

 PDF
PDF Citation
Citation Print
Print



서 론
인체 내 전체 미생물의 95% 정도는 대장을 포함한 소화기관에 주로 존재하고, 그 외 1%가 호흡기, 생식기, 구강, 피부등에도 널리 분포하기 때문에 여러 학자들의 연구가 장내세균에 집중되고 있다. 인체 공생 미생물의 대다수를 차지하는 장내미생물(gut microbiota) 은 출생 후 면역기관과 내장신경총 발달에 기여하고 지속적으로 중요한 면역작용에 관여하며, 약물에 대한 반응을 조절하고, 신진대사에 큰 영향을 주는 등 신체대사활동에 중요한 역할을 한다. 특히 2000 년대 이후에 차세대 염기서열 분석기법(next-generation sequencing) 이 보편화되면서 메타게노믹스(metagenomics) 연구가 매우 활발해졌는데, 이러한 분석법에 의하면 식이, 인종, 나이, 항생제, 스트레스, 심리적 요인, 산모의 건강, 자연분만 등의 출생 방법, 환경 요인과 운동 등이 장내미생물 분포에 영향을 주는 것으로 알려지고 있다.1 남녀 차이가 나타나기 시작하는 사춘기, 임신주기별 차이, 폐경기 그리고 노령 등 연령의 차이를 결정하는 것이 결국 성호르몬이고 성호르몬의 대표격인 에스트로젠(estrogen)과 테스토스테론(testosterone) 은 장내미생물과 면역세포에 직접적인 영향을 미쳐 위장관계 여러 질환에 복잡한 영향을 미칠 것으로 추측되지만 장내미생물 연구에 있어 성차의 중요성이 인식되기 시작한 것은 최근의 일이다.2 이와 동시에 에스트로젠과 안드로젠(androgen) 이 장내미생물에 영향을 주고 반대로 이들 장내미생물이 에스트로젠과 안드로젠 대사에 영향을 주는 것이 알려지면서 “microgenderome” 이라는 단어가 만들어졌으나,3 아직 이에 대한 개념이 잘 알려져 있지않다. 한편 성호르몬은 유전자 수복(repair) 이나 후성학적 기전에도 영향을 주기 때문에 후성학적 변이와 이에 의한 생리학적 영향 또한 남자와 여자에서 다르게 나타난다. 이러한 상호관련성을 연구하는 성차의학(sex/gender-specific medicine) 이란 생물학적, 사회문화적 차원과 여자와 남자에 미치는 이들의 영향을 고려하는 학문이라 할 수 있겠다. 본고에서는 장내미생물에 영향을 주는 식이, 연령, 성차에 대해 알아보고 이제 연구결과가 나오고 있는 과민성 장증후군(irritable bowel syndrome), 만성 염증성 장질환(chronic inflammatory bowel disease) 과 대장암(colon cancer) 에서의 장내미생물의 성차에 대해 간략하게 알아보고자 한다.
Go to : 
본 론
1. 장내미생물에 영향을 주는 요소
장내미생물은 연령에 따라 변화하는데, 그 주된 이유는 성호르몬의 차이 그리고 연령에 따른 식이와 여러 환경요인에 영향을 받기 때문이다. 장내미생물의 역할은 인체와의 상호작용을 통해 병원균 침입을 방어하는 면역체계를 성숙시키고 비타민과 단분쇄지방산(short chain fatty acid, SCFA)을 생산하여 영양분을 공급하여 인체 대사조절에 관여한다. 그러나 항상성(homeostasis) 을 유지하고 있던 상호관계가 깨지거나 장내미생물의 불균형(dysbiosis) 이 초래되면 인체의 건강이 안 좋아지고 질병이 발생한다. 이러한 장내미생물은 장상피세포에 부착하여 생존하는 장내미생물(mucosal gut microbiota)과 장 내강에 생존하는 장내미생물(luminal microbiota)로 구분할 수 있다(Fig. 1).4 이들은 상피세포, 대식세포, 수지상세포(dendritic cell), T 림프구, B 림프구에 의한 항상성 반응을 활성화시켜 장내에서 잠재적인 병원성 세균 산물과공존할 수 있게 한다. 즉 정상적인 상태에서 장내미생물의 역할은 병원성의 선천면역(innate immunity) 및 적응면역(adaptive immunity) 반응을 조절하고, 장 점막의 면역체계를 성장시키는 데 중요한 역할을 하지만 유익균과 유해균의 균형이 깨지는 순간 우리 몸의 건강을 해치는 방향으로 진행할 수 있는 것이다.
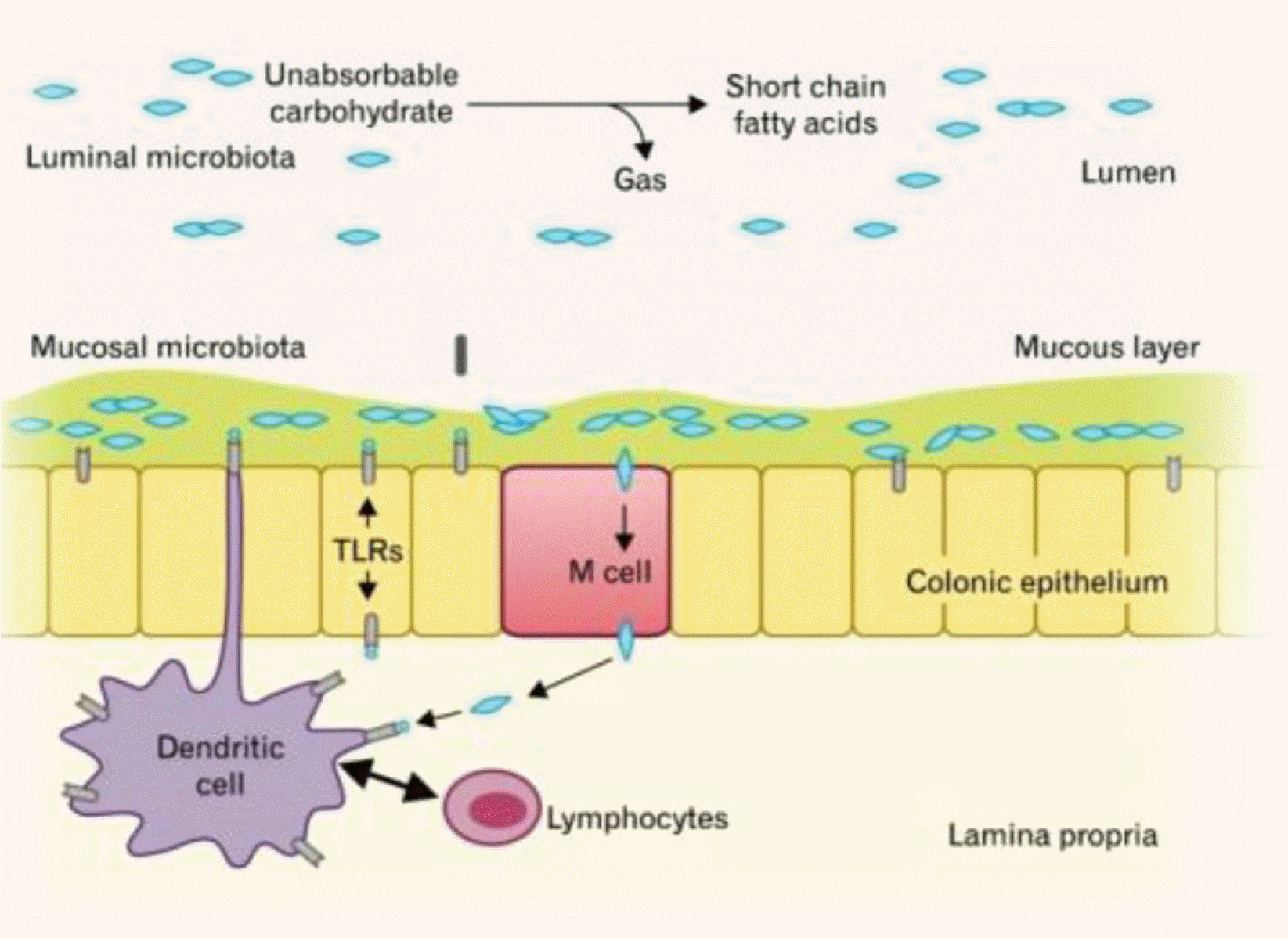 | Fig. 1Luminal and mucosal colonic microbiota and their roles in gut homeostasis. TLR, toll-like receptor. Adapted from Seo et al.4 with permission.
|
1) 식이에 따른 장내미생물 변화
식이는 장내미생물 조성에 관여하여 인체의 건강에 영향을 주는데, 98명의 분변을 채취분석한 연구에 의하면, 분변 내 미생물은 주로 Bacteroides 와 Prevotella 가 우세한 군집을 이루고 있다. 장기간의 식이에 의해 단백질이나 동물성 지방이 풍부한 음식을 섭취하는 경우에는 Bacteroides 가, 탄수화물이 풍부한 음식을 먹은 경우에는 Prevotella 가 증가하였다.5 이러한 식이에 따른 장내미생물의 변화는 어린 시절부터 나타나는데, 이탈리아 도시와 아프리카 농촌의 어린이들을 비교한 연구에서 14명의 건강한 아프리카 어린이에서는 섬유소와 다당류를 가수분해하는 Prevotella와 Xylanibacter 가 풍부하여섬유소로부터 에너지를 얻고, 생물다양성(biodiversity) 이 풍부하였다.5 또한 15명의 이탈리아 도시 어린이보다 SCFA 와대장암 예방 효과가 있는 뷰티르산(butyrate) 의 농도가 높았으며, E. coli, Shigella, Salmonella가 적게 발견되었다.5 아쉽게도 이 연구에서는 남녀 어린이로 구분하여 보고하지 않아 성차에 대한 결과는 알 수 없지만 이들 어린이들은 사춘기 전이고 그 숫자가 작아 차이가 나기는 어려웠을 것으로 추측된다.
