

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Ankylosing spondylitis (AS) is a chronic inflammatory disease characterized by new bone formation that eventually leads to ankylosis of the spine and sacroiliac joints [1,2]. Clinically, the most striking feature of AS is enthesitis, which is characterized by inflammation of a ligament, tendon, or ligament attachment site and diffuse bone edema following soft tissue inflammation. Enthesitis is considered to have pivotal role in initiation and progression of bony ankylosis in AS [3]. However, little is known about the underlying molecular mechanism of bony ankylosis and enthesitis in AS.
Among several important pro-inflammatory cytokines, tumor necrosis factor (TNF) is thought to play a key pathophysiological role in AS. Increased expression of TNF has been reported in the serum or affected sacroiliac joint of AS patients [4,5]. Anti-TNF treatment, which is widely used in AS, not only improves clinical outcomes but also reduces radiographic progression [6]. Therefore, to improve our understating of the underlying mechanisms of bony ankylosis in AS, there is need for a thorough investigation of the enthesis response to TNF.
Dickkopf 1 (DKK1) is an inhibitory molecule of the Wnt/β-catenin signaling pathway, which can stimulate new bone formation by regulating osteoblastogenesis and osteoclastogenesis [7]. Since new bone formation is a key pathological feature of AS, several efforts have been made to elucidate whether DKK1 is involved in pathogenesis of AS [8-11]. However, previous studies have shown controversial results regarding the serum DKK1 levels in AS patients [12,13]. In addition, the relationship between TNF and DKK1 expression in enthesis and its underlying molecular mechanism targeting bone formation of AS have not been reported so far.
In this study, we sought to determine the serum DKK-1 levels in AS patients and elucidate the underlying mechanism of TNF-mediated DKK1 expression in enthesis cells. We also reported here that DKK1 overexpression is involved in bone formation in enthesis. These results suggest that TNF-induced DKK1 expression may play an important role in pathological bone formation in AS.
Go to : 
MATERIALS AND METHODS
Ethical approval
This study was performed in accordance with the recommendation of institutional guidelines and the Ethics Committee of Hanyang University Hospital (IRB 2014-05-002) and Hanyang University Guri Hospital (IRB 2014-05-001). Written informed consent was obtained from all subjects.
Patients
We obtained sera from 103 patients with AS (mean age 45.4±5.0 years) and 30 healthy controls (mean age 32.1±5.2 years). All of the AS patients and healthy controls (HCs) was male. All AS patients were satisfied with modified New York criteria [14].
Facet joint entheses were obtained from four AS patients (mean age 42.1±5.7 years, 2 males) undergoing spinal correction surgery and five controls (mean age 56±11.8 years, 3 males) who underwent spinal surgery for non-inflammatory spinal diseases. All AS patients were HLA-B27 positive patients. Two patients (50%) were treated with biologic agents [15].
DKK1 measurement
The human sera from the AS and HCs were collected in a separator tube and allowed to clot for an hour at 4°C. The samples were then centrifuged for 15 minutes at approximately 3,000 rpm at 4°C. All sera were immediately aliquoted and stored at −80°C. The collected sera were tested for human DKK1 (DB100B; R&D, Minneapolis, MN, USA) by a commercial ELISA kit according to the manufacturer’s protocols. Since the classic DKK1 standard curve ranges are from 0∼2,000 pg/mL, we fixed the values as 2,000 pg/mL if the DKK1 levels go over 2,000 pg/mL.
Human enthesis cells and bone formation in vitro
The isolation and characteristics of the primary enthesis were previously described [15]. Briefly, the spinal bones were repeatedly washed with pre-chilled 1× PBS twice to remove the suspension cells. The excess or adjuvant soft tissues of the bone were collected. The collected enthesis tissues were incubated in DMEM (10082147; Gibco, Waltham, MA, USA) containing collagenase type 1 (17100-017; Gibco) and antibiotics (15140-122; Gibco) with shaking. On the next day, the isolated cells were seeded and used for experiments in the second or fourth passages. Before the cell stock storage, the primary enthesis cells were checked for mycoplasma using PCR methods (6601; Takara, Kyoto, Japan). To induce bone formation, the cells were seeded in DMEM medium and then stimulated with the conditional medium including ascorbic acid, β-glycerolphosphate, and dexamethasone. The conditional medium was exchanged every three days. The experimental design and measurement methods for changes in bone formation are shown in Figure 2A.
