

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Hepatic cells synthesize most of the coagulation factors, anticoagulants and constituents of the fibrinolytic system. Tissue factor (TF) is expressed on smooth muscle cells of blood vessels and activates coagulation system [1]. TF is constitutively expressed in blood vessels and its expression is induced in Kupffer cells and stellate cells [2,3]. Initiation of liver TF significantly increased coagulation and tissue injury and the use of anticoagulants have a protective effect against xenobiotics-induced liver toxicities [4,5]. Previous studies, demonstrated increased TF in different pathological conditions [6,7]. Similarly, our previous study indicated an increased TF expression in monocrotaline/lipopolysaccharide (MCT/LPS)-induced idiosyncratic liver toxicity in mice [8].
Retinoids are essential for cell proliferation, differentiation, reproduction, normal immune system and vision [6]. Retinoic acid (RA) Signaling is controlled by DNA binding of homodimer (RXR-RXR) or heterodimer (RAR-RXR) to RA response element RXRE or RARE, respectively [9]. This facilitates transcription of certain genes that regulate cellular apoptosis, differentiation and proliferation [10].
Clinically, RAR agonists are successfully used in the treatment of psoriasis and cancers and experimentally, RXR agonists possess hypoglycemic effect in type II diabetes [11]. Released RAR-α as speckled droplet has been found in vitro in activated hepatic stellate cell (HSC) [12]. The released lipid droplets of RXR-α and RAR-α by MCT/LPS-induced liver injury has been reported and indicated to be TF dependent [13]. Furthermore, both receptors have been confirmed to be released from the pericentral hepatocytes and HSCs as speckled droplets in thioacetamide-induced liver injury in mice [14].
The present study aims to investigate the possible release of RXR-α and RAR-α as speckled lipid droplets in liver toxicity induced by N-acetyl-p-aminophenol (APAP) in mice. In addition, the interdependence between RXR-α, RAR-α and TF will also be clarified. Furthermore, the role of cell cycle-apoptotic marker, caspase-3 and cyclin D1, during liver toxicity induced by APAP in mice will also be explored.
METHODS
Animals and experimental design
Male Swiss albino mice, 20–25 g weight and 5 weeks age, were used. Mice were allowed standard fed and access to water ad libitum. Mice were acclimatized for one week in a cyle of light/dark, 12 h each. Mice were classified into four groups (8 mice each); control group received saline (0.2 ml/animal, i.p.), APAP group treated with a single dose of (APAP, 800 mg/kg, i.p.). The other two groups were injected with TF-antisense (TF-AS) or scrambled TF (TF-SC) deoxyoligonuclutides, ODNs (6 mg/kg ODNs, i.v. in 100 µl saline in a single dose 3.5 h prior APAP treatment) [15]. Mice were euthanized by cervical dislocation, 24 h after APAP injection and liver tissue samples and blood were collected. Liver tissue samples were handled using standard histopathological, immunofluorescence, biochemical procedures. All processes related to animal maintenance, sampling and treatments were in accordance with Al-Azhar laboratory research ethical committee (AZR-19-0143).
Chemicals and antibodies
APAP, chemiluminescent peroxidase substrate kit and HRP-conjugated goat IgG secondary antibody were provided from Sigma-Aldrich (St. Louis, MO, USA). Rabbit polyclonal RXR-α, RAR-α antibodies and TF Rabbit polyclonal were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Rabbit polyclonal antibody for fibrin was purchased from Dako (Carpinteria, CA, USA). TF mouse monoclonal antibody was provided from Thermo Scientific Pierce (Rockford, IL, USA). Cleaved caspase-3 and cyclin D1 rabbit monoclonal antibody were obtained from cell signaling technology (Leiden, Netherlands). Alexa fluor 488 goat anti-mouse and was obtained from Invitrogen (Carlsbad, CA, USA). Cy3 goat anti-rabbit was obtained from Jackson Immunoresearch (West Grove, PA, USA).
ODNs of tissue factor
Antisense and scrambled ODNs for TF were purchased from integrated DNA technologies (Integrated DNA technologies, Coralville, IA, USA). The used sequences in the current study, mouse TF antisense and scrambled ODNs, are as the follow: TF-SC, 5`-TGACGCAGAGTCGTA-3`; TF-AS, 5`-CATGGGGATAGCCAT-3`.
Histopathology and liver tissue samples
Sections of the livers were immerged in Davidson's fixing solution for one day then transferred to 70% ethyl alcohol, subsequently, dehydrated in gradient ethanol concentrations, defatted by xylene and embedded in paraffin. After that, sections were cut to 4 µm thickness and mounted on super-frosted slides.