한편 식이에 대한 장내미생물에 대한 연구의 대표적인 것이 고지방식이에 대한 변화이다. F344 백서에서 전체 칼로리의 60% 이상을 지방이 차지하는 고지방식이를 8주간 섭취하게 한 결과 인체 나이 6세에 해당하는 6주령 수컷 쥐, 암컷쥐와 인체 나이 80세 이상에 해당하는 2년령 암컷 쥐에서는 고지방을 해결하는 장내세균의 재편성이 잘 되었으나 2년령수컷 쥐에서는 이러한 적응이 나타나지 않는 성차가 있었다.6 이러한 식이가 인체 장내미생물에 영향을 주려면 6개월 정도 기간이 필요하다고 추정되었으나 최근 마우스에서의 보고를 보면 고지방식이에 대해 단 하루 만에도 차이가 나타나, 7 식이가 장내미생물에 미치는 영향은 아주 빠름을 시사하였다. 여자가 남자보다 과일, 채소를 선호한다는 국내외 보고8,9를 감안해 볼 때 남녀에 따른 장내미생물의 차이는 성호르몬의 영향과 함께 남녀의 식이 차이를 그 원인 중 하나로 생각할 수있겠다.
2) 연령에 따른 장내미생물 변화
출생 방법에 따라 장내미생물의 성차를 보이다가 보통 소아기에서는 사라지는데, 사춘기 전에는 성호르몬의 차이가 없기 때문으로 추정된다.10 이후 사춘기가 되면 성차가 나타나기 시작하여 노령기에 접어들 때까지 지속되는데 이는 장(intestine) 의 성숙이 호르몬 변화와 거의 비례하여 진행하고 이러한 변화는 장내미생물 변화로 나타나기 때문이다.10
(1) 출생 전·후의 장내미생물
태아가 출생하는 순간 태아의 장에 장내미생물이 정착하게 되는데 출생 후 수일 내지 수주간 Bacteroides, Bifidobacterium, Parabacteroides, Escherichia/Shigella 가 신생아 장내미생물의 대부분을 차지한다.11 한편 분만방법에 따라 신생아 장내미생물에 차이가 있는데, 자연분만으로 태어난 신생아 장에는 질에서 들어온 세균이 대부분을 차지하지만 복부절개분만(cesarean section, C-section) 으로 출생한 신생아의 경우는 산모 피부나 노출된 환경, 즉 분만에 관여한 의료진, 다른 신생아와 관련된 세균이 대부분을 차지한다.12 이러한 출생 경로에 따른 장내미생물의 차이는 소아까지 지속되어 체질량지수가 증가하거나 소아비만으로 연결된 경우가 있고,13 복부절개분만으로 태어난 신생아는 비결막염(rhinoconjunctivitis)과 같은 알레르기 질환이 많은데, 특히 여러 번의 복부절개를 통해 태어난 여자 신생아에 가장 흔하다.14,15 또한 메타분석에 의하면 복부절개분만에 의해 태어난 아이들에서 자폐증 질환이 많은데, 분만 방법에 따른 장내미생물의 차이가 그원인 중 하나로 제시되고 있다.16,17 한편 엄마의 스트레스에 대해 남자 아이가 더 큰 영향을 받는다고 알려진 바 있는데, 그기전에 대한 설명은 다음과 같다. 즉, 산모의 스트레스는질에 가장 많이 존재하는 분류균(taxon)이자 신생아 장에 가장 많은 상주균인 Lactobacillus 의 감소를 유발한다.18 이러한 Lactobacillus 감소는 모유를 분해하여 유산(lactic acid) 을 만들어 산성 장내환경(acidic gut environment) 을 만들어야 하는 중대한 시점에 차질을 가져오는데,19-21 특히 남아에서만Lactobacillus 의 감소로 인해 혐기성 세균인 Bacteroides 및 Clostridium의 증가를 보였다. 신생아에서 이러한 장내미생물조성의 변화는 뇌 발달에 중요한 영양분인 히스티딘(histidine) 이나 글루탐산염(glutamate)을 적절히 제공하지 못하는 상황을 유발할 수 있는데, 여아에서는 이러한 변화가 나타나지 않았다.22,23 이러한 여러 가지 증거들은 산모의 질내 미생물이 신생아 장내미생물에 남녀별로 영향을 주고 출생 전 스트레스가 장과 뇌 발달에 매우 중요한 성차요소임을 보여준다.
(2) 소아기 및 사춘기 시기의 장내미생물
소아 및 사춘기의 장내미생물은 구성과 기능 변화를 거치는데, 구성 측면에서 좀 더 복잡해지고 성인 장내미생물에 비해 불안정한 성향을 보인다는 보고도 있지만,24 소아기에서는 성호르몬의 발달이 없어 성차를 보이지 않는다는 보고가 많다. 이후 사춘기에는 남녀의 장내미생물의 차이가 나타나기 시작하는데,25 마우스를 이용한 연구를 보면 수컷에서는 사춘기의 변화가 성인 시기에 좀 더 성숙되지만 암컷에서는 사춘기에서의 변화가 성인까지 그대로 유지되었다.26 이에 대한 증거로 수컷 마우스를 사춘기에 거세하여 테스토스테론을 줄이면 성인이 되어도 장내미생물의 성차가 나타나지 않는데, 이는 테스토스테론이 성적 두형태(dimorphic)의 장내미생물 조성을 결정함을 시사한다.26 흥미로운 것은 성인 수컷 마우스에서 가장 다양성이 크고 숫자가 많은 맹장 내용물을 사춘기의 암컷 마우스에 주입하면 암컷의 장내미생물 및 대사체 양상이 남성화되고 암컷에서의 테스토스테론이 상승하여 성인까지 유지됨이 밝혀진 바 있다.27 이에 더 나아가 성장한 수컷 마우스의 맹장 내용물과 안드로젠 수용체 길항제인 flutamide 를 같이 주입하면 암컷 마우스에서 장내미생물이 남성화 되지 않아 테스토스테론 상승이 수컷의 맹장 내용물 이식과 장내미생물 변화의 하향 흐름(downstream) 에 있어서 결정적임을 시사하였다.27 이와 반대로 암컷 마우스의 대장 내용물을 수컷에 주었을 때는 에스트로젠이 올라가거나 여성으로서의 특징이별로 나타나지 않아 이 또한 남녀 차이가 있었다.10
(3)장년기 및 노년기의 장내미생물 변화
사춘기에서 성인으로의 이행기에 나타나는 장내미생물의 변화는 일련의 스트레스, 감염, 식이, 항생제에 더 잘 적응하는 방향으로 일어난다.28 만성적인 사회 스트레스는 장 장벽기능(intestinal barrier function) 을 파괴하고 세균의 조성을 변화시키는데, 이때 세균의 림프조직에의 접근이 용이해져 장상피세포의 면역이 활성화된다. 이러한 과정에서 남성에서는 여성에 비해 Bacteroides 와 Prevotella 가 많아짐으로써 스트레스에 대한 장내미생물의 성차를 보인다.29 이렇듯 장년기남녀는 호르몬의 변화에 성차를 보이는데 남성은 테스토스테론을 일정하게 유지하고 여성은 호르몬 변화의 주기를 규칙적으로 타기 때문에 결과적으로 장내미생물의 차이가 나타난다.30,31 유명한 ELDERMET consortium에서는 건강과 관련된 장내미생물과 관련한 노화 현상으로 약함(frailty), 영양 상태, 대사 계수(metabolism score) 염증표지자의 변화를 꼽았는데,32-34 나이가 들면서 나타나는 장내미생물의 핵심적 변화로는 Bacteroides species 및 Clostridium 세균의 비율이 증가 하는 것이다.32 또한 100세가 넘는 사람에서는 Firmicutes 와 혐기성 세균이 증가하는데,35 70세 이상에서는 장의 생리적 기능이 장내미생물의 구성에 영향을 주었다.25,36,37 예를 들어 연령에 따라 증가하는 변비 환자의 대장 미생물의 양상은 대장 이행시간과 비례하는데, 38 이때는 대변이 딱딱해지고 Methanobrevibacter 및 Akkermansia 균이 증가하면서 Ruminococcaceae-Bacteroides 장유형(enterotype) 이 증가하였다.38 이와는 반대로 대장의 빠른 이행시간을 의미하는 설사 내지 무른 변에서는 Firmicutes (Faecalibacterium, Lactococcus, Roseburia) 및 Prevotella 장유형이 증가하였다.39 한편 35,292 명의 독일 성인에서의 연구에 의하면 총 집락형성단위(colony-forming units, CFU) 는 연령 차이를 보였는데, 예를 들어 대장균과 Enterococci spp.는 연령에 따라 증가하고 Bacteroides spp.는 감소하였으며 Lactobacilli 및 Bifidobacteria 는 연령에 따른 변화가 없었다.36,40 한편 6, 31, 74주령과 2년령 수컷 쥐 맹장 내용물에서 SCFA를 직접 측정한 본 연구팀의 실험에서 특히 31주령(인체 나이 30세)에서SCFA 를 주로 생성하는 Lachnospiraceae 와 Ruminococcaceae가증가해서 연령의 차이를 보여주었다(Fig. 2).41 다만 암컷 쥐와의 비교 실험이 안되어 이러한 SCFA 생성 장내미생물의성차를 알아보지 못하였다는 한계가 있었다.