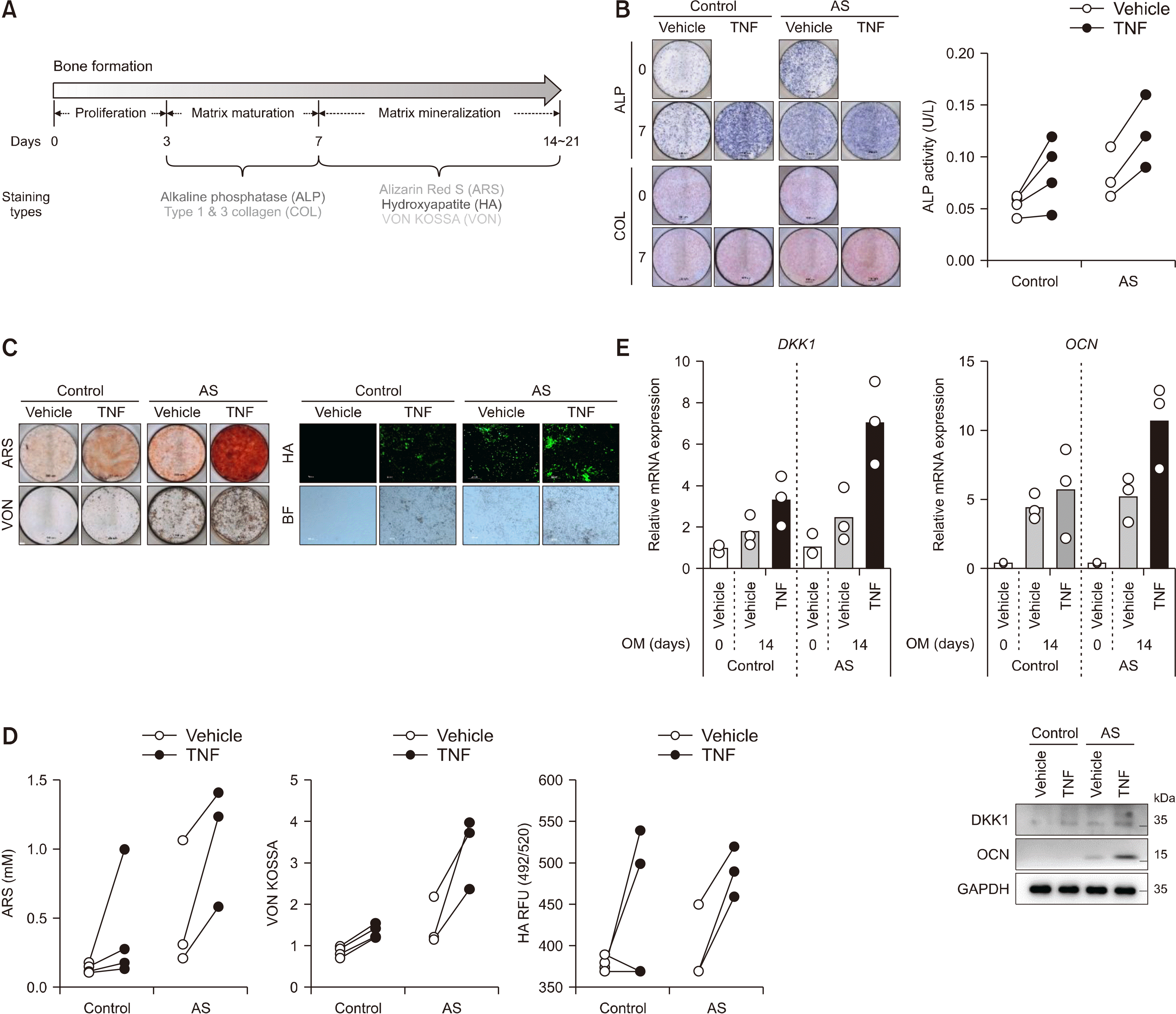 | Figure 2Treatment of TNF promoted bone formation of enthesis. (A) Experimental design for inducing bone formation in primary enthesis cells. Both control- and AS-enthesis cells were followed by bone formation inducing conditional medium for the indicated days. As indicated days, the differentiated cells were subjected to various experimental bone forming activities of (B) ALP and Collagen (COL) staining using Sirius red (left) and intracellular ALP activity (right), (C) Alizarin Red S (ARS), Von Kossa (VON), and Hydroxyapatite (HA) stain images, and (D) Quantification of bone formation indicators stain; ARS, VON, and HA (control=4, AS=3). All of the staining images is representative of three independent experiments. (E) RT-qPCR and immunoblotting results of differentiated enthesis cells at 14 days (control=3, AS=3). Each dots display individual values. TNF: tumor necrosis factor, AS: ankylosing spondylitis, ALP: alkaline phosphatase.
|
Reagents
Recombinant human TNF (300-01A; Peprotech, Cranbury, NJ, USA) and BAY 11-7082 (B5556; Sigma, St. Louis, MO, USA) were used.
