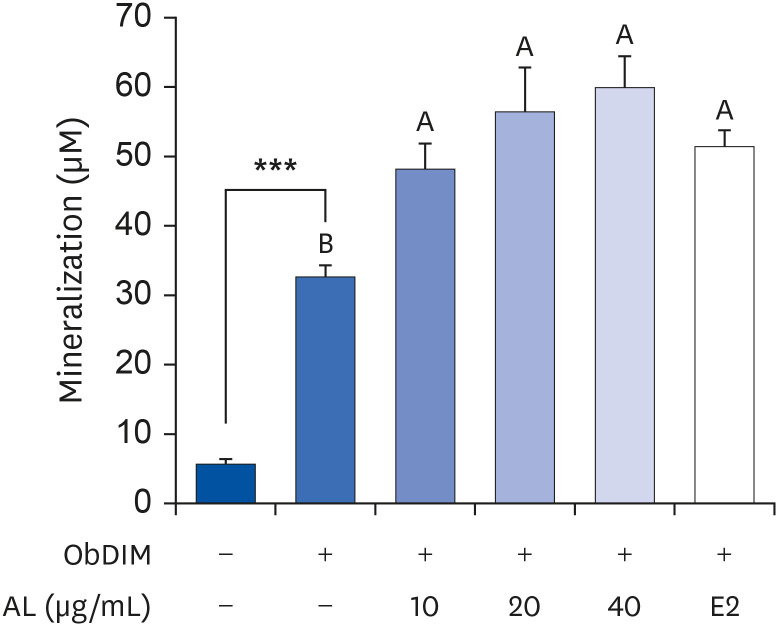
Fig. 5
Effect of P. japonicus leaf extract on osteocalcin production in MC3T3-E1 cells.
MC3T3-E1 cells were plated at 2 × 104 cells/well in 48-well plates and incubated for 24 h. After incubation for 24 h, cells were incubated for 14 days in ObDIM containing various concentrations of AL and E2. Media conditioned for 24 h were collected. Levels of osteoblast-secreted osteocalcin in 24 h-conditioned media were measured. Each bar represents the mean ± standard error of the mean (n = 4).
ObDIM, osteoblast differentiation-inducing medium; AL, the aqueous extract of P. japonicus leaves; E2, 100 nM 17-estradiol.
***P < 0.001 significantly different from ObDIM-untreated group. Means without a common letter differ significantly at P < 0.05.
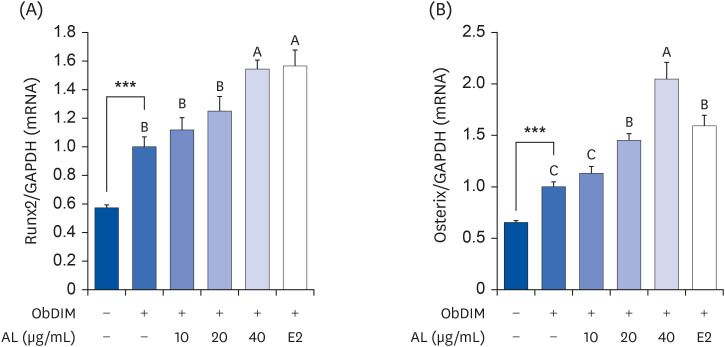
AL upregulates Runt-related transcriptional factor 2 (Runx2) and Osterix expressions in osteoblasts
To investigate the effect of AL on the expression of osteogenic transcriptional factors, Runx2 and Osterix mRNA expressions were determined by performing real-time RT-PCR. Similar to other results, the mRNA expressions of Runx2 and Osterix were dramatically upregulated by ObDIM. The mRNA expression of Osterix upregulated by ObDIM was augmented by AL treatment at 40 µg/mL (Fig. 6A). In addition, treatment of cells with 20 or 40 µg/mL AL significantly increased the ObDIM-upregulated mRNA expression of Runx2 in osteoblasts (Fig. 6B).
Fig. 6
Effect of P. japonicus leaf extract on Runx2 and Osterix mRNA expressions in MC3T3-E1 cells.
MC3T3-E1 cells were plated at 2 × 104 cells/well in 48-well plates and incubated for 24 h. After incubation for 24 h, cells were incubated for 3 days in ObDIM containing various concentrations of AL and E2. Total RNA in cells was extracted, reverse transcribed, and real-time PCR was conducted. Relative mRNA expression levels of Runx2 and Osterix in MC3T3-E1 cells were analyzed. Targeted mRNA expression was normalized to that of GAPDH and is presented relative to the ObDIM-treated group. Each bar represents the mean ± standard error of the mean (n = 4).
Runx2, Runt-related transcriptional factor 2; ObDIM, osteoblast differentiation-inducing medium; AL, the aqueous extract of P. japonicus leaves; E2, 100 nM 17-estradiol; PCR, polymerase chain reaction; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
***P < 0.001 significantly different from the ObDIM-untreated group. Means without a common letter differ significantly at P < 0.05.
DISCUSSION
In this study, when the leaves and stems of P. japonicus were extracted with hot water or 95% ethanol, both extract types were shown to contain large amounts of total phenolic compounds and total flavonoids. This was more so in P. japonicus leaves than in stems and in the aqueous extract than in ethanol (Table 2). It has been reported that P. japonicus contains flavonoids, sesquiterpenes, triterpenes, and various types of phenolic compounds [17]. According to a recent study by Ahn et.al. [8], 26 polyphenolic compounds were detected in a P. japonicus ethanol extract. Among the phenolic compounds, the amounts of 5-O-caffeoylquinic acid and fukinolic acid were large, and the content of kaempferol was the highest among the flavonoids present. The contents of quercetin, astragalin, and nocotiflorin were high as well. In a methanol extract of P. japonicus, kaempferol and quercetin [11] and fukinolic acid, fukiic acid, and other hydroxycinnamoylquinic acid derivatives [23] were detected. In a hot water extract of P. japonicus, petasitesins, cimicifugic acid, fukinolic acid, and dicaffeoylquinic acids were detected [17].