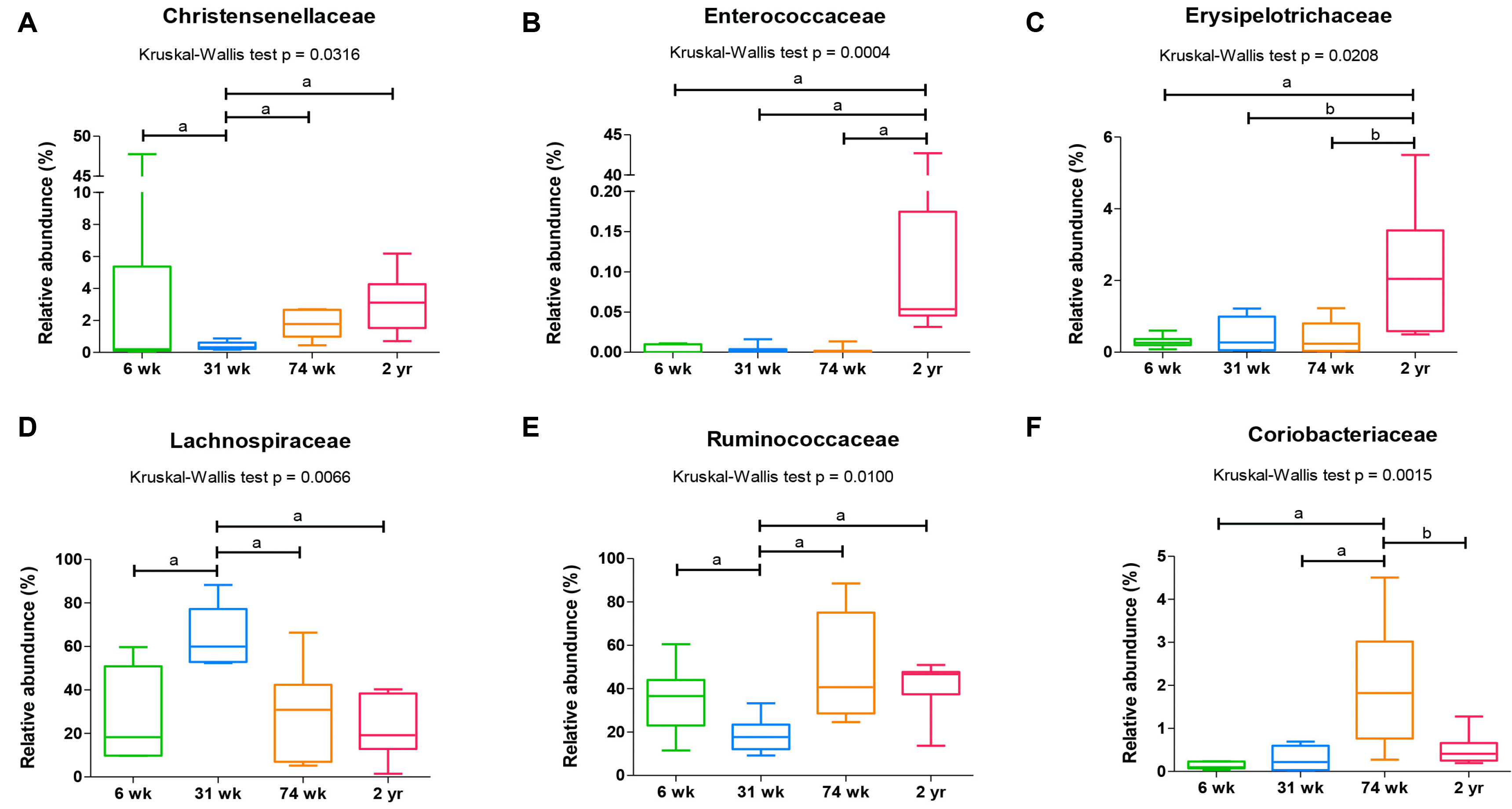 | Fig. 2The compositional characteristics of cecal microbiota vary with age at the (A-F) family level. ap< 0.01 from Mann-Whitney U test with Holm-Bonferroni correction as post hoc analysis followed by Kruskal-Wallis test. Six weeks, 6-week-old; 31 weeks, 31-week-old; 74 weeks, 74-week-old; 2 years, 2-year-old. ap<0.01; bp<0.05. Adapted from Choi et al.41 with permission.
|
(4)폐경 전후의 장내미생물 변화
난소의 활동이 떨어지고 호르몬의 변화가 급격하게 나타나는 여성에게 노화는 매우 역동적인 현상이다. 폐경 후의 장내미생물 변화를 보면 폐경 전에 비해 장내미생물 조성이 다르고(Fig. 3A), 특히 폐경 후 Firmicutes/Bacteroidetes (F/B) 의 비율이 높아지는데, 이는 남성보다 더 높았다(p=0.013) (Fig. 3B).42 이에 더하여 폐경 후에는 Lachnospira, Roseburia 비율이 높아지고 Prevotella, Parabacteroides 및 Bilophila genera 가 증가하면서 혈장 glucagon-like peptide-1, inter-leukin-6 (IL-6), monocyte chemoattractant protein-1 이 증가하였는데,42 이는 호르몬 변화에 의해 장내미생물이 큰 영향을 받는 것을 시사하는 것이다.
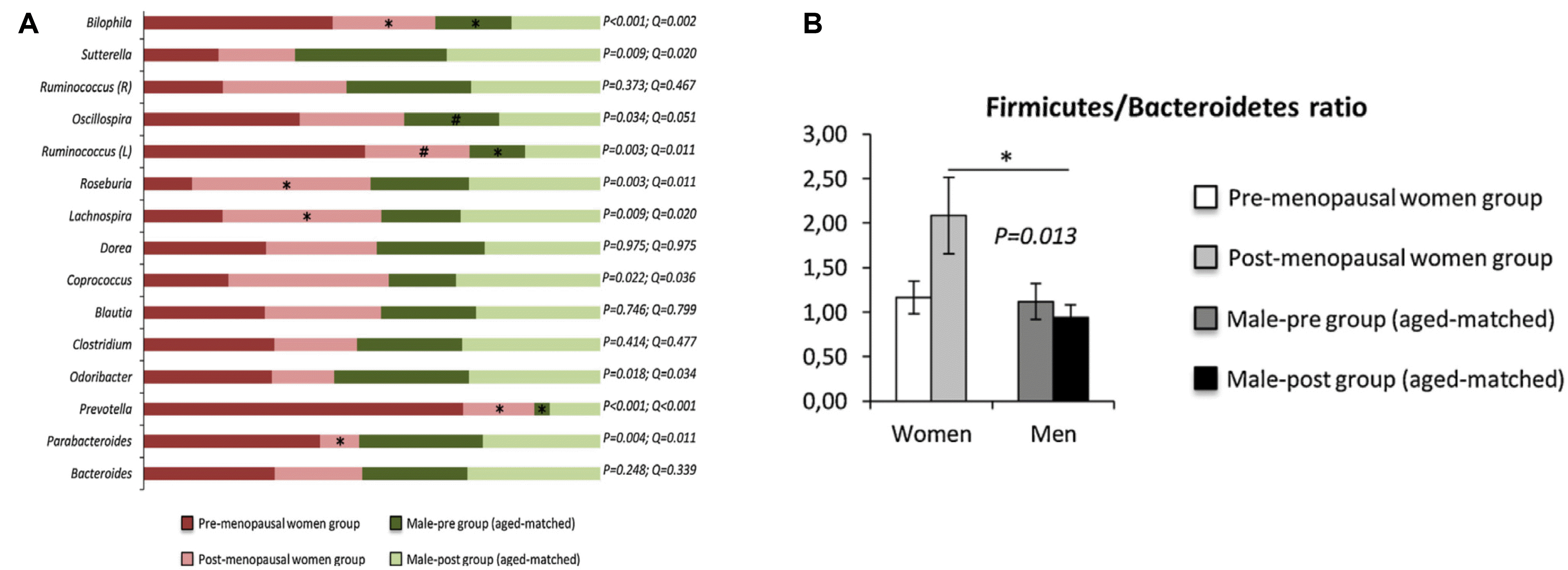 | Fig. 3The changes of gut microbiota after menopause. (A) Gut microbiota differences between pre- and post-menopausal women. (B) A higher Firmicutes proportion in post-menopausal women than in pre-menopausal women. Firmicutes/Bacteroidetes ratio was higher in post-menopausal women (p=0.013) than their corresponding male (age-matched) control group. Adapted from Santos-Marcos et al.42 with permission.