Luciferase assay
The 293T cells were co-transfected with the luciferase gene tagged human promoter and firefly plasmid using Lipofectamine 3000 (L3000008; Thermo, Waltham, MA, USA). The cells were then incubated for 48 hours and analyzed by the firefly luciferase activity (E1500; Promega, Madison, WI, USA) according to the manufacturer’s protocol. Luciferase activity was measured using a Luminometer (Berthold; Bad Wildbad, Baden-Württemberg, Germany). Two types (1 kb and 0.35 kb) of Human DKK1 promoters, three types (WT, S536A, S5336E) of human NF-κB promoters, and firefly were generously obtained from Prof. Jeong-Yeon Lee (Department of Pathology, College of Medicine, Hanyang University, Seoul, Korea) [16,17].
Quantitative reverse transcriptase-PCR
The complementary DNA were generated using reverse transcriptase (K1622; Thermo). Quantitative PCR (qPCR) was performed on a CFX96 Real-time thermocycler from Bio-Rad using iQ SYBR Green Supermix (170-8882AP; Bio-Rad, Hercules, CA, USA) according to the manufacturer’s protocol. For all samples, the target gene expression was normalized to GAPDH. The oligonucleotide primer sequences are given in Table 1.
Table 1
Primer sequences for qPCR
![]()
Immunoblotting
For the immunoblot analysis, the cell pellets were lysed in RIPA buffer (50 mM Tris-HCl [pH 8.0], 150 mM NaCl, 0.1% SDS, 0.6% Na-deoxycholate, 1% Triton X-100) that was supplemented with protease (535140; Calbiochem, St. Louis, MO, USA) and phosphatase (5870; Cell signaling, Denvers, MA, USA) inhibitor cocktails. The lysed samples were incubated on ice for 30 minutes followed by centrifugation at 12,000 g for 30 minutes at 4°C. The lysates were separated by SDS-PAGE and electrophoretically transferred to nitrocellulose membranes (10600002; GE Healthcare, Chicago, IL, USA) in a transbuffer. The membranes were blocked with 5% non-fat milk in Tris-buffered saline with 0.1% Tween 20 and incubated with specific primary antibodies, followed by incubation with horseradish peroxidase-conjugated secondary antibodies. The antibodies that were used are given in Table 2. The membranes were visualized with Pierce ECL (32106; Thermo), and Uvitech System (UVITEC, Cambridge, UK).
Table 2
Primary antibodies used in IB, IF, and ChIP
![]()
Immunofluorescence
Immunofluorescence was performed as previously described [18]. Isolated enthesis cells were grown on cover glasses and fixed with 10% formalin for 10 minutes. The cells were then permeabilized with 0.3% Triton X-100 and in 3% bovine serum albumin (BSA) in 1× PBS for an hour at room temperature. The cells were then incubated with the appropriate primary DKK1 antibody in antibody diluent (S3022; DAKO, Carpinteria, CA, USA) overnight at 4°C. Antibody binding was visualized using the Cy3-conjugated anti-rabbit secondary antibody (111-165-144; Jackson Immunoresearch, West Grove, PA, USA). The nuclei were counterstained with DAPI (H-1200; VECTASHIELD, Burlingame, CA, USA). To visualize the stained cells, a fluorescence microscope (Leica Microsystems, Wetzlar, Germany) was used. The primary antibodies that were used are given in Table 2.
Chromatin immunoprecipitation-quantitative PCR (ChIP-qPCR)
ChIP assays (17-295; Millipore, Burlington, MA, USA) were performed as previously described [19]. In brief, the cells were stimulated in 10 cm dishes and fixed with a final concentration of 1% formaldehyde. These cells were sonicated in 1× PBS with protease inhibitor and then immunoprecipitated with protein A agarose beads conjugated to the phos-NF-κB antibody. The DNA purification was performed using the phenol/chloroform/isoamyl alcohol (P3803; Sigma) method. The DNA was precipitated using 3 M sodium acetate and eluted with DEPC water. Approximately 5∼10 ng of eluted DNA was used for qPCR. The quantification and calculation of ChIP-qPCR have been previously described [20]. The DKK1 promoter sequences were obtained from the Eukaryotic Promoter Database (EPD, http://epd.epfl.ch). The primers used in the ChIP-qPCR are provided in Table 1.
