

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Irritable bowel syndrome (IBS) is a common functional gastrointestinal (GI) disorder, with a worldwide prevalence of 3–5%, and is more common in women [1]. It is defined as a chronic and recurrent abdominal pain, associated with defecation or changes in the stool form or frequency [2]. It is a functional disorder because it is diagnosed when any detectable organic disease that may explain the symptoms is absent. It is classified into diarrhea-predominant, IBS-D; constipation-predominant, IBS-C; mixed type of diarrhea and constipation, IBS-M; or unclassified, IBS-U; according to the characteristics of the predominant symptoms [3]. Many underlying pathophysiologic factors have been proposed, including altered GI motility, visceral hypersensitivity, abnormal gas propulsion and expulsion, infection, low-grade mucosal inflammation, intestinal permeability change, food intolerance, bile acid malabsorption, central neurohormonal dysregulation, enteric neuroendocrine abnormalities, immunologic or psychologic factors, and genetic factors. However, the exact etiology of IBS has not been fully elucidated [2].
With an increasing interest in whether the gut microbiota affects GI function, dysbiosis has been proposed as a pathophysiologic factor in IBS based on the concept of the gut-brain interaction. Two consecutive rifaximin trials have indicated that modulation of the gut microflora may alleviate IBS symptoms, especially for IBS-D [45]. Rifaximin, a poorly absorbable antibiotic, may contribute to inducing microbial homeostasis in the intestine, along with its major effects on the bactericidal/bacteriostatic activity and anti-inflammatory effects on the intestinal mucosa [6]. A fecal microbiota transplantation study involving IBS subjects and healthy donors, who significantly differed in the bacterial strains before transplantation, showed that the transplantation was associated with changes in gut microbiota along with improvement in the symptoms and quality of life, which lasted up to 4 weeks [7].
While understanding the pathophysiology of IBS, the role of diet cannot be overlooked because diet is an important external factor that influences the distribution and composition of intestinal microbiota. During the process of digestion and absorption, the intestinal epithelium and foods interact closely with different types and proportions of microorganisms in the GI tract. The intestinal microbiota is also responsible for maintaining intestinal homeostasis through immune regulation.
In this review, we have addressed the mechanisms by which diet affects IBS through the modulation of gut microbiota based on the hypothesis that gut microbiota imbalance is an etiology of IBS. We have also suggested a dietary strategy for the amelioration of IBS symptoms.
INTESTIANL PHYSIOLOGY UNDERLYING THE ROLE OF GUT MICROBIOTA IN IBS
The gut is a complex organ composed of many different types of cells with diverse functions, including intestinal epithelial cells such as enterocytes, goblet cells, enteroendocrine cells, Paneth cells, M cells, immune cells, smooth muscle cells, enteric neurons, and interstitial cells of Cajal [8910].
Intestinal cell functions
The various types of intestinal cells are involved in maintaining biological homeostasis (Table 1). GI tract cells play a distinctive role in food digestion, nutrient absorption and GI motility. Furthermore, GI tract cells contribute to various immune responses, including the mucosal barrier, in addition to producing hormones and neurotransmitters. Enterocytes are the major cells specialized in nutrient absorption, but also function as activators of the innate and adaptive immune system, which sense pathogens and release antimicrobial substances. M cells are antigen delivery cells, and dendritic cells are involved in phagocytosis and antigen presentation [1112]. Paneth cells secrete antibacterial mediators and growth factors, thereby maintaining the stem cell repertoire that lines the crypt walls [13]. Goblet cells, whose primary function is mucus production, line between the layers of enterocytes—a loosely adherent layer and another layer firmly attached to the mucosa. The latter layer is free of bacteria [14]. Through these multiple functions performed by the different cells, the intestinal epithelium maintains the structural integrity of the gut mucosal barrier against the penetration of microbes and enhances immune function [15]. GI motility is a consequence of the coordination of neural and smooth muscle cells and interstitial cells of Cajal [16]. Enteroendocrine cells also contribute to motor function by secreting hormones [17]. Thus, disturbances in normal intestinal cell functions may be associated with the pathogenesis of IBS, including impaired bowel motility, persistent low-grade mucosal inflammation and visceral hypersensitivity [1418].