Immunofluorescence technique
Liver slides were rehydrated by series of ethanol concentrations after being deparaffinized. The sections were the washed using 0.05% tween 80 in phosphate buffered saline (PBS) at 7.4 pH. Tissues were boiled in microwave with DAKO solution, pH 6.0 for 15 min for antigen retrieval step. The sections were incubated with blocking buffer (10% horse in 1% BSA serum in PBS) for 1 h. The sections were then incubated with the suitable 1ry Ab at 37°C for 2 h and then at 4°C overnight. The slides were then washed with washing solution and incubated for 30 min with the suitable secondary Ab conjugated with goat anti-rabbit Cy3 or goat anti-mouse Alexa 488. The nucleus was stained with 1:5,000 4,6-diamidino-2-phenylindole in distilled water and then mounted by fluoromount. Captured figures were analyzed using fluorescence microscope (Leica DM 5500B; Leica Biosystems Inc., Buffalo Grove, IL, USA) and image j software (NIH). The florescence intensity was assessed by measuring its mean reading for three to six fields per section for at least 6 mice per each study group using Imagej/NIH software and confirmed by the microscope software.
Protein extraction and Western blot analyses
In the current work, we performed extraction of the protein and Western blotting analysis. In brief, we used the buffer, Prep protein extraction (catalog #163-2086; Bio-Rad Inc., Hercules, CA, USA) for lysing liver tissue samples of the different groups. Tissue sample lysates was separated and the concentrations of the protein were measured by using Bradford protein assay kit. A 20 µg protein from each sample was mixed with Laemmli sample buffer (loading buffer) and run through 10% sodium dodecyl sulfate–polyacrylamide gels. Blotting to a nitrocellulose membrane (Millipore, Burlington, MA, USA) was performed the gels. The nitrocellulose membrane containing proteins was incubated with polyclonal rabbit TF antibody and after washing with tween-20/PBS, the membrane was incubated with HRP-conjugated goat IgG secondary antibody. Proteins were then visualized by using an enhanced chemiluminescent kit. For equal loading confirmation, membrane was re-incubated with beta actin antibody.
RESULTS
Effect of APAP in the presence or absence of ODNs on hematological and blood chemistry parameters
Fig. 1 and Table 1 show that injection of 800 mg/kg single dose of APAP (i.p.) causes significant increase in serum alanine transaminase (ALT), count of white blood cells (WBCs) and thrombocytopenia in comparison with the vehicle treated (control) group. No significant change is recorded between APAP treated mice or APAP + TF-SC treated group concerning ALT level and WBCs and platelets counts. In contrast, administration of TF-AS before APAP ameliorates the changes in ALT level and WBCs and platelets counts when compared to APAP-treated and APAP + TF-SC groups (Fig. 1A and Table 1).
Effect of APAP with or without ODNs co-treatment on TF and fibrin proteins expression
TF and fibrin proteins show constitutive expression in the sinusoidal area in the pericentral zone in control group (Figs. 1B–E and 2). Injecting the animals with APAP significantly increased TF and fibrin proteins in hepatocyte cells around the central veins. Group treated with APAP + TF-AS demonstrate decreased expression of TF and fibrin, increased by APAP alone. APAP + TF-SC treated mice show non significant change of TF or fibrin compared to APAP-injected mice (Figs. 1 and 2, respectively).
Effect of APAP with or without ODNs co-treatment on RAR-α and RXR-α proteins expression
Immunofluorescence staining revealed a constitutive expression of RAR-α and RXR-α in the pericentral area in vehicle treated group. Additionally, animals treated with APAP and APAP + TF-SC show a scattered expression of both RXR-α and RAR-α in the pericentral zone (as indicated by arrows). Animals injected with TF-AS in the presence of APAP display marked decrease in RXR-α and RAR-α expression around CV (Fig. 3A–D, respectively).
Effect of APAP with or without ODNs co-treatment on cyclin D1 and cleaved caspase-3 proteins expression
Normal mice sections show basal expression of cyclin D1 and cleaved caspase-3. An increased deposition of both proteins predominantly around the CV occurs, 24 h after APAP injection, upon comparison with control group. Cyclin D1and cleaved caspase-3 protein expression is strongly increased in APAP-treated, group mostly in the pericentral area. Similar expression pattern of liver cyclin D1 and cleaved caspase-3 occurs in group treated with APAP and TF-SC. No deposition of cyclin D1 and cleaved caspase-3 is observed in the tissues of APAP + TF-AS injected animals (Fig. 4A–D, respectively).