Statistical analysis
Data are presented as means±standard deviations, medians (interquartile range, IQR), or percentages, as appropriate. Differences of age between the two groups were assessed using the Student’s t-test. Comparison for serum DKK1 levels between the two groups was assessed by analysis of covariance (ANCOVA) model after adjusting for age. Statistical analyses were conducted using SPSS (version 26.0; IBM Co., Armonk, NY, USA) or GraphPad Prism 6 software. Graphical data representation was performed using GraphPad Prism 6 or 9 software. p-values<0.05 were considered statistically significant.
Go to :
RESULTS
Serum DKK1 levels were higher in AS than HCs
AS patients were older than those in the HC group (45.4±5.0 years vs. 32.1±5.2 years, p<0.001). The median (IQR) DKK1 levels in AS patients were also significantly higher than those of the HCs after adjusting for age (917.4 [615.3∼1,310.0] pg/mL vs. 826.2 [670.3∼927.8] pg/mL, p=0.043). However, the broad range was shown in the serum DKK1 levels of AS patients (Figure 1).
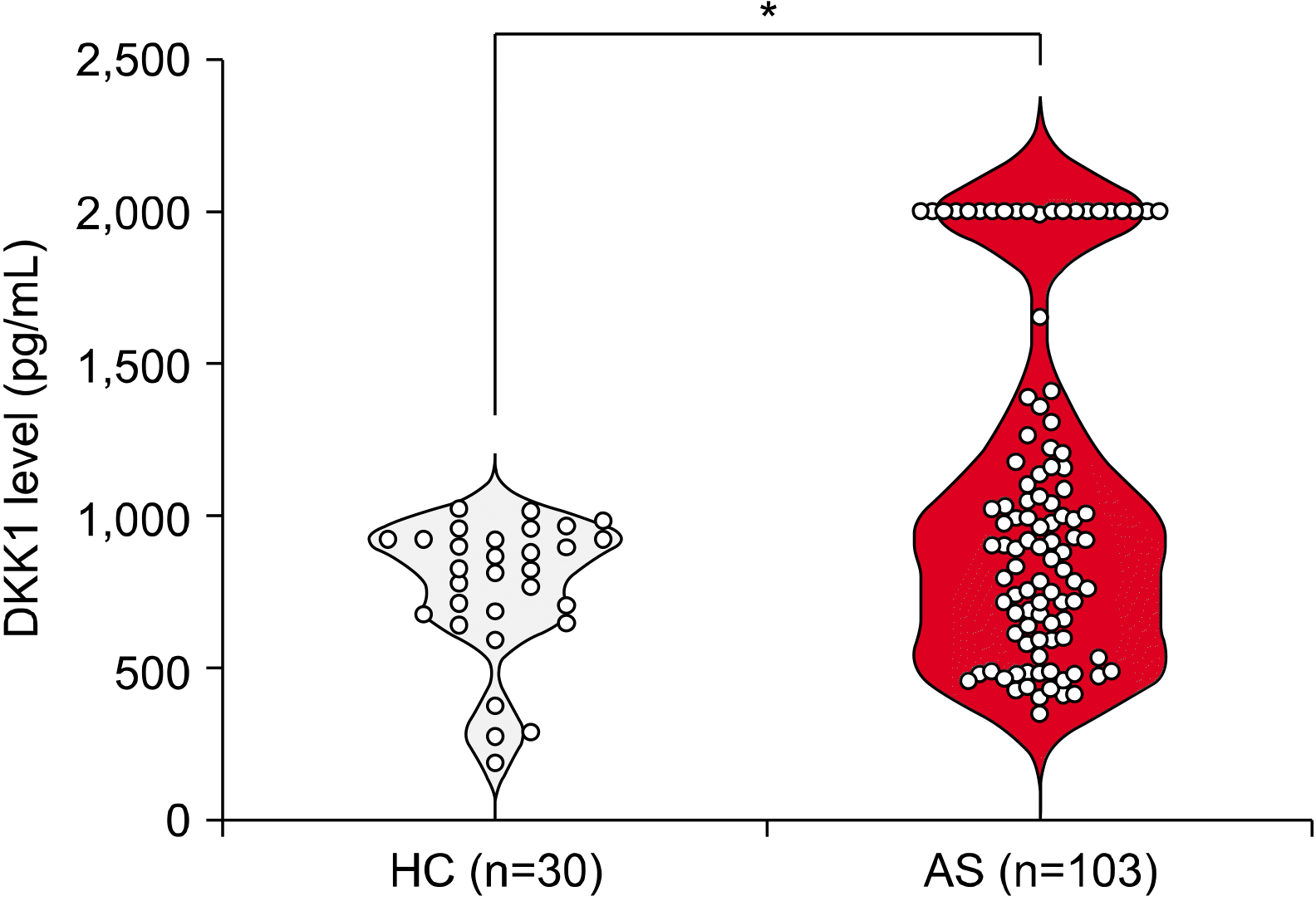 | Figure 1Serum DKK1 levels were higher in ankylosing spondylitis (AS) patents than those of control patients after adjusting for age. The human DKK1 levels were measured in the collected sera (healthy control [HC], n=30; AS, n=103) using ELISA. Since the classic DKK1 standard curve ranges 0∼2,000 pg/mL, we fixed the values as 2,000 pg/mL if the DKK1 levels go over 2,000 pg/mL. The bars show means±standard error of the mean; *p<0.05.