Table 1
Intestinal cell functions involved in IBS pathophysiology
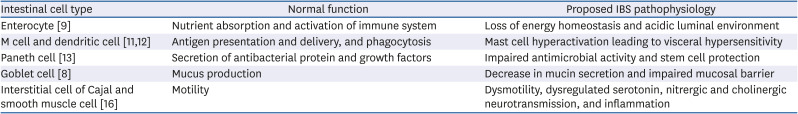
| Intestinal cell type | Normal function | Proposed IBS pathophysiology |
|---|---|---|
| Enterocyte [9] | Nutrient absorption and activation of immune system | Loss of energy homeostasis and acidic luminal environment |
| M cell and dendritic cell [1112] | Antigen presentation and delivery, and phagocytosis | Mast cell hyperactivation leading to visceral hypersensitivity |
| Paneth cell [13] | Secretion of antibacterial protein and growth factors | Impaired antimicrobial activity and stem cell protection |
| Goblet cell [8] | Mucus production | Decrease in mucin secretion and impaired mucosal barrier |
| Interstitial cell of Cajal and smooth muscle cell [16] | Motility | Dysmotility, dysregulated serotonin, nitrergic and cholinergic neurotransmission, and inflammation |
Gut microbiota and IBS
The human GI tract harbors more than 100 trillion microorganisms and the large intestine carries approximately 1011 microbial cells/mL [19]. It is estimated that 150 to 400 species dwell in human intestine. Gut microbiota are composed of different bacteria species taxonomically classified by species, genus, family, order, class and phylum. Dominant phyla in the healthy adult population are Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, Fusobacteria, and Verrucomicrobia [2021]. The 2 phyla Firmicutes and Bacteroidetes represent 90% of gut microbiota. The Firmicutes phylum consists of genera such as Lactobacillus, Bacillus, Clostridium, Enterococcus, and Ruminococcus. The Bacteroidetes is composed of genera such as Bacteroides and Prevotella. The Actinobacteria phylum, which is relatively less abundant, is mainly represented by the Bifidobacterium genus [22].
Metagenomic studies have suggested an increased ratio of Firmicutes to Bacteroidetes in patients with IBS [23]. The gut microbiota continues to change throughout the span of human life, and the composition and distribution of microbial species are affected by various external and internal factors (Table 2) (Fig. 1) [242526272829]. Although the individual diversity and richness of the intestinal bacterial phylotypes are different, they form functionally similar intestinal flora. It has been understood that these stable and balanced clusters of microorganisms are maintained in the GI tract and they competitively defend against the invasion of various pathogens.
Table 2
Factors that alter the gut microbiota profile
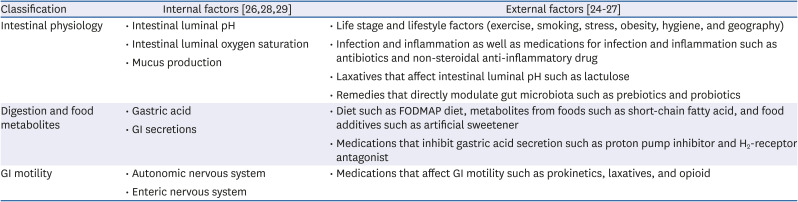
| Classification | Internal factors [262829] | External factors [24252627] |
|---|---|---|
| Intestinal physiology | • Intestinal luminal pH | • Life stage and lifestyle factors (exercise, smoking, stress, obesity, hygiene, and geography) |
| • Intestinal luminal oxygen saturation | • Infection and inflammation as well as medications for infection and inflammation such as antibiotics and non-steroidal anti-inflammatory drug | |
| • Mucus production | • Laxatives that affect intestinal luminal pH such as lactulose | |
| • Remedies that directly modulate gut microbiota such as prebiotics and probiotics | ||
| Digestion and food metabolites | • Gastric acid | • Diet such as FODMAP diet, metabolites from foods such as short-chain fatty acid, and food additives such as artificial sweetener |
| • GI secretions | • Medications that inhibit gastric acid secretion such as proton pump inhibitor and H2-receptor antagonist | |
| GI motility | • Autonomic nervous system | • Medications that affect GI motility such as prokinetics, laxatives, and opioid |
| • Enteric nervous system |
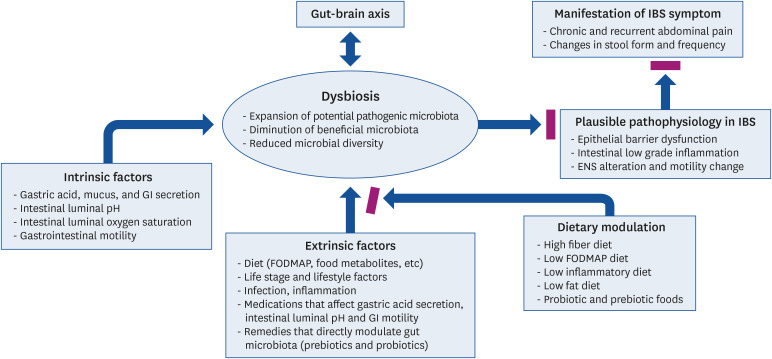
Fig. 1
Interrelation between IBS and dysbiosis of the intestinal microbiota. Many extrinsic and intrinsic factors can influence the dysbiosis of intestinal microbiota, which has been suggested as a novel pathophysiologic mechanism of IBS. A hypothesis is proposed that dietary modulation may improve IBS symptoms by mitigating the dysbiosis.
ENS, enteric nervous system; FODMAP, fermentable oligosaccharides, disaccharides, monosaccharides, and polyols; GI, gastrointestinal; IBS, irritable bowel syndrome.