|
2. 장내미생물의 성차
남자와 여자의 신체세포 대비 장내세균의 비율(the ratio of bacteria to human cells, B:H ratio)은 여성에서 2.2로 남성의 1.3 배보다 높아,43-48 장내미생물 연구에서 남녀를 구분하는 것이 중요하다. 1,135명을 대상으로 한 보고를 보면 여성은 장내미생물의 다양성이 높았고 피임제 복용이나 난소절제술에 영향을 받았으며 항생제 내성 유전자를 가지고 있었는데, 이는 아마도 빈번한 항생제 복용과 관련이 있어 보였다.49 최근 장내미생물의 성차에 관한 논문들이 많아지고 있는데,43,50 이는 2014년 이후 차세대 염기서열 분석기법(next-generation sequencing) 이 정교해지면서 좀 더 자세한 정보가 나오고 있기 때문이다. 성호르몬이 면역세포에 미치는 영향도 달라서 여성 에스트로젠이 B 림프구에 영향을 주는 반면 남성 테스토스테론은 T 림프구 증식을 억제하고 Toll-like 수용체 기전을 방해하는 효과가 있었으며 여성 에스트로젠과 달리 장상피세포의 장벽에 영향을 주지 않았다.51
1) 에스트로젠과 장내미생물
생식기관은 정교한 피드백으로 구성되는 다양한 호르몬에 의해 조절되는데 에스트로젠을 분비하는 기관으로는 난소, 부신피질, 지방조직 등이 있다. 에스트로젠 호르몬은 에스트로젠 수용체 β (estrogen receptor β, ERβ)와의 결합을 통해세균의 대사를 직접 조절하기도 하고,52 역으로 세균이 에스트로젠 대사에 적극 개입하기도 한다.3
(1) 에스트로젠의 장내미생물 조절
에스트로젠과 테스토스테론은 장내미생물과 면역세포에 직접적인 영향을 미친다. 즉 β-estradiol은 수지상세포를 자극하여 IL-12 과 interferon-γ를 분비하게 하고 이들은 다시 친염증 사이토카인(proinflammatory cytokines) 의 분비를 활성화한다.51 또한 β-estradiol은 B 림프구의 생존을 늘리고 다세포군 B 림프구(polyclonal B lymphocyte) 를 활성화시킴으로써 장내 침투성의 증가와 함께 친염증 환경을 조성하여 장내미생물이 점막고유층(lamina propria) 으로 이동하고 이들은 다시 염증을 유발하는 염증과정고리(inflammation processes loop)를 형성한다.51 결과적으로 에스트로젠 자체가 면역세포에의 지속적인 영향을 통해 장내미생물의 조성에 영향을 미치고 점막투과성을 증가시킴으로써 장내미생물의 분포에 관여함을 시사하였다. 한편 장내미생물은 에스트로젠의 영향을 받을 뿐 아니라 장내미생물 또한 에스트로젠 대사에 관여함이 알려졌다.3 즉, 결합에스트로젠(conjugated estrogens) 이 혈액을 통해 돌다가 담즙과 신장으로 분비되면 장내미생물이 가지고 있는 β-glucuronidase가 담즙으로 흘러나온 에스트로젠을 비결합형(deconjugated forms) 으로 변화시킨다(Fig. 4A).3 이러한 활동성 비결합형 에스트로젠이 혈중으로 흡수되어 장간순환(enterohepatic circulation) 으로 들어가 생식기관 외 근육, 신경계 그리고 혈관계 에스트로젠 수용체 ERα 및 ERβ에 작용한다(Fig. 4A).3 다만 장내세균 중에는 β-glucuronidase가 풍부한 세균이 있는 반면 없는 세균도 많아 에스트로젠의 혈중 농도에 영향을 주는 세균은 한정되어 있다(Fig. 4B).3 남성과 폐경기 여성에서는 소변의 총 에스트로젠 수치와 분변 미생물의 풍부도와 a-다양성(a-diversity) 사이에 연관성이 있고 난소가 아닌 다른 기관에서 분비되는 에스트로젠도 대변Clostridia 및 Ruminococcaceae 와 관련성이 있으며 대변 β-glucuronidase 활성도는 대변 에스트로젠과 반비례 관계가 있었다.50 그러나 폐경기 전 여성에서는 이러한 결과가 달라 장내미생물의 에스트로젠 대사 관여에 대해서 난소 에스트로젠과 비난소(non-ovary) 에스트로젠이 다른 영향을 받음을 시사하였다.53 즉, 폐경기 전 여성의 에스트로젠은 장내미생물이나 β-glucuronidase 효소와 관련성이 없고 비난소에서 나오는 에스트로젠이 장내미생물이나 β-glucuronidase와 연관성이 있음이 추측되었다.53
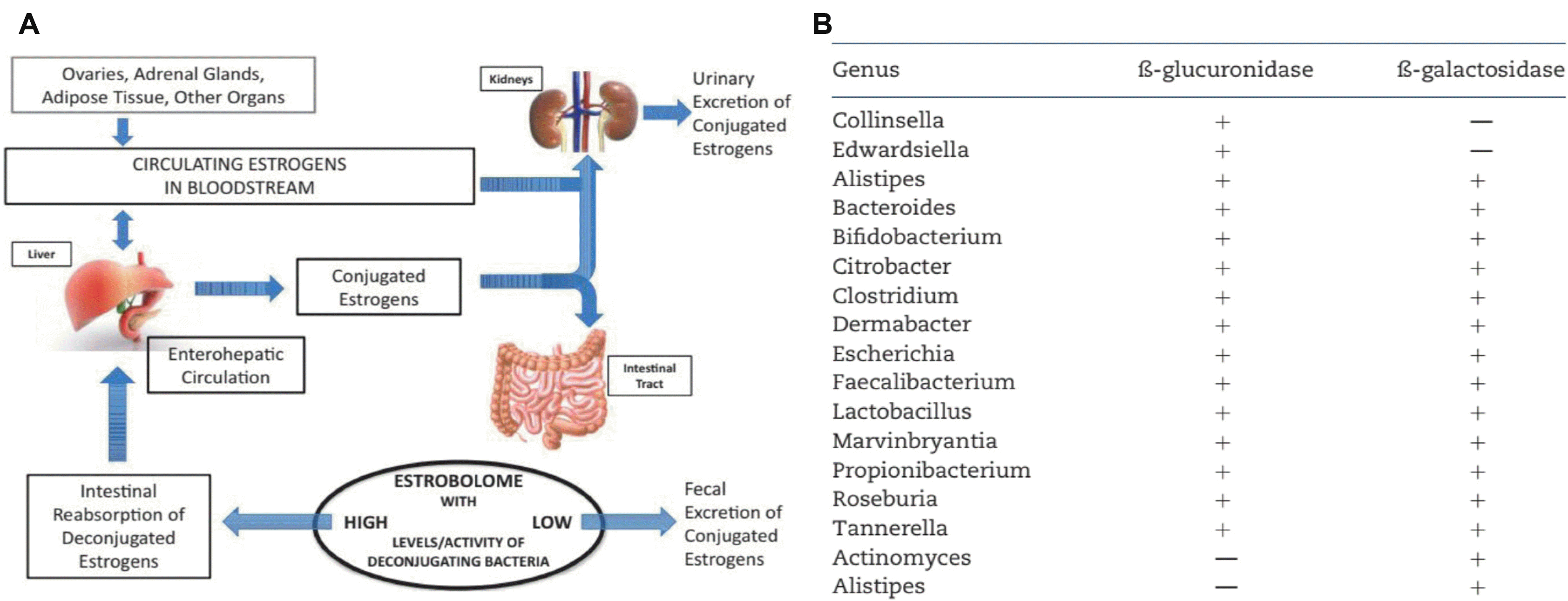 | Fig. 4The estrobolome and enterohepatic circulation of estrogens. (A) Estrogens are primarily produced in the ovaries, adrenal glands, and adipose tissue and circulate in the bloodstream in free or protein-bound form and first undergo metabolism in the liver, where estrogens and their metabolites are conjugated. Conjugated estrogens are eliminated from the body by metabolic conversion to water-soluble molecules, which are excreted in urine or in bile into the feces. The conjugated estrogens excreted in the bile can be deconjugated by bacterial species in the gut with beta-glucuronidase activity (constituents of the ‘estrobolome’), subsequently leading to estrogen reabsorption into the circulation. (B) Several gut microbiota have beta-glucuronidase. Adapted from Kwa et al.3 with permission.
|
2)안드로젠과 장내미생물
안드로젠 또한 에스트로젠과 비슷하게 장내미생물과의 상호 긴밀한 관련성을 보인다. 즉, 안드로젠에는 비결합형과 글루큐론산형(glucuronidated form) 이 있는데 마우스에서의 글루큐론산형 테스토스테론(glucuronidated testosterone) 과 다이하이드로테스토스테론(dihydrotestosterone, DHT) 은 소장에서 매우 높은 반면 free DHT형은 대장 말단에서 높았다.54 또한 나이 어린 수컷 쥐에서는 free DHT가 혈청보다 대변에서 70배나 높았는데, 무균 마우스에서는 글루큐론산형테스토스테론과 DHT는 높지만 free DHT 는 낮음이 알려지면서 장내미생물이 안드로젠의 탈글루큐론산화에 관여함이 시사되었다.54 남성에서의 테스토스테론 및 여성의 에스트로젠과 장내미생물과의 상관관계를 살펴본 결과 남녀 각각 테스토스테론과 에스트로젠이 높은 사람에서는 장내미생물의 다양성이 높음이 밝혀졌다.55 즉, 테스토스테론이 높은 남성에서는 Acinetobacter, Dorea, Ruminococcus, Megamonas가 많았고, 에스트로젠이 높은 사람에서는 Bacteroidetes 가 높은 반면 Firmicutes 가 낮았으며 에스트로젠이 낮은 여성에서는 Slackia 그리고 Butyricimonas Genera가 높았다.55
3. 위장관 질환에서의 장내미생물 성차
기능성 위장관장애(functional gastrointestinal disorders, FGIDs) 의 병태생리는 위장관 운동기능의 이상, 내장과민성, 감염, 염증, 장내미생물의 변화 등이 중요한 병태생리로 생각되고 있다. 감각 이상, 운동기능의 변화, 자율신경계와 중추신경계의 통합된 활성 그리고 장내미생물의 영향의 결과로 위장관 증상이 발생한다는 뇌-장관-장내미생물 축이라는 개념이 제시되고 있으며 이들 질환은 여성에 호발한다. Helicobacter pylori의 만성 감염으로 발생하는 위암, 장내미생물에 영향을 받는 대장암의 경우도 남성에서 그 발병률이 약 2배 높다. 또한 역학이나 병인에 있어 성차를 보이는 만성 염증성 장질환에서의 에스트로젠의 역할 및 면역과 긴밀한 연관성을 가지고 있는 장내미생물의 성차에 대해서도 최근 알려지기 시작하였다. 이에 본고에서는 연구 결과가 나오고 있는 과민성 장증후군, 만성 염증성 장질환과 대장암에서의 장내미생물의 성차에 대해 간략하게 알아보고자 한다.