|
Treatment of TNF promotes bone formation
The experimental design and analysis tools for bone formation are shown in Figure 2A. Under bone formation conditions, TNF treatment accelerated matrix maturation (alkaline phosphatase [ALP] and collagen [COL] stains) (Figure 2B) and mineralization (Alizarin Red S rheumatoid arthritis synovium, Hydroxyapatite [HA], and Von Kossa [VON] stains) (Figure 2C and 2D) compared to treatment with vehicle in both control and AS enthesis cells. As shown in Figure 2E, the mRNA and protein expressions of DKK1 and OCN increased after TNF treatment in both control and AS enthesis cells. Notably, the accelerated bone formation and DKK1 expression induced by TNF were pronounced in the AS enthesis compared to those of control.
TNF treatment activates the phosphorylation of NF-kB protein to induce DKK1 mRNA expression in enthesis
Previous studies have suggested that TNF is strongly associated with DKK1 expression in TNF transgenic mice and the rheumatoid arthritic synovium [21,22]. To explore the underlying molecular mechanism of TNF-induced DKK1 expression, we treated control enthesis cells with TNF. After TNF stimulation, the mRNA expression of DKK1 was upregulated compared to those of control (Figure 3A). The TNF-treated enthesis exhibited an increase of the DKK1 protein levels, as revealed by immunofluorescence (Figure 3B). Moreover, the TNF treatment induced the phosphorylated-S536 NF-κB and expression of DKK1 proteins in enthesis (Figure 3C). The activity of wild type (WT) NF-κB promoter was upregulated in a dose-dependent manner of TNF treatment, but not in mutants (S536A and S536E) (Figure 3D). Intriguingly, TNF treatment activated two (1 kb and 0.35 kb) types of the human DKK1 promoter, but showed relatively higher response in the 0.35 kb DKK1 promoter (Figure 3E). Furthermore, the phos-S536 NF-κB protein was enriched at two putative binding sites within the 0.35kb DKK1 promoter in response to TNF-treatment (Figure 3F). Conversely, the NF-κB inhibitor (BAY) suppressed TNF-induced DKK1 expression by reducing NF-κB phosphorylation (Figure 3G). Taken together, these results suggest that TNF activates NF-κB protein phosphorylation to induce DKK1 mRNA expression in enthesis.