GUT-BRAIN AXIS (GBA) AND IBS
Since the fact that brain can disturb intestinal function, IBS symptoms have been considered rather as a psychologic manifestation [30]. Over the past decade, accumulating evidence has indicated that dysregulation of the GBA, in which the microbiota plays a critical role, has been highlighted in the pathogenesis of IBS. The notion of the GBA is that there exists a crosstalk between the brain and the gut, thereby implying that the crosstalk shares the ability to affect their individual functioning [31]. GBA is a bidirectional network which includes the central nervous system, the autonomic nervous system, the enteric nervous system, and the hypothalamic pituitary adrenal axis. It is mainly involved in the regulation of GI motility, secretion of digestive enzymes, and communication with other organs using various neurotransmitters such as acetylcholine, dopamine, and serotonin. Among the various systems involved, the enteric nervous system is the main network that not only regulates gut motility and secretion, but also is involved in the immune system and maintenance of intestinal homeostasis [32]. Thus, proper functioning of the GBA is critical for the physiology of the digestive system, and the dysregulation of the GBA through interactions with gut microbiota has been suggested as an etiology of IBS. In fact, gut microbiota and its metabolites can affect GI motility, its alteration being a hallmark of IBS, by influencing pathways involving enteric neurons, neuroglia, or enteric muscular macrophages [33]. Thus, dysbiosis, although reported only in a portion of the patients with IBS, might be a candidate causal mechanism underlying the altered GI motility in IBS.
The tryptophan pathway has been highlighted as a metabolic pathway for microbiota-associated GBA. In these metabolic pathways, molecules such as serotonin, kynurenine, and indole derivatives might affect the gut microbiota via the GBA [34353637]. Tryptophan is an essential amino acid obtained from the diet and is an important precursor of serotonin (5-hydroxytryptamine [5-HT]). Although plasma tryptophan concentrations may not differ between patients with IBS and healthy controls [38], it has been hypothesized that tryptophan is a key amino acid in IBS, because of the relative importance of 5-HT in functional GI disorders [38]. Alterations in 5-HT homeostasis have been reported in IBS patients, which might be associated with dysmotility because the 5-HT mucosal content in IBS patients is lower than that in healthy individuals [39]. The depletion of host tryptophan resulting from indoleamine-2,3-dioxygenase activation or dietary restriction reduced the proliferation of Lactobacillus [40], some of which are reported to be tryptophan-utilizing bacteria [344142]. Restoration of tryptophan levels selectively induced the expansion of Lactobacillus [40]. It appears that impaired tryptophan metabolism, which can modulate the GBA, may potentially participate in the manifestation of IBS symptoms associated with both local and central nervous system.
DYSBIOSIS IN IBS
Dysbiosis refers to gut microbial imbalance in their composition and function, which are characterized by one or more of 3 types; expansion of potential pathogenic microbes in the gut microbiota community, diminution of beneficial microbiota, and/or decrease in microbial diversity [4344] (Fig. 1).
In terms of beneficial bacteria, the composition and activities of lactobacilli and Bifidobacterium are well known to be compromised in IBS [45]. In patients with severe IBS, Prevotella and Methanobacteriales such as Methanobrevibacter smithii and Methanosphaera stadtmanae were reduced with increasing severity of symptoms in parallel with increased Bacteroides [46]. In this study, dietary factors did not affect the gut microbial signature in severe IBS cases. Although it is still not clear whether IBS causes microbial changes or vice versa [47], dysbiosis may disrupt the epithelial barrier function and elicit low-grade inflammation, which can be a cause of visceral pain. Dysbiosis is also expected to activate the cellular and mucosal immune system, possibly resulting in low-grade inflammation and a leaky gut. Furthermore, alterations in the enteric nervous system and changes in gut motility due to dysbiosis are also anticipated to cause visceral hypersensitivity and bowel habit changes [48]. Regarding the expansion of potential pathogenic microbes, the family Enterobacteriaceae (phylum Proteobacteria), family Lactobacillaceae and genus Bacteroides (phylum Bacteroidetes) [49] were demonstrated to be present in IBS. In addition, Enterobacteriaceae-containing pathogenic bacteria with a corresponding reduction in Lactobacillus can aggravate IBS symptoms by producing organic acids [50], and Bacteroides can worsen the symptoms, especially in IBS-D patients by inducing low-grade mucosal inflammation [5152]. Thus, one might speculate that a new strategy—expansion of beneficial microbiota such as lactobacilli and bifidobacteria and reduction of those bacteria considered as harmful (Clostridium, Escherichia coli, Salmonella, Shigella and Pseudomonas), may alleviate IBS symptoms [53]. Lastly, microbial diversity and distribution according to dietary patterns have been regarded to play a crucial role in IBS pathogenesis by modulating the gut microenvironment, including colonic fermentation, gut microbiome composition and gut immune system [545556]. In fact, diet can affect motility, permeability, visceral sensation, gut-brain interactions, immune regulation, and neuroendocrine function by interacting with the gut epithelium and microbiota [57]. The dynamic and flexible components of the gut microbiota can be influenced by diet, thereby affecting the IBS manifestation [5859].