1) 과민성 장증후군
과민성 장증후군은 기질적 이상 없이 변의 형태나 배변 습관의 변화와 함께 복통이나 복부 불편감이 반복되는 것을 특징으로 하는 만성 기능성 위장관장애(FGIDs)이다. 과민성 장증후군은 매우 흔한 질환 중의 하나로 여성의 유병률은 7-24%, 남성은 5-19%로 여성에서 많다.56-58 일차 의료기관을 방문하는 가장 흔한 원인 중의 하나이고 소화기 증상뿐만 아니라 편두통(migraine), 섬유근통(fibromyalgia) 등의 소화기와 관련이 없는 증상이 잘 동반되기도 한다.59,60 과민성 장증후군은 여자에서 좀 더 많이 발생할 뿐 아니라 감염 후 과민성장증후군의 경우가 많고 설사형이 많은 남자와는 달리 여자는 뇌-장관축(brain-gut axis) 의 영향을 더 많이 받고 변비형이 많다.61 최근에는 뇌-장관축(brain-gut axis) 에 영향을 많이 줄 뿐 아니라 장벽에 큰 역할을 하는 장내미생물을 넣어 뇌-장관-장내미생물축(brain-gut-microbiome axis) 이라는 학문적 용어가 만들어진 바 있다.
(1) 과민성 장증후군에서 성호르몬의 영향
과민성 장증후군에서 관찰되는 장 운동성 변화, 내장과민성, 자율신경계 이상 등이 뇌-장관축에 의하여 조절된다고 알려져 있다(Fig. 5).62 이에 더하여 여성호르몬은 신경조정시스템과 감정시스템과 연관관계를 가지면서 스트레스, 장 운동 및 내장통에 대한 감수성에 영향을 준다.63 또한 에스트로젠과 프로제스테론은 평활근 수축을 억제하며 프로제스테론은 대장의 연동운동을 조절하는 세로토닌(5-hydroxytryptamine) 시스템을 조절한다고 알려져 있는데 이로 인해 과민성 장증후군 여자 환자는 혈액 난소 호르몬이 낮은 월경을 하지 않는 시기 외에 변비 증상을 호소한다고 해석되고 있다.64 또한 여자는 남자보다 위장관 통과시간이 길고,65 여성의 난소호르몬의 농도가 높은 시기에 대장 통과시간이 더 느려진다고 알려진 바 있어 성호르몬이 대장 수축과 밀접한 관련성이 있음을 알 수 있다.58 한편 남성호르몬의 역할에 대해서도 알려지기 시작하고 있는데, 안드로젠은 만성 통증증후군을 억제하고 실험적 통증모델에서 진통 효과가 있다고 알려진 바 있다.66 아마도 안드로젠 농도, 수용체, 활동 부위가 과민성 장증후군과 만성 통증증후군 발생에 역할을 하는 것으로 추측된다.67 최근 연구에 의하면 과민성 장증후군이 있는 중년기 남자가 정상인에 비해황체형 성호르몬이 낮고 과민성 장증후군 증상이 테스토스테론과 반비례관계가 있음이 보고된 바 있다.68 젊은 남자 과민성 장증후군 환자에서 성호르몬결합 글로불린(sex hormone-binding globulin) 과 테스토스테론이 증가되어 있는 것은,68 과민성 장증후군 발생에 있어 남성호르몬도 어떤 역할을 하지 않을까 추측되지만 과거 보고와는 조금 상반된 것이라 추후 더 많은 연구가 필요하다 생각된다.
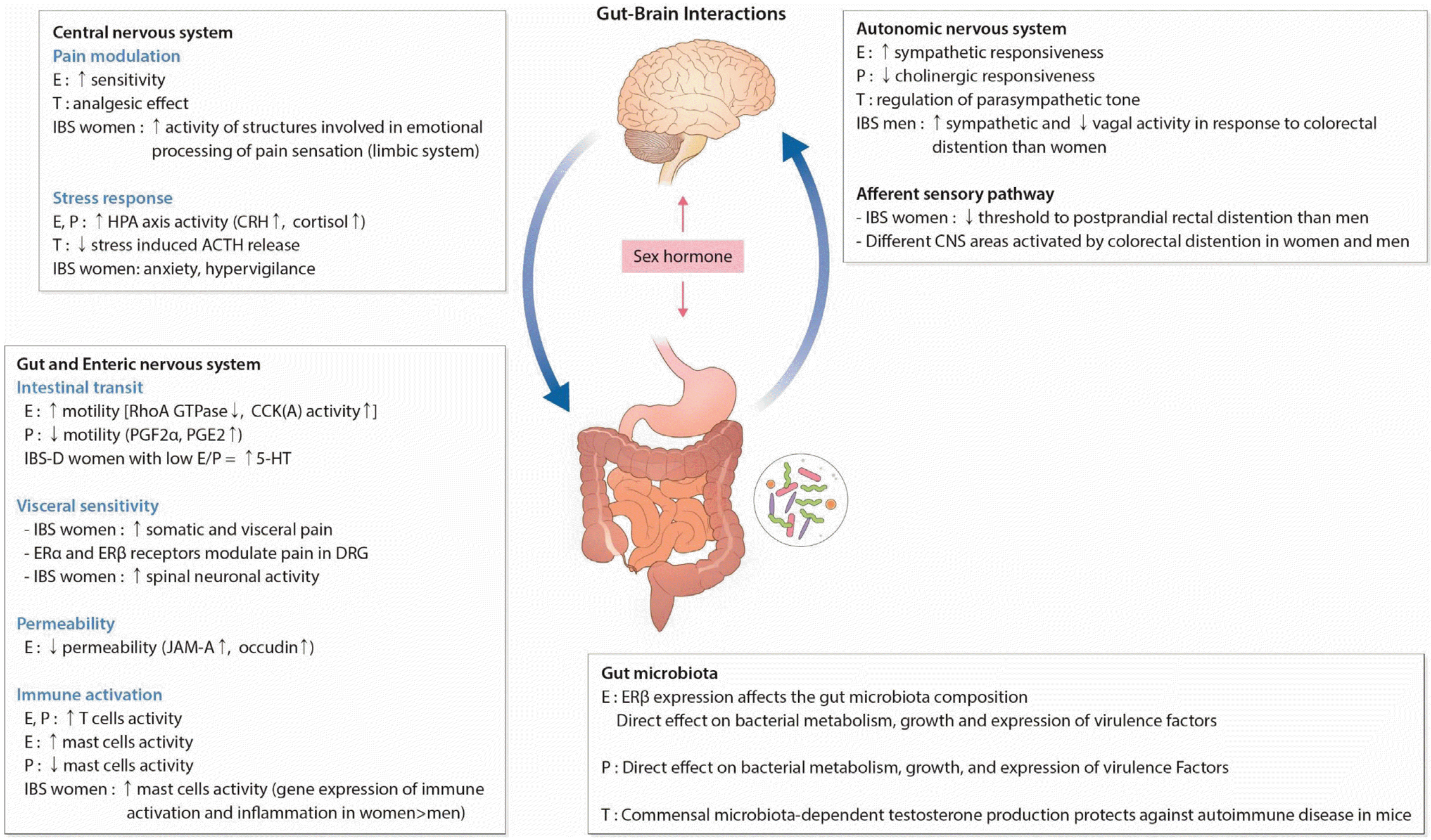 | Fig. 5Brain-gut axis and sex hormones interaction in irritable bowel syndrome (IBS). Sex hormones influence peripheral and central regulatory mechanisms involved in the pathophysiology of IBS contributing to the alterations in stress response, visceral sensitivity and motility, intestinal barrier function, and immune activation of intestinal mucosa. Sex hormones also have direct effects on the gut microbiota and enteric nervous system. E, estradiol; T, testosterone; P, progesterone; CRH, corticotropin-releasing hormone; ACTH, adrenocorticotropic hormone; GTP, guanosine-5’- triphosphate; CCK, cholecystokinin; PG, prostaglandin; 5-HT, 5-hydroxytryptamine; ER, estrogen receptor; DRG, dorsal root ganglion; JAM, junctional adhesion molecule. Adapted from Kim and Kim62 with permission.