Effect of APAP in the presence or absence of ODNs on histopathological features
Histopathological evaluation of APAP, in the presence or absence of TF ODNs, treated groups is performed. APAP-treated mice displyed liver injury in the form of alteration of the normal liver architectures. They also show hemorrhage, necrosis, alteration of the sinusoidal architecture and fatty degeneration. APAP + TF-SC treated mice show similar pattern when compared with APAP treated mice. The alteration is more obvious in the pericentral area of the liver than the periportal zone. On the other hand, APAP + TF-AS treated animals demonstrate normal histopathological architectures (Fig. 5).
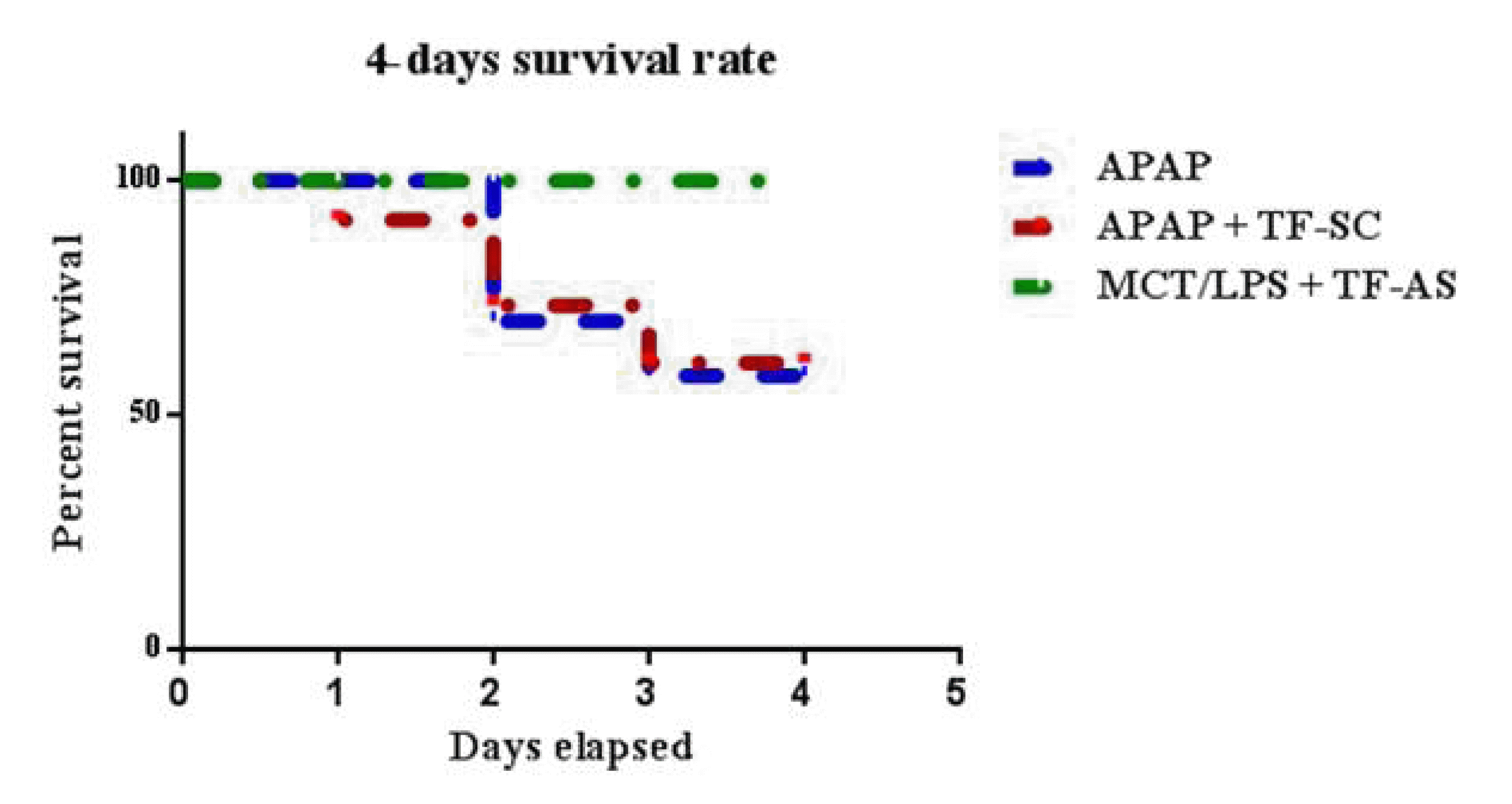
Effect of APAP with or without ODNs co-treatment on animal survivability
In the current study, the animal survival was analyzed in different groups over 4 days. Mice treated with APAP and APAP + TF-SC showed 60% survivability at the end of the experiment. Whereas, mice treated with APAP + TF-AS have 100% survival rate (10/10) (Fig. 6).