DIETARY MODULATION FOR THE RELIF OF IBS SYMPTOMS
It is known that the intestinal microbiota plays a role in the further degradation of food metabolites that are already digested by the host, and this may affect GI epithelial cells. Thus, human host cells and microbiota, which exist in approximately equal proportions, communicate with each other to regulate metabolic homeostasis and the immune function [6061]. Interestingly, the gut microbiota also uses dietary nutrients for their survival and growth. The ingested foods and their metabolites can alter the normal gut microbiota profile and colonic fermentation, resulting in the dysregulation of gut permeability, motility and visceral hypersensitivity in IBS [56].
It has been postulated that the dietary modulation of gut microbiota can be one of the strategies to relieve IBS symptoms, especially because diet is consumed every day and habitual diet modification has a cost-effective advantage over expensive drug therapy. In general, dietary intervention in IBS includes regular meals, reducing the intake of IBS causable foods and a high-fiber diet [62]. The effect of increasing the beneficial and reducing the harmful microbiota through dietary modification has been reproduced in several studies [6364656667]. Thus, a diet that limits the intake of “offending” foods that trigger these alterations, or a diet that corrects the microbiota dysbiosis, would be useful to manage IBS symptoms (Table 3). These dietary benefits have been largely attributed to reduction in the levels of colonic fermentation or decreased antigen activation of the gut immune system. It is important to consider that diet significantly affects the composition of the gut microbiome [5255]. Additional prebiotics, which are food compounds that can feed the microbiota such as fructo-oligosaccharides and galacto-oligosaccharides, and/or probiotics, which are useful live microorganisms such as Lactobacillus and Bifidobacterium, may provide further beneficial effects to induce microbial homeostasis and improve symptoms [68].
Table 3
Dietary modulation of gut microbiota for IBS symptom relief

| Diets | Microbiota changes | IBS symptoms | Ref. |
|---|---|---|---|
| High fiber diet | Fibers are the main source of microbiota-accessible carbohydrates that increases the gut microbiota diversity and produces beneficial metabolites such as short-chain fatty acid. | Soluble fibers relive symptoms by softening the stool. High fiber diet has been a standard care particularly for IBS-C. | [76] |
| Low FODMAP diet | Actinobacteria, Bifidobacterium, and Faecalibacterium prausnitzii were decreased by low FODMAP diet. | Improves overall IBS symptoms such as visceral pain, bloating and quality of life. | [90] |
| Low inflammatory diet | Bacteroidetes phylum was more abundant in vegans and vegetarians. | Alleviates chronic low-grade mucosal inflammation, which helps enterocyte healing and nutritional support to the microbial environment. | [102128] |
| Low fat diet | Increases bacterial diversity and beneficial butyrate-producing bacteria. | Beneficial effects on intestinal inflammation. | [133] |
| Nuts and other plant-based fat increases the genera Bifidobacterium, Roseburia, and Faecilibacterium. |
High fiber diet
Fibers are the main source of microbiota-accessible carbohydrates in the human diet, and fiber supplementation can increase the diversity of gut microbiota and beneficial metabolites. Dietary fibers, a type of low-absorbed carbohydrate, are naturally present in foods such as fruits, vegetables, whole grains, and legumes [69], and are fermented by enteric bacteria in the colon to produce short-chain fatty acids (SCFAs) and gases [70]. The primary SCFAs produced in the gut are acetate, propionate, and butyrate [71]. SCFAs and other metabolites are thought to mediate beneficial effects, including the preservation of gut epithelial integrity, energy homeostasis, and inflammatory signal inhibition and immunity [72]. Unfortunately, to date, it is not yet clear whether SCFAs and their metabolites may conduct the beneficial effects on IBS through the modulation of gut microbiota environment [7374].
IBS is believed to be caused by a deficient intake of dietary fiber, and most physicians recommend that patients with IBS increase their intake of dietary fiber to relieve their symptoms. However, different types of dietary fiber have different physical and chemical properties.
Dietary fibers are divided into soluble and insoluble fibers. Soluble fibers, which slow digestion, are rich in oatmeal, nuts, seeds, beans, lentils, peas, and some fruits such as apples, blueberries, and vegetables. Psyllium, a common soluble fiber supplement, is known to be effective for relieving symptoms in IBS [75]. Insoluble fiber is found in foods, including wheat bran, vegetables, and whole grains but is not directly effective for relieving symptoms. Although insoluble fibers are generally poorly fermented by gut microbes, their presence in the diet increases the gut transit rate and thus reduces the time for colonic bacterial fermentation of non-digested foods [64]. Although debatable, the standard care for IBS has been a high-fiber diet, particularly in IBS-C patients, given the absence of serious side effects and its potential benefit on gut microbiota [76]. In IBS-C, fiber also has the effect of softening the stool, although it does not alleviate the pain. In a clinical setting, fiber supplements are hence recommended to begin at a low dose and increase slowly because fiber can aggravate IBS symptoms such as bloating, gas, and pain.