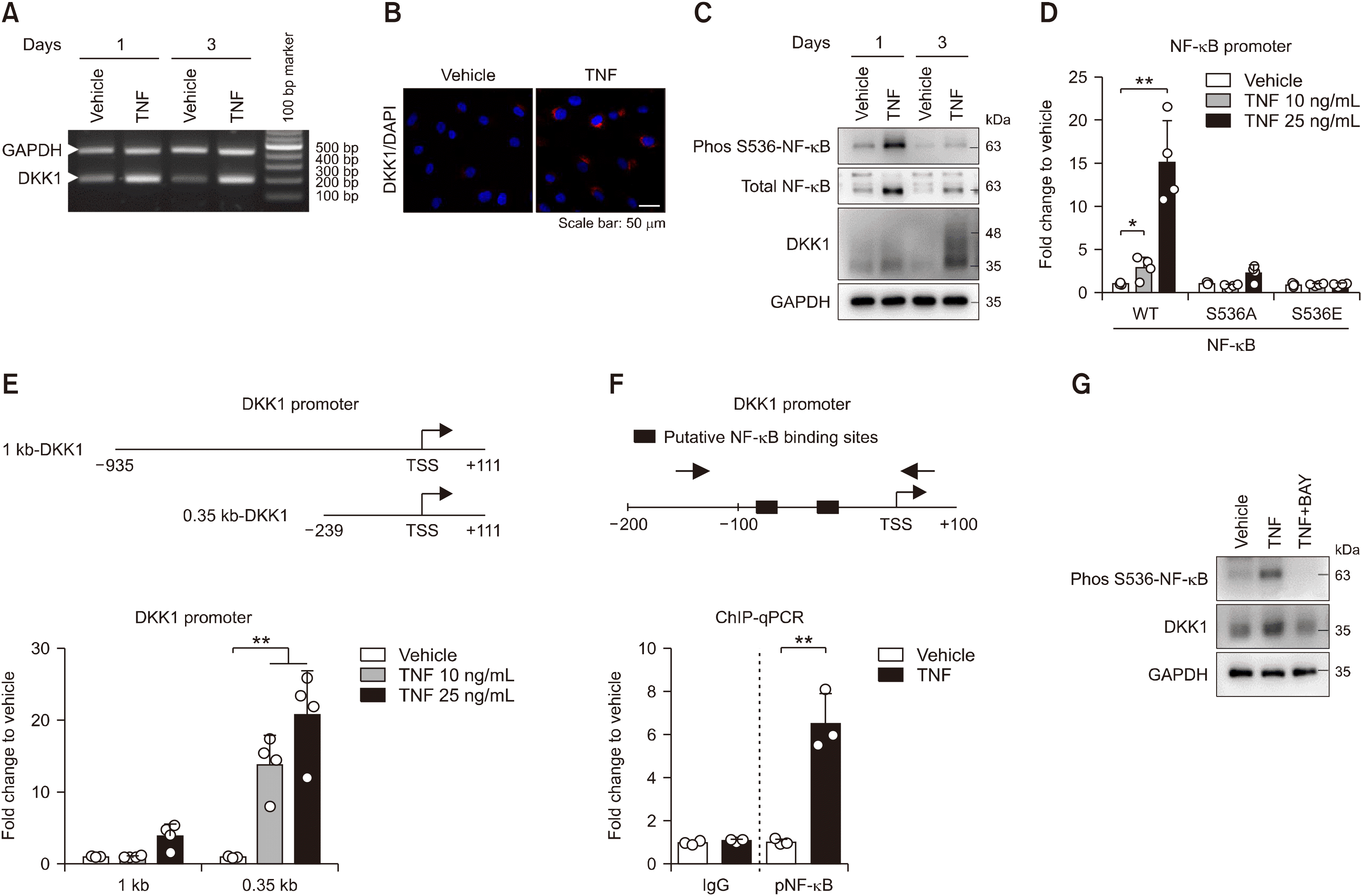 | Figure 3TNF treatment activates NF-κB protein phosphorylation to induce DKK1 mRNA expression in enthesis. Control enthesis cells were stimulated with TNF on days 1 and 3, and analyzed by (A) RT-PCR for DKK1 and GAPDH, and (B) immunofluorescence for DKK1 at day1. (C) Control enthesis cells were stimulated with TNF on days 1 and analyzed by immunoblotting as indicated proteins. (D) Control enthesis cells were co-transfected with three types (WT, S536A mutant, and S536E mutant) of NF-κB, followed by treatment with TNF as indicated doses for 24 hours, and analyzed for luciferase activity (n=4). (E) Control enthesis cells were co-transfected with two types (1 kb and 0.35 kb) of the human DKK1 promoter and beta-gal, followed by treatment with TNF as indicated doses for 24 hours, and analyzed for luciferase activity (n=4). (F) Human primary synovial cells were stimulated with 25 ng/mL TNF for 24 hours, followed by ChIP assays with the phos S536 NF-κB antibody (n=3). (G) Control enthesis cells were stimulated as indicated for a day and analyzed by immunoblotting. Each dot displays an individual value. Values represent means±standard deviations. TNF: tumor necrosis factor, AS: ankylosing spondylitis, ALP: alkaline phosphatase, WT: wild type, ChIP: chromatin immunoprecipitation. *p<0.05, **p<0.01.
|
DKK1 overexpression promotes bone formation
To determine whether DKK1 expression affects bone formation in enthesis, we manipulated DKK1 overexpression in enthesis under bone formation stimulation and observed physiological changes in bone formation. We first confirmed the DKK1 overexpression using RT-qPCR 48 hours after transfection (Figure 4A). There was no significant difference in ALP and COL expression by DKK1 overexpression (Figure 4B), while the calcified nodules and bone formation indicators were increased in DKK1 overexpressed cells compared to controls (Figure 4C and 4D). Taken together, these findings suggest that overexpression of DKK1 promotes bone formation in enthesis.