There is abundant evidence that dietary polyphenols have multiple health benefits in various diseases from infectious diseases to chronic metabolic syndrome and cancer. The health benefits of P. japonicus, such as radical scavenging activities, anti-inflammatory activities, and anti-allergic properties, are also considered to be due to polyphenolic compounds contained in P. japonicus (reviewed in [1]). Recently, it was reported in several in vitro and in vivo experiments that kaempferol has osteoprotective effects (reviewed in [1]). Kaempferol's wide range of health benefits, such as antioxidant, anti-inflammatory, anti-obesity, and anti-cancer, have been known for a long time [2425]. Kaempferol has been reported to exert anti-osteoporotic effects by regulating estrogen receptor, bone morphogenetic protein-2, nuclear factor-kappa B, mitogen-activated protein kinase, and mammalian target of rapamycin signaling pathways (reviewed in [1]). In this study, the HPLC-based analysis of AL showed that various polyphenol components were detected. Although kaempferol was also detected, its content in the AL extract was lower than that in the other extracts (data not shown). Although the exact composition of AL was not identified in our study, it was shown that AL contains a considerable amount of polyphenol compounds. Therefore, it can be inferred that these polyphenol components are involved in the functionality of P. japonicus.
All of the 4 extracts of P. japonicus (water and ethanol extracts from leaves and stems) increased ALP activity in MC3T3-E1 cells, and among those effects, AL was particularly effective (Fig. 2). We used only AL as the treatment component in subsequent experiments because AL had the highest content levels of total phenolic compounds and flavonoids, and its ALP activity increase was the greatest. AL increased collagen synthesis dose-dependently at concentrations of 10–40 μg/mL (Fig. 3). These results suggest that AL has a promotional effect in the early-stage of the osteoblast differentiation process. We analyzed osteocalcin production and the degree of mineralization to investigate whether P. japonicus also affects the osteoblast differentiation process in the late-stage. The results showed that when MC3T3-E1 cells were treated with 10 to 40 μg/mL of AL, both degrees of mineralization and osteocalcin production increased (Figs. 4 and 5). These results imply that AL can promote the osteoblast differentiation process in both the early and late stages.
Bone is actively maintained by a coordinated balance between osteoblasts and osteoclasts. Osteoblasts perform bone formation functions while osteoclasts perform bone resorption functions [26]. Osteogenesis is precisely regulated by internal and external factors, including hormones, growth factors, transcriptional factors, and signaling pathways related to bone formation. The molecular mechanisms of bone formation involve 3 main steps: proliferation, extracellular matrix maturation, and mineralization [27].
Differentiation of osteoblasts is regulated by transcription factors. Runx2 is one of the most important transcription factors essential in the osteogenesis process and acts in the differentiation of mesenchymal cells into osteoblasts and chondrocytes [28]. Runx2 is the furthest upstream transcription factor in the regulation of osteoblast differentiation [29]. Its expression is upregulated in immature osteoblasts but downregulated in mature osteoblasts. Runx2 promotes the differentiation of multipotent mesenchymal cells into immature osteoblasts, resulting in immature bone formation, maintaining cells in the immature stage (preosteoblasts). Runx2 triggers osteocalcin expression by binding to the cis-acting element of the osteocalcin promoter region [30]. Osterix (also known as Sp7) is a zinc finger-containing osteoblast-specific transcription factor, which is also essential for the differentiation and proliferation of osteoblasts [31]. Osterix is required for the final commitment of the osteoblast lineage [32]. Osterix is directly involved in mesenchymal cells turning into osteoblasts, blocking the differentiation of mesenchymal cells into chondrocytes. Moreover, Osterix promotes the maturation of preosteoblasts and their differentiation into osteoblasts [33]. Osterix gene expression can be induced directly or indirectly; that is, Msx2 directly induces Osterix, while BMP2 indirectly induces Osterix through Runx2 and Dlx5. Once Osterix expression is triggered, then the expression of proteins involved in the production of osteoblasts, such as type 1 collagen, osteonectin, osteopontin, and bone sialoprotein, increases [29].
In this study, we investigated whether AL's effect on osteoblastogenesis is due to its influence on osteoblast transcription factors Runx2 and Osterix. As shown in Fig. 6, when MC3T3-E1 cells were treated with 20 or 40 µg/mL AL, the mRNA expressions of Runx2 and Osterix were significantly increased. Therefore, we observed that AL increases the expression of osteoblast transcription factors, promotes the synthesis of collagen and osteocalcin, and eventually increases mineralization.
In conclusion, among the 4 P. japonicus extracts, the aqueous leaf extract had the highest total phenolic compound and total polyphenol content. It also produced the highest ALP activity, which is a major biomarker of osteoblast differentiation. In MC3T3-E1 cells that induced osteoblast differentiation, collagen content, mineralization, and osteocalcin production were significantly increased, and expressions of Runx2 and Osterix mRNA, transcription factors that regulate osteoblast differentiation, were also significantly increased. These results indicate that AL regulates osteoblast differentiation at least in part through Runx2 and Osterix. Therefore, an aqueous extract of P. japonicus leaves is highly likely to be useful as an alternative therapeutic for preventing and treating osteoporosis. In further studies, it is necessary to identify the functional components of P. japonicus leaves and to verify whether it has similar effects in in vivo studies.
 XML Download
XML Download