Low fermentable oligosaccharides, disaccharides, monosaccharides, and polyols (FODMAP) diet
FODMAP are short-chain carbohydrates that are resistant to digestion and can cause digestive problems, especially in IBS patients. FODMAPs move slowly in the small intestine, thereby increasing the water content due to osmotic pressure causing diarrhea. In the colon, fermentation by colonic bacteria may lead to gas production and water secretion, precipitating luminal distension and stimulating abnormal motility [50]. Fermented metabolites from FODMAP may include SCFAs such as acetate, propionate, and butyrate and variable amounts of hydrogen and carbon dioxide [77]. These metabolites may alter the colonic microbial environment and affect the IBS symptoms. For example, increased production of SCFAs may increase luminal acidity. In double-blind placebo-controlled trials, FODMAPs exacerbated IBS symptoms such as abdominal pain, bloating, increased flatulence, and changes in bowel habits [7879]. Fructo-oligosaccharides primarily contribute to colonic distention via microbial fermentation and subsequent gas production [70]. Furthermore, FODMAP-induced symptoms have been suggested to be associated with baseline visceral hypersensitivity in patients with IBS [808182].
There has been growing insight into the beneficial effects of a low FODMAP diet in the management of IBS symptoms. Several randomized controlled trials have demonstrated the efficacy of a reduced high FODMAP food intake in improving overall IBS symptoms such as visceral pain, bloating, and quality of life [7983848586878889] (Table 4). A recent review of low FODMAP dietary therapy reported that at least 50% of patients with IBS experienced symptomatic benefits [87]. In a 9-week clinical trial, a low FODMAP diet not only alleviated IBS symptoms compared to maltodextrin (placebo) and FODMAP diet but also decreased serum levels of interleukin (IL)-6, IL-8, total SCFAs, n-butyric acid, and fecal bacteria (Actinobacteria, Bifidobacterium, and Faecalibacterium prausnitzii) compared to the baseline levels, suggesting that a low FODMAP can induce changes in levels of inflammatory cytokines, SCFA and gut microbiota profile [90].
Table 4
Clinical trials investigating the effect of the low FODMAP diet on the microbiota in IBS
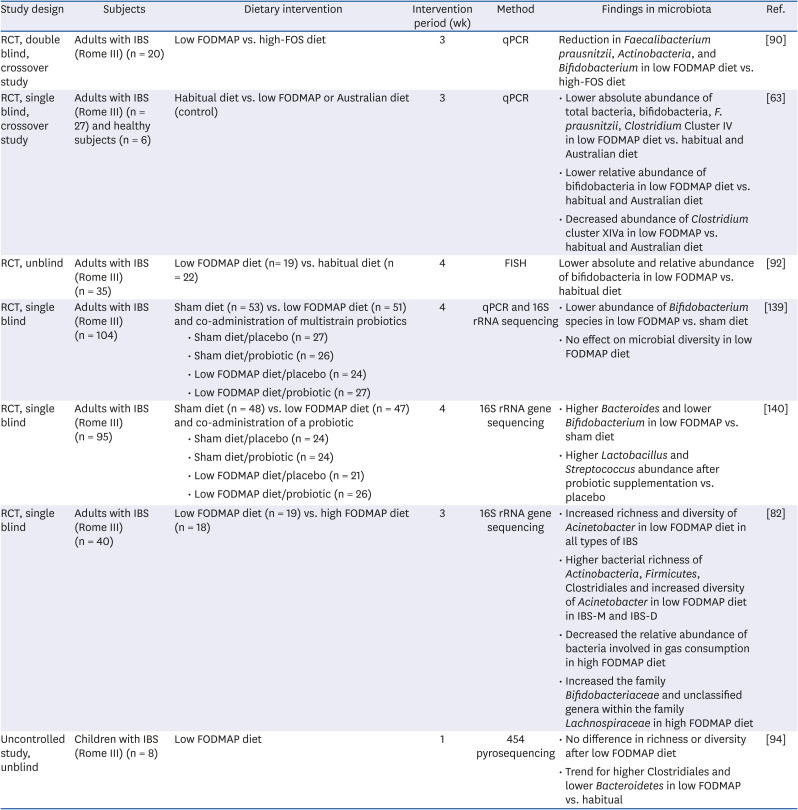
| Study design | Subjects | Dietary intervention | Intervention period (wk) | Method | Findings in microbiota | Ref. | |
|---|---|---|---|---|---|---|---|
| RCT, double blind, crossover study | Adults with IBS (Rome III) (n = 20) | Low FODMAP vs. high-FOS diet | 3 | qPCR | Reduction in Faecalibacterium prausnitzii, Actinobacteria, and Bifidobacterium in low FODMAP diet vs. high-FOS diet | [90] | |
| RCT, single blind, crossover study | Adults with IBS (Rome III) (n = 27) and healthy subjects (n = 6) | Habitual diet vs. low FODMAP or Australian diet (control) | 3 | qPCR | • Lower absolute abundance of total bacteria, bifidobacteria, F. prausnitzii, Clostridium Cluster IV in low FODMAP diet vs. habitual and Australian diet | [63] | |
| • Lower relative abundance of bifidobacteria in low FODMAP diet vs. habitual and Australian diet | |||||||
| • Decreased abundance of Clostridium cluster XIVa in low FODMAP vs. habitual and Australian diet | |||||||
| RCT, unblind | Adults with IBS (Rome III) (n = 35) | Low FODMAP diet (n= 19) vs. habitual diet (n = 22) | 4 | FISH | Lower absolute and relative abundance of bifidobacteria in low FODMAP vs. habitual diet | [92] | |
| RCT, single blind | Adults with IBS (Rome III) (n = 104) | Sham diet (n = 53) vs. low FODMAP diet (n = 51) and co-administration of multistrain probiotics | 4 | qPCR and 16S rRNA sequencing | • Lower abundance of Bifidobacterium species in low FODMAP vs. sham diet | [139] | |
| • Sham diet/placebo (n = 27) | • No effect on microbial diversity in low FODMAP diet | ||||||
| • Sham diet/probiotic (n = 26) | |||||||
| • Low FODMAP diet/placebo (n = 24) | |||||||
| • Low FODMAP diet/probiotic (n = 27) | |||||||
| RCT, single blind | Adults with IBS (Rome III) (n = 95) | Sham diet (n = 48) vs. low FODMAP diet (n = 47) and co-administration of a probiotic | 4 | 16S rRNA gene sequencing | • Higher Bacteroides and lower Bifidobacterium in low FODMAP vs. sham diet | [140] | |