DISCUSSION
Several factors appear to play important roles in the liver toxicity induced by APAP such as alterations of the mitochondria, cytokines and reactive nitrogen species or reactive oxygen species [16,17]. Our results indicate that injection of mice with APAP induces liver toxicity; indicated by the significant elevation of serum ALT level and decreased blood platelet with elevated WBCs counts. Moreover, fibrin and TF protein expression is increased in hepatocytes around CV after APAP treatment.
Upon treatment with APAP, excessive WBCs contribute to liver inflammation and injury [18,19]. In addition, liver injury by co-treatment with MCT/LPS in mice lead to migration of platelets from the blood into the injured tissues and attaching fibrin, leading to thrombocytopenia accompanied with elevated WBCs count [15]. Additionally, an earlier report indicated a decreased protein C, plasma antithrombin concentrations and hepatic fibrin accumulation in mice treated with APAP [20]. Another study stated that, the cell surface protein, integrin, is expressed, in vitro, on platelets cell membrane facilitating fibrin binding and forms “cohesive strength” causing platelet to be aggregated with fibrin network [21]. Furthermore, APAP treatment causes elevation of TF in acute liver injured patients [22]. During vascular endothelial cell damage in liver, TF activates the coagulation extrinsic pathway and results in the development of fibrin network [22]. Accordingly, the occurrence of thrombocytopenia along with hypercoagulability in injured liver tissue (high expression of liver TF and fibrin expression) could be explained by the activation of coagulation system induced by APAP in the form of TF which lead to migration of the platelets from the blood into injured tissue causing thrombocytopenia.
HSCs are expressing RAR-α and RXR-α as confirmed by their co-localization with the HSCs marker, α SMA, using double immunofluorescence staining [13]. It has been documented that HSCs loses their vitamin A contents as lipid droplets upon its activation [23]. Moreover, up-regulation of RAR-α protein in activated cultured HSCs, as large insoluble speckled droplets, was seen [12]. Furthermore, retinoids regulate HSC differentiation through activation of RARs and/or RXRs and enhance expression of RAR-α and β that correlates with fibrosis induced by porcine serum [24]. However, no data were found discussing the correlation between the liver injury and loss of retinoid receptors (RXR-α or RAR-α) in APAP-induced liver injury. Abdel-Bakky et al. [13], described that MCT/LPS-induced liver toxicity increased the released lipid droplets containing RXR-α and RAR-α which depend on TF activation. Similarly, it has been established that thioacetamide liver toxicity is associated with loss RXR-α and RAR-α from the pericentral hepatocytes in mice [14]. In the same way, the present data indicate that RXR-α and RAR-α are lost from pericentral cells as lipid droplets after APAP injection in mice (Fig. 3).
APAP toxicity is associated with many common events including apoptosis [25]. The current results display that mice treated with APAP show significant increase in protein expression of cleaved caspase-3 around CV. Interestingly, a previous report showed the involvement of caspase-3 in APAP induced liver toxicity in mice [26]. Also, Wu et al. [27] revealed that increased caspase-3 expression in APAP treated mice was observed around CV. In addition, Irbesartan mitigated APAP-induced caspase-3 expression in mice model [28]. Similarly, Li et al. [29] showed that apoptosis, induced by activated HSCs, is characterized by increased caspase-3 in-vitro. Another significant finding is the increased cyclin D1 protein expression after APAP injection. Earlier study demonstrated that mice treated with lower dose, 300 mg/kg, of APAP increased expression of cyclin D1 while its expression was decreased after using large dose (600 mg/kg) [30]. Unlike the previous study, we used large dose of APAP (800 mg/kg) which resulted in an increased expression of cyclin D1 around the central vein. Increased hepatocytes cyclin D1 expression indicates that the cells are ready to DNA replication (G1/S phase transition) and completed G1 restriction point. It has been demonstrated earlier that Cyclin D1 enhances the “caspase-dependent apoptosis” controlled by the intrinsic pathway of the mitochondria [31]. Accordingly, our data support the concept that, increased cyclin D1expression may potentiate “caspase-3-dependent apoptosis” in APAP treated mice and that increased expression of cyclin D1 could be due to compensatory liver cell proliferation resulting from APAP injection. The current findings show that TF-AS injection efficiently decreases TF and fibrin proteins in the pericentral zone. Also it ameliorates abnormalities of ALT, WBCs and Platelets. In addition, it decreases cyclin D1 and caspase-3 protein expression in liver of mice treated by overdose of APAP and preventes release of RXR-α and RAR-α from pericentral cells. In conclusion, all the above mentioned abnormalities including, thrombocytopenia, pericentral fibrin deposition, enhanced “caspase-3-dependent apoptosis” by cyclin D1 and pericentral loss of RXR-α and RAR-α as lipid droplets in APAP-induced liver injury are TF dependent.




 XML Download
XML Download