|
(2) 과민성 장증후군에서 장내미생물의 성차장내미생물 연구에 있어서는 대변과 장 점막의 조직 검사를 통해 진행하는 방법이 있다. 장관 내에 존재하는 미생물(대변에서 검출되는 미생물)과 장 점막에 부착된 미생물 중 장내미생물의 대부분을 차지하는 세균은 대사 과정을 통해 장의항상성 유지에 중요한 역할을 하며 탄수화물 발효 및 가스생성을 통해 과민성 장증후군 환자에서의 복부팽만감이나 방귀 증상을 유발할 수 있다. 반면 장 점막에 부착된 세균은 상대적으로 양이 적지만 장 점막 면역체계와 상호작용에 의한 면역 활성화 등을 통해 관련 증상을 유발할 수 있다. 이처럼 장내미생물은 장의 정상적인 생리 및 면역 조절에 중요한 역할을 하는데, 과증식된 장내미생물에 의한 장 운동 이상, 면역활성화(immune activation) 그리고 비정상 발효 등이 과민성장증후군 발생에 기여할 것으로 생각된다. 보통 대변에서의 문(phylum) 에 속하는 세균의 조성은 Firmicutes, Bacteroidetes, Actinobacteria, Verrucomicrobia 순인데 본 연구자들이 12명의 대조군과 7명의 설사형 과민성 장증후군 환자의 대변을16S rRNA metagenomic 시퀀싱 한 결과 과민성 장증후군에서는 대조군보다 Firmicutes 는 감소하고, Bacteroidetes 가 거의 두 배, Proteobacteria 와 Fusobacteria phylum (문, 門)이 증가하는 양상을 보였다(Fig. 6).69 또한 phylogenetic investigation of communities by reconstruction of unobserved states 분석한 결과 Acidaminococcaceae, Sutterellaceae 및 Desulfovibrionaceae 는 과민성 장증후군 환자에서 증가하였고 Enterococcaceae, Leuconostocaceae, Clostridiaceae, Peptostreptococcaceae, Lachnospiraceae는 과민성 장증후군 환자에서 의미 있게 감소하였다(Fig. 7).69 이처럼 과민성 장증후군에서의 장내미생물 변화는 확실한데, 특히 담즙재흡수에 영향을 주는 이차 담즙산 생성(pathway ko00121) 에 관여하는 K07007 (baiN, 3-dehydro-bile acid delta 4, 6-reductase) 및 K15874 (baiI, bile acid 7 beta-dehydratase) 가 대조군보다 과민성 장증후군에서 의미 있는 감소를 보임으로써(Fig. 7J-7K),69 설사형 과민성장증후군의 발생에 대한 장내미생물의 영향을 시사하였다.69 하지만 대조군 12명, 설사형 과민성 장증후군 7명으로 예수가 너무 작아 남녀 차이를 분석하기는 어려웠다. 이러한 직접적인 증거 외에도 장내미생물이 뇌-장관축과 장벽(barrier)에 중요한 역할을 하며(Fig. 5),62,70,71 장내미생물의 지속적인 감소와 뇌-장관축의 평형이 깨지면 저도의 염증과 면역기전의 불균형이 유발되고 이들은 장내 분비, 내장통, 장운동의 변화로 이어진다고 설명되고 있다.72 성호르몬은 세균 대사에 영향을주고 에스트로젠 수용체 ERβ를 비롯한 steroid nuclear 수용체 발현을 통해 세균의 독성인자 발현에 영향을 준다.52 또한 유아 시기에 미생물 노출은 성호르몬 농도에 영향을 주어 인슐린 의존형 당뇨 마우스 모델에서 자가면역 질환 진행을 변경한다고 알려진 바 있다.27 이와 비슷한 맥락으로 사춘기 수컷 마우스 모델에서 테스토스테론의 분비가 많아지면 장내미생물에 변화가 오고 자가면역 질환과 관련이 있는 T와 B림파구에 변화를 가져온다고 보고되었다.27
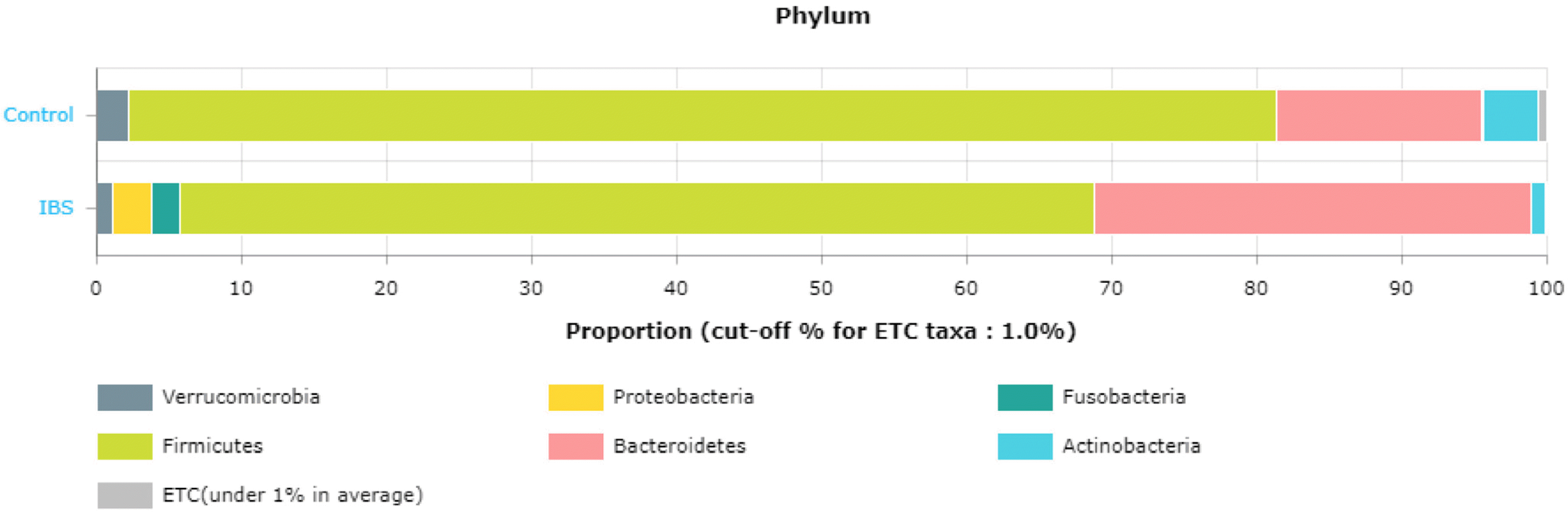 | Fig. 6Compositional and functional changes in the gut microbiota in irritable bowel syndrome patients. In term of phylum Firmicutes decreased and Bacteroidetes, Proteobacteria, and Fusobacteria increased. IBS, irritable bowel syndrome. Modified from Lee et al.69 with permission.
|
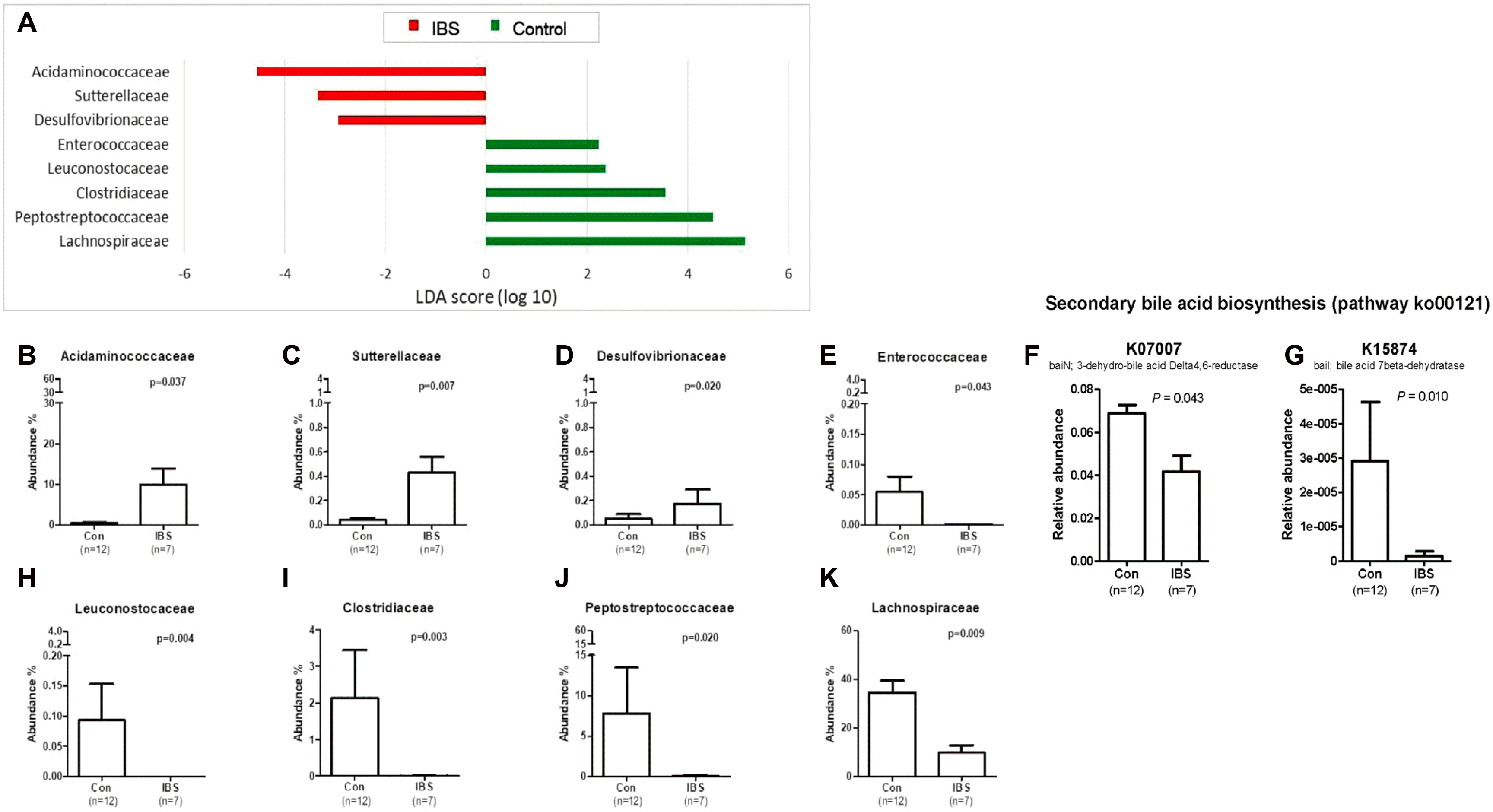 | Fig. 7Differences in the microbiota composition between irritable bowel syndrome (IBS) samples and normal control samples. (A) Linear discriminant analysis (LDA) effect size (LEfSe) of the gut microbiota at the family level. Abundance ratios of the families that differed between the IBS and normal control groups according to LEfSe: (B) Acidaminococcaceae, (C) Sutterellaceae, (D) Desulfovibrionaceae, (E) Enterococcaceae, (F) Leuconostocaceae, (G) Clostridiaceae, (H) Peptostreptococcaceae, (I) Lachnospiraceae, (J) K07007 (baiN, 3-dehydro-bile acid delta 4,6-reductase) and (K) K15874 (baiI, bile acid 7 beta-dehydratase). p-values were assessed with the Wilcoxon rank sum test. Con, control; IBS, irritable bowel syndrome; LDA, linear discriminant analysis. Adapted from Lee et al.69 with permission.