| • Sham diet/placebo (n = 24) | • Higher Lactobacillus and Streptococcus abundance after probiotic supplementation vs. placebo | ||||||
| • Sham diet/probiotic (n = 24) | |||||||
| • Low FODMAP diet/placebo (n = 21) | |||||||
| • Low FODMAP diet/probiotic (n = 26) | |||||||
| RCT, single blind | Adults with IBS (Rome III) (n = 40) | Low FODMAP diet (n = 19) vs. high FODMAP diet (n = 18) | 3 | 16S rRNA gene sequencing | • Increased richness and diversity of Acinetobacter in low FODMAP diet in all types of IBS | [82] | |
| • Higher bacterial richness of Actinobacteria, Firmicutes, Clostridiales and increased diversity of Acinetobacter in low FODMAP diet in IBS-M and IBS-D | |||||||
| • Decreased the relative abundance of bacteria involved in gas consumption in high FODMAP diet | |||||||
| • Increased the family Bifidobacteriaceae and unclassified genera within the family Lachnospiraceae in high FODMAP diet | |||||||
| Uncontrolled study, unblind | Children with IBS (Rome III) (n = 8) | Low FODMAP diet | 1 | 454 pyrosequencing | • No difference in richness or diversity after low FODMAP diet | [94] | |
| • Trend for higher Clostridiales and lower Bacteroidetes in low FODMAP vs. habitual | |||||||
RCT, Randomized controlled trials; IBS, irritable bowel syndrome; FODMAP, fermentable oligosaccharides, disaccharides, monosaccharides, and polyols; FOS, fructo-oligosaccharides; qPCR, quantitative polymerase chain reaction; FISH, fluorescence in situ hybridization; IBS-M, irritable bowel syndrome-mixed type of diarrhea and constipation; IBS-D, irritable bowel syndrome-diarrhea-predominant.
A low FODMAP diet may have an adverse effect on the gut microbiota because low FODMAPs have reduced levels of dietary prebiotics, such as fructo-oligosaccharides, galacto-oligosaccharides, and fibers, which may reduce beneficial intestinal microbiota and decrease the production of SCFAs that have important protective and trophic activity on the colonocytes [91]. Some studies have shown that a low FODMAP diet reduced Bifidobactrium [9293], which has been regarded as a worse symptom profile, suggesting the supplementation of bifidobacteria-containing probiotics, which has been reported to reduce IBS symptoms, to the low FODMAP diet [93]. In a human study that compared the microbial effects between low and high FODMAP diets, low FODMAP diet increased the richness and diversity of Actinobacteria, unlike the studies that showed a reduction in bifidobacteria with a low FODMAP diet, and a high FODMAP diet decreased the relative abundance of bacteria involved in gas consumption [82]. A series of studies investigating the effect of low FODMAP on gut microbiota and its metabolites have shown mixed and sometimes conflicting results in contrast to the symptomatic improvement by low FODMAP [6394]. The conflicting results are probably due to the study design including different subjects, study durations and/or different control diets, heterogeneity of low FODMAP diet, and methods to measure gut microbiota. It appears that the effect of low FODMAP through low luminal osmolality, reduced gas production, and GI motility may overcome the adverse microbial effects or the microbial changes caused by the low FODMAP diet, hence having minimal or no effect on IBS symptoms. However, a simple analysis of feces cannot reliably reveal the actual GI tract picture. Other approaches, such as metagenomics, transcriptomics, and proteomics, can provide information that is more reliable. If a low FODMAP diet does adversely affects the gut microbiota profile in IBS and is associated with worsening IBS, we need to determine a way to deal with these adverse effects, such as supplementation with probiotics or modulation of ingredient composition of the low FODMAP diet. In an interesting study using metagenomic sequencing in children with IBS [95]; at the baseline non-responders were enriched with the genus Turicibacter from the family Turicibacteraceae. This suggests that the profile of microbiome before starting a low FODMAP diet can be useful to predict responders and non-responders.