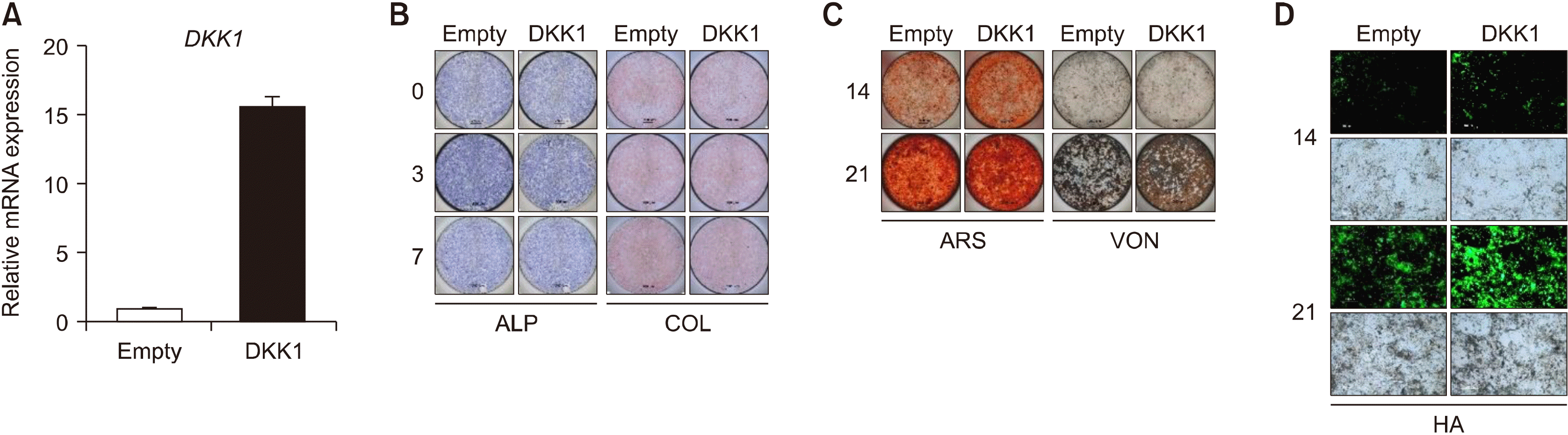 | Figure 4DKK1 overexpression promotes bone formation of enthesis. Enthesis cells were transfected with an empty vector or DKK1 plasmid and then stimulated to osteogenic differentiation. (A) Transfection efficiency was confirmed by RT-qPCR. As indicated days, differentiated cells were subjected to (B) ALP and COL staining, (C) ARS and VON staining, and (D) Hydroxyapatites staining. Representative images are shown. Scale bar is 200 μm. ALP: alkaline phosphatase, COL: collagen, ARS: Alizarin Red S, HA: hydroxyapatite, VON: Von Kossa.
|
Go to :
DISCUSSION
TNF is known to promote bone formation by regulating the expression of various osteogenesis-related genes (such as ALP, RUNX2, and OCN) in various human derived cells such as mesenchymal stem cells, and enthesis cells [23,24], But, it has not been investigated how TNF mediates bone formation in spinal enthesis cells. Here, we reported that AS patients showed higher serum DKK1 levels than those of controls and TNF treatment promoted DKK1 expression and bone formation in enthesis cells. Intriguingly, the TNF-promoted bone formation was more pronounced in AS enthesis cells than those of controls. We also suggested the regulatory molecular mechanism of TNF-induced DKK1 expression in enthesis. DKK1 overexpression in enthesis exhibited an increase in bone formation. Taken together, TNF-induced DKK1 expression is the possible key player in the pathological bone formation of AS.