|
한편 과민성 장증후군에서 소장 세균 과증식(small intestinal bacterial overgrowth, SIBO) 이 중요한 역할을 할 수 있으리라는 것이 제시되어 왔다. 정상적으로 위는 연동 운동 및 위산으로 인한 낮은 pH 때문에 상대적으로 적은 수의 세균을 포함하고 있어 위 내용물 1 mL 당 0-1×103 CFU/mL 인 반면 십이지장 및 공장에는 1~1×104 CFU/mL, 근위부 회장에는 1~1×105 CFU/mL, 말단 회장에는 1×105~1×108 CFU/mL 그리고 대장에는 1×1010~1×1012 CFU/mL 의 세균이 존재한다. 소장에는 주로 Lactobacillus spp., Enterococcus spp., Streptococcus spp. 그리고 구강 내 상재균인 양성 호기성세균과 통성 혐기성 세균(facultative anaerobes) 이 존재하며 대장에는 Bacteroides spp., Clostridium spp., Bifidobacterium spp., Fusobacterium spp., Eubacterium spp. 와 같은 혐기성 세균이 약 99.9%로 대부분을 차지한다. SIBO는 대장 및 원위부 소장에 상재하는 세균이 근위부 소장으로 확장되어 서식하는 것을 말하며 일반적으로 근위부 소장에서 세균이 1×105 CFU/mL 이상으로 증식하여 존재하는 경우로 정의되는데, 최근 과민성 장증후군에서 SIBO 의 유병률을 조사한 12개 연구의 메타분석 결과를 보면 정상인과 비교하여 과민성 장증후군에서의 SIBO 는 4.7배 높다고 보고하였으나 측정 방법이나 진단 기준이 일정하지 않고 일관된 결과를 보여주지 못하여 추가 연구가 필요한 상황이다. 또한 여자에서 변비형 과민성 장증후군이 많고 복부팽만감 및복통이 많았기에 아마도 SIBO 도 여자에 많지 않을까 추측되지만 성차에 대해서는 전혀 언급된 바가 없어 추후 연구가 더 되어야 할 분야이다.
2) 만성 염증성 장질환
염증성 장질환(inflammatory bowel disease, IBD) 은 소장 및 대장에서 면역체계 조절 이상으로 인한 만성 재발성 염증질환들에 대한 통칭이다. 염증성 장질환의 대표적 질환은 크론병(Crohn’s disease, CD) 과 궤양성 대장염(ulcerative colitis, UC)인데 면역반응 조절 이상이 중요한 기전으로 생각된다.73,74 최근 IBD의 성차에 대한 연구 보고 중 남녀에서의 발생 빈도에 대한 연구가 많은데, 이들은 공통적으로 성호르몬이 차이가나는 연령에 따라 다름을 보여주고 있다. CD의 경우 소아에서는 남자에서의 발생이 높다가 사춘기가 넘어서면 여자에서의 발생률이 월등하게 높아지는 것이다(Fig. 8).75 이와는 다르게 UC의경우 45세까지는 남녀에서 다르지 않다가 45세 이후에는 남자에서 높은 발생률을 보인다.76 이러한 성차는 성호르몬에 따른 장내미생물의 차이가 가장 큰 원인으로 설명되고 있고 이를 매개하는 중요한 기전으로 성호르몬에 의해 조절되는 면역세포들이 있다(Fig. 9).75 이러한 보고는 IBD 연구에 있어 남녀 각각을 분리하여 분석해야 하고 동물 모델을 사용하는 경우 수컷과 암컷 모두에서 연구해야 함을 시사한다.
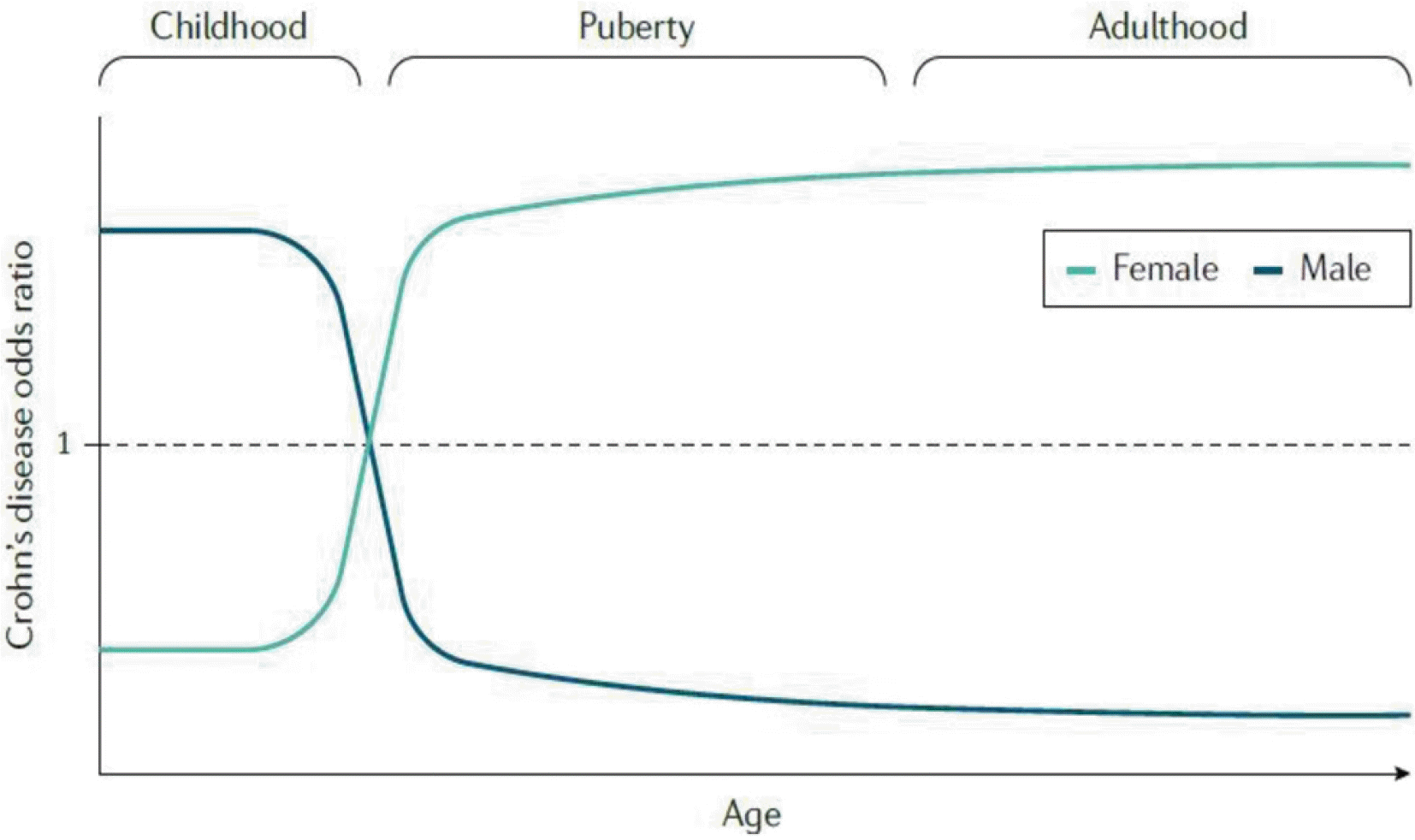 | Fig. 8Sex-specific risk of Crohn’s disease throughout the lifespan in Western countries. During early childhood, boys have a higher risk than girls of developing Crohn’s disease. However, around the time of puberty, the risk of Crohn’s disease in girls increases substantially, potentially owing to shifting hormone levels. The majority of population-based studies of Western cohorts show an increased risk of Crohn’s disease among women compared with men. Adapted from Goodman et al.75 with permission.
|
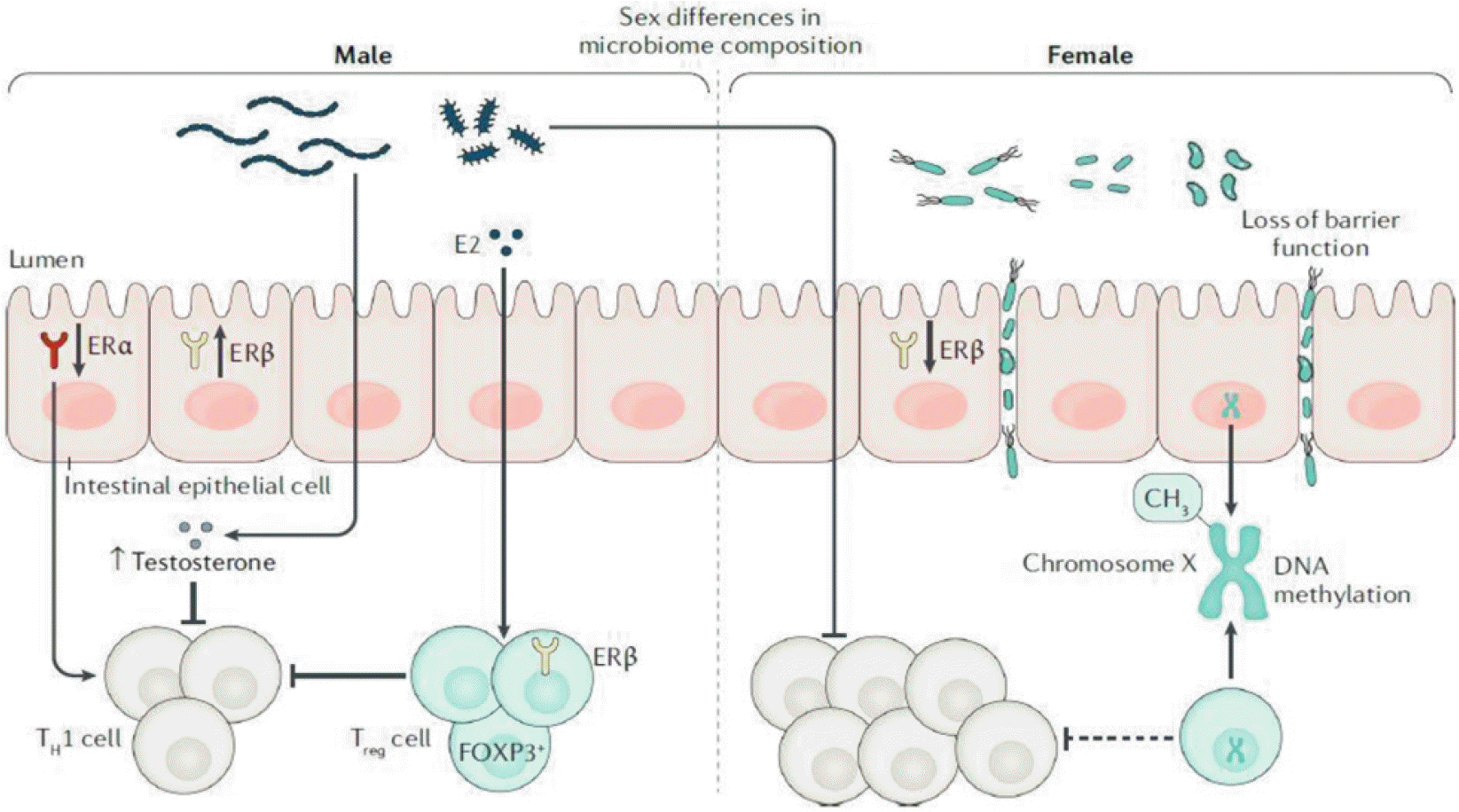 | Fig. 9Eestrogen (E2) has also been shown to have immunoprotective effects during experimental colitis by inducing regulatory T (Treg) cell expansion and function in male mice in an oestrogen receptor (ER)-β-dependent manner. In addition, decreased ERβhas been observed in both epithelial and Treg cells from female mice prone to ileitis and/or colitis, which can promote intestinal epithelial barrier dysfunction and worsening of disease. Interestingly, decreased expression of ERαin male mice results in exacerbation of intestinal inflammation, but has the opposite effect in female mice, suggesting that skewing towards ERβ signalling induces global protection from colitis. Finally, increasing evidence indicates that differential transcription (for example, by DNA methylation) of genes located on chromosome X can regulate molecules that can also affect downstream sex-based differences in the pathogenesis of IBD. TH1 cell, T helper 1 cell. Adapted from Goodman et al.75 with permission.