Collectively, the low FODMAP diet seems to change the gut microbiota profile in the direction against symptomatic relief or in other words, not imposing a beneficial effect. The long-term beneficial effects of a low FODMAP diet on the interaction between the gut epithelium and microbiota has not been established [96]. Further studies focusing on the long-term effects of a low FODMAP diet on the clinical outcomes and dynamics of gut microbiota composition in patients with IBS are warranted.
Low inflammatory diet
Low-grade mucosal inflammation has been proposed as an important component of the IBS pathophysiology. Even in normal mucosa, the interaction between host immune cells and commensal microorganisms may induce inflammation, but it is a well-controlled and sustained low-grade subclinical inflammation. In pathologic conditions, such as IBS, increased mast cells and activated T lymphocytes were found in the colonic mucosa of IBS patients compared with normal control subjects [979899]. In severe IBS, lymphocytic infiltration in the myenteric plexus and inflammatory cell intrusion into the muscularis externa was shown to even lead to motor dysfunction of the GI tract [100]. Individuals with IBS have been shown to have a high incidence of low-grade systemic inflammation [101]. Thus, alleviating chronic, low-grade mucosal inflammation through diet, especially with an anti-inflammatory diet, may be an effective therapeutic strategy through enterocyte healing and nutritional support to the microbial environment [102].
The Dietary Inflammatory Index (DII) was developed to estimate the efficacy of a diet on chronic sustained inflammation (meta-inflammation) and the risk of inflammation-mediated diseases. DII determines the inflammatory potential from a maximum anti-inflammatory diet to a maximum pro-inflammatory diet by calculating the pro- and anti-inflammatory indices of each of the diet components [103]. The Western diet, which is high in saturated fat and simple carbohydrates, is a representative pro-inflammatory diet and known to be associated with the increased risk of IBS [104105], whereas the Mediterranean diet (MD), which is high in high-fiber whole grains, nuts, legumes, fruits, vegetables and fish, has been regarded as a low inflammatory diet [106107108]. In MD, the inclusion of saturated fat is limited, and the majority in the dietary fat content is monounsaturated fats (olive oil) and omega-3 polyunsaturated fatty acids (fish). Extra virgin olive oil, a major constituent of the MD and enriched with oleic acid and polyphenols, has a major beneficial role in exerting antioxidant activity, thereby positively affecting several biomarkers of oxidative stress [109110111112113]. These polyphenols are directly absorbed or metabolized in the intestine and transformed into active metabolites, where they may bring about a significant local effect through their interaction with the gut microbiota, which in-turn increases the levels of Bacteroidetes and reduces the Firmicutes/Bacteroidetes ratio [114]. They potentially modulate the oxidative status of the intestinal barrier, inflammation, and immune response of the host [113]. High nut consumption is a characteristic of MD. Walnuts, almonds, and many other nuts have been demonstrated to have a protective role against inflammation [115], suggesting that nuts are a component causing the anti-inflammatory effect of MD [116]. Furthermore, the daily intake of walnuts significantly affects the gut microbiome by increasing probiotic- and butyric acid-producing species [117]. Although few studies have suggested symptomatic improvement of IBS by MD [118], it could be worthwhile to study the effects of MD on IBS due to its anti-inflammatory effects and beneficial microbial effects.
A diet enriched with nutrients that are known to have anti-inflammatory properties such as vitamins C, D, and E, beta-carotene, polyunsaturated fatty acids, flavonoids, and dietary fiber can be a low inflammatory diet [119]. Interestingly, the host blood vitamin levels are associated with IBS symptoms. The level of serum vitamin D, which is known to be associated with immune function and inflammation, tended to be lower in patients with IBS [120]. Moreover, low vitamin D status correlated with severe symptoms in patients with IBS. In a recent report, the severity of symptoms significantly decreased after vitamin D3 supplementation in IBS-D patients with low vitamin D status as compared to the control group [121122123124]. It is well known that low vitamin B-6 status is linked to inflammatory conditions, and interestingly, low intake of vitamin B-6 was correlated with a high IBS symptom score [125]. However, there is no evidence that supplementation with these vitamins can actually improve IBS symptoms.