DKK1 has been recognized as an important regulator of joint remodeling [21]. Due to the bone-forming disease nature of AS, DKK1 has been implicated in the pathophysiology of radiographic AS progression. Interestingly, blocking DKK1 led to sacroiliac joint fusion in an arthritis mice model (TNF transgenic mice), although it did not promote inflammation in the sacroiliac joint [8]. Moreover, a recent study showed that DKK1 significantly reduced the incidence and severity of kyphotic and ankylosing spinal phenotypes in the modified collagen induced arthritis model [25]. Therefore, DKK1 is a well-known potential inhibitor of the Wnt signaling pathway. Elevated DKK1 levels are linked to bone resorption, while decreased levels are associated with new bone formation. However, accumulated data have suggested that DKK1 plays a dual stage-dependent role in osteoblast differentiation. Some studies have shown that DKK1 decreased in extracellular matrix maturation but increased in the mineralization phase of osteoblasts differentiation; therefore, DKK1 is involved in mineralization-related physiological changes [9,26-28]. In this study, we also showed that DKK1 directly promoted bone formation in enthesis. We also identified a discrepancy between our results and the impact of DKK1 on AS-like joint destruction, which was shown in previous studies using murine arthritis models [8,25]. This discrepancy may be associated with a difference between experimentation and reality, or between mice and humans. Alternatively, the difference may reflect a dual role of DKK1 or different activities between functional DKK1 and circulating DKK1. Regardless, further studies are needed to close these gaps.
Previous human studies have produced inconsistent results regarding the serum DKK1 levels in AS patients. In some studies, the serum levels of DKK1 were elevated in AS patients compared to controls [29-31], but reduced [11,32-34] or comparable [10,35,36] in other studies. Thus, relationship between serum DKK1 levels and radiographic changes in AS is still controversial. We found that the serum DKK1 levels of AS were higher than those of controls after adjusting for age. In addition, the DKK1 levels range was wide in AS patients. The previous inconsistence results of the serum DKK1 levels and the wide range of serum DKK1 levels might be associated with a broad range of medical conditions among the patients. Moreover, a recent meta-analysis revealed that the serum DKK1 levels might be associated with the severity of radiological progression or inflammation. In this study, the serum DKK1 levels of AS patients were comparable with that of HCs. However, the serum DKK1 levels in patients with high mSASSS (>30) scores and in patients with abnormal C-reactive protein (>10 mg/L) were significantly lower than that of HCs. In contrast, the serum DKK1 levels in patients with normal CRP (≤ 10 mg/L) were increased [13]. Therefore, further studies are needed to investigate the serum DKK1 levels with consideration of the patients’ medical conditions.
Few studies have suggested that stimulation of TNF induces DKK1 expression in osteoblasts and synovial cells [22,37], but its underlying mechanism is not elucidated so far. To provide molecular mechanism of TNF-regulating DKK1 expression, 0.35 kb of human DKK1 promoter activity was significantly increased depending on dose of TNF treatment, suggesting that 0.35 kb element within DKK1 promoter has a critical role for TNF-induced DKK1 expression (Figure 3E). Additionally, results from ChIP assay showed an increased binding affinity of phos-NF-κB to two putative sites within 0.35 kb promoter by TNF treatment (Figure 3F). Taken together, our results suggest that TNF-activated phos-NF-κB protein binds 0.35 kb within DKK1 promoter to induce DKK1 transcript level.
The present study has several limitations. First, we compared the serum DKK1 levels between AS patients and HCs. However, we did not consider the medical conditions that may influence the DKK1 levels, such as disease activity. Second, the present study showed that TNF induced DKK1 expression by NF-κB activation. However, the regulatory molecular mechanisms and the functional role of DKK1 in TNF-promoted bone formation process remain unclear. Despite these limitations, we confirmed that TNF induces DKK1 expression and bone formation using human AS enthesis cells. In addition, we attempted to identify other inflammatory cytokines that regulate DKK1 expression among various inflammatory cytokines related to AS pathophysiology (such as TNF, IL-1B, IL-6, IL-11, IL-17A, IL-23, IL-32, and LPS). Interestingly, only TNF treatment led to DKK1 upregulation in human enthesis cells (data not shown). These data suggest that TNF plays a key role in pathological bone formation, emphasizing anti-TNF treatment can reduce radiographic progression in AS. We also extended the previously reported functional role of DKK1 in bone formation [26,28]. We provided novel insights into the molecular linkage between TNF and DKK1. Our results widen fundamental understanding of bone formation and the pathogenesis of radiographic progression in AS.
Go to :
CONCLUSION
The serum DKK1 levels of AS patients was higher than that of HCs. The TNF-induced bone formation was remarkable in AS enthesis cells. TNF also induced DKK1 expression by NF-κB activation. In addition to increased bone formation in DKK1 overexpression, our results suggest that TNF-mediated DKK1 plays a role in the radiographic progression of AS.
Go to :
 XML Download
XML Download