A recent report proposed a food-based DII associated with IBS and suggested that a diet with a low DII could be helpful for IBS patients [126]. Another report showed that a more pro-inflammatory diet was associated with certain microbes, such as Ruminococcus torques, Eubacterium nodatum and Acidaminococcus intestine [127]. In a recent report, the Bacteroidetes phylum was more abundant in vegans and vegetarians than in omnivores, and high consumption of vegetables consistent with an MD is more associated with beneficial microbiome-related metabolomic profiles compared to individuals consuming a Western diet [128]. Further randomized controlled trials may provide more insight into the beneficial effect of a low inflammatory diet on the gut microbiota and subsequent improvement of IBS symptoms.
Low fat diet
A certain proportion of IBS patients can develop symptoms in response to the consumption of foods rich in fat and the avoidance of fatty meals is one of the most frequent dietary recommendations for improving IBS symptoms [129].
Dietary fat can modify the gut microbiota, and the host lipid metabolism can be affected by the gut microbiota through the regulation of fat absorption, storage, and energy harvesting from the diet [130]. Compared to low-fat diets, high-fat diets have frequently been shown to increase the abundance of intestinal microbiota [131132]. The different effects of plant- and animal-based fat sources on the gut microbiota have been examined. In a recent report, nuts and other plant-based fat sources such as avocados and olive oil increased the genera Bifidobacterium, Roseburia, and Faecilibacterium, all of which are reported to have beneficial metabolic effects [133]. Similarly, different effects of saturated and unsaturated fats on intestinal microbiota have also been reported. In a mouse model, a diet enriched with coconut oil (high in saturated fat) induced a relatively increased abundance of Allobaculum, Anaerofustis, F16, Lactobacillus reuteri, and Deltaproteobacteria and a relatively lower abundance of Akkermansia muciniphila compared to diets enriched with soy oil (high in polyunsaturated fat) in the cecum bacterial community [134]. Functionally, the genes of cecal microbiota of high-coconut oil-fed mice were predicted to deplete the pathways involved in fatty acid and amino acid metabolism. Polyunsaturated fatty acids and their metabolites can exhibit beneficial effects on intestinal inflammation, which has been suggested as a mechanism involved in IBS. Another study demonstrated that a high saturated fat diet increased the gram-negative intestinal microbiota and subsequently increased endotoxemia [135]. Metabolic endotoxemia through the gut microbiota composition and altered intestinal permeability, along with a disturbed membrane barrier, may contribute to mucosal inflammation by saturated fat [136]. Collectively, fats from plant-based sources such as nuts and dietary patterns rich in polyunsaturated and monounsaturated fats such as soy oil and olive oil have been associated with increased bacterial diversity and beneficial butyrate-producing bacteria, and hence, a positive metabolic influence [133].
Probiotic and prebiotic foods
IBS is associated with dysbiosis, which is characterized by the loss of diversity and changes in the composition of beneficial and pathogenic microbiota. Thereafter, theoretically, supplementing the diet with beneficial bacteria can be helpful for the treatment and improvement of the IBS symptoms. However, the role of the effectiveness of probiotic supplementation in IBS patients has not been clarified because most of the studies so far have been small-sized and short-term studies [137]. Nevertheless, probiotics have been used to improve the overall abdominal symptoms and bloating.
Prebiotics are defined as compounds in food that induce the growth or activity of beneficial microorganisms such as Bifidobacterium, which show anti-inflammatory effects in the intestines. Prebiotic foods include asparagus, leek, banana, garlic, barley, roots, flaxseeds, wheat bran, apple, cocoa and whole wheat flour. However, there is insufficient evidence for the beneficial effects of prebiotics in patients with IBS [137]. Although prebiotics usually do not improve GI symptoms in patients with IBS, the severity of flatulence was improved by prebiotics at doses ≤ 6 g/d, while inulin-type fructo-oligosaccharides worsened flatulence, suggesting that variations in the type and dose of the prebiotics in the diet may affect the improvement or exacerbation of the symptom [138].
CONCLUSIONS
Although the etiology of IBS remains unclear, alterations in gut microbiota have been postulated as an etiologic factor in the development of IBS and IBS symptoms. This takes place due to human intestinal microbiota coexisting with the host intestinal cells in a symbiotic relationship, participating in metabolism, and maintaining homeostasis. Even though we do not know the exact healthy intestinal microbial environment to date, modulation of a daily diet that is currently known to improve the microbial environment might be the most physiologic and effective strategy in the treatment of IBS.
Thus, in addition to the empirical dietary treatment that limits the foods that cause the IBS symptoms and retains foods that improve symptoms, dietary modulation of the microbiota profile, including a high-fiber diet, the effects of which have traditionally been recognized without significant side effects, could also be useful. Low FODMAP diets, low inflammatory diets including MD, low fat diet and pre- and probiotic foods have been attempted to improve IBS symptoms. It is now necessary to determine whether these kinds of diets that aim to improve microbial balance are safe and effective in patients with IBS. Well-designed large population studies will provide further insight into dietary modulation for the relief of IBS symptoms. Moreover, the possible long-term nutritional consequences that can occur when certain foods are restricted or maintained should also be investigated so that patients with IBS do not suffer from the deficiency of essential nutrients.
 XML Download
XML Download