|
(1) IBD 에서의 성호르몬의 영향
에스트로젠은 유해균으로부터 숙주를 보호하는 내장 장벽의 밀착연접관련 단백질 발현에 영향을 주는데 azoxymethane/dextran sulfate sodium (AOM/DSS) 으로 대장염을 유발한 마우스 모델에서 17β-estradiol 을 처치하면 mucin 2 (MUC2), zonula occludens 1 (ZO-1), OCLN, claudin 4 (CLDN4) 유전자의 발현 증가로 대장 투과성이 감소하고 염증과 관련된Krüppel-like factor 4 (KLF4), nuclear factor κB (NF-κB), inducible nitric oxide synthetase (iNOS), cyclooxygenase-2 (COX-2) 유전자와 단백질 발현이 감소함을 보고한 바 있다.77 이러한 결과는 정상 여자 대장상피세포주인 CCD841CoN 에서 17β-estradiol 을 주었을 때 NF-κB와 COX-2 발현을 감소시키고 항산화효소인 heme oxygenase-1 (HO-1) 과 NAD(P)H-quinine oxidoreductase-1 (NQO-1) 발현을 증가시켜서 염증을감소시키는 것으로 확인한 바 있다.78
여성의 성호르몬인 17β-estradiol, 프로제스테론(progesterone) 그리고 황체형성호르몬(luteinizing hormone) 은 장과 관련된 기능 즉 위 수축, 장관이행 및 통증에 대한민감도에 영향을 주는데,75 특히 에스트로젠은 IBD 발생 그리고 임상 증상이 심해지는 것과 관련이 있다.75 이것은 경구피임제(oral contraceptive pills) 사용 후 IBD가 악화된 예와 관련성이 있다.79,80 2008 년도에 시행된 메타분석을 보면 20만 명의 여성 CD나 UC 환자에서 경구 피임제에 노출되면 CD에 걸릴 상대위험도(RR) 가 1.46 (95% CI 1.26-1.70) 로 높았다. 또한 덴마크에서의 자궁내막증(endometriosis) 에 걸린 사람 중에서IBD에 걸릴 확률(standardized incidence ratio) 은 자궁내막증이 없는 경우에 비해 1.5 배(95% CI 1.4-1.7) 로 높아 증가된 에스트로젠이 IBD 를 악화시킬 수 있음을 시사하였다.81 이 외에도 에스트로젠 상승과 IBD 증상과의 관련성은 월경 시기 근처에 IBD 증상이 심해지는 현상과 IBD 진단 12개월 전 월경불순 현상이 나타난 사실 등이 이를 좀 더 반증하고 있다.82
(2) IBD 환자에서의 장내미생물의 성차
여러 질환에서 성차에 따른 장내미생물이 병인으로 밝혀지고 있는데 IBD도 이에 포함되고 있다.83 그 이유는 IBD 발생에서의 면역의 불균형이 매우 중요한데 장내미생물이 이러한 면역불균형을 유발하기 때문이다.84 마우스 IBD 모델에서의 연구를 보면 장내미생물의 변화가 오는 경우가 많은데,78,85 특히 난소절제술 후 장내미생물의 변화가 온다.27,51,86,87 수컷 마우스 장염에서 에스트로젠은 regulatory T (Treg) cell 확장을 통해 면역보호 효과를 보이는데 이는 ERβ에 비례하였다(Fig. 9).75,83 또한 암컷 쥐에서 ERβ가 감소하면 Treg cells가 감소하면서 소장 염증과 대장 염증이 심해진다고 알려지고 있다(Fig. 9).83 수컷쥐에서 ERα 발현이 감소하면 장 염증이 증가하지만 암컷쥐에서는 반대 현상이 나타나는 것으로 보아 ERβ signaling이 강해지면 장염 보호 효과를 나타내는 것으로 생각된다.83 또한 급성 2, 4, 6-trinitrobenzenesulfonic acid (TNBS) 수컷과 암컷 동물모델에서의 대변 미생물 연구 결과를 보면 야생형(wild type) 과 tumor necrosis factor (Tnf)−/− 수컷과 암컷의 염증이 달랐다. 야생형과 Tnf−/− 모두 수컷에서 암컷보다 염증이 심하였고 연령에 따라 염증 정도가 달랐다.88 한편 C57BL/6 마우스 대변 장내미생물 분석에서 수컷과 암컷 사이에 β-다양성에 차이가 있었으나 염증 모델로 사용되는 IL-10 결핍(knock out) 마우스에서 이러한 차이가 없어지고 F/B 비율 또한 수컷쥐에서 암컷 쥐에서보다 증가하여 이러한 면역 변화가 장내미생물에 큰 영향을 주는 것을 시사하였다.85 또한 암컷 쥐에서는 Proteobacteria가 야생형보다는 IL-10 결핍 마우스에서 증가하는 등 성차를 보였다.85 이러한 염증성 장질환 모델에서의 장내미생물 변화는 성차에 따라 면역 변화와 연계하여 만성장염 발생에 역할을 함을 알 수 있다.
3) 대장암
소화기 질환에서의 성차가 존재하는 대표적 질환은 대장암 (colorectal cancer)이다. 대장암의 전구 단계인 대장 선종(colonic adenoma) 과 대장암의 역학에서 남녀 차이가 극명한데 대장암의 생존율은 여성에서 높은 것으로 알려져 있다. 이러한 성차의 원인으로 여성호르몬인 에스트로젠이 대표적으로 연구되어 왔고 식이 등 사회환경적 요인도 연구 대상이다. 여성은 남성보다 대장암 발생이 5-7세 늦으며 우측대장(상행결장)에 대장암이 많이 발생하며, 우측에 납작한 용종(톱니 선종, serrated adenoma) 이 빈번히 발견되는 특징을 가지고 있다.56 또한 중간암(interval cancer, 대장경을 3-5년 이내에 받은 환자에서 발생하는 대장암)의 발생 비율은 여성에서 높은데, 그 이유로 우측대장 톱니 선종의 대장경 발견이 더디거나 염색체 불안전성(chromosomal instability, CIN pathway) 기전보다 빠른 속도로 진행하는 현미부수체 불안정성(microsatellite instability) 및 CpG island methylator phenotype 경로가 여성 대장암에서 더 주된 기전이기 때문으로 설명되고 있다.56,57 한편 폐경 후 여성에서 호르몬 대체요법으로 대장암 위험이 56% (p=0.003) 감소한 바 있어 여성호르몬의 대장암 발생 억제 효과를 잘 보여주고 있는데, 에스트로젠수용체 중 ERβ가 중요하다. 에스트라다이올(E2)과 결합한 ER β는 에스트로젠 반응요소(estrogen response element)와 결합하거나 전사인자들과의 상호작용으로 유전자의 전사를 직접적으로 조절하는 기능이 있다.58 한편 이러한 남녀의 대장암성차에 관여하는 요소로서 성호르몬과 밀접한 관련이 있는 장내미생물이 거론되고 있다.87
(1) 대장암에서의 성호르몬의 영향
최근 대장암에서의 성호르몬의 영향을 보기 위해 AOM/DSS 염증성 대장암 동물모델을 이용한 연구가 시행되고 있다. AOM/DSS 대장암 모델 연구에서 암컷에 비해 수컷 마우스의대장암 발생률이 높았으며, 수컷에 에스트로젠 처리를 하면 대장암 발생률이 낮아졌다.89 이러한 17β-estradiol의 대장암예방효과는 항산화, 항염증을 매개하는 전사 인자인 nuclear factor erythroid 2- related factor 2 (Nrf2) 에 의해 매개됨이 밝혀졌다.90 또한 암컷 AOM/DSS 대장암 모델에서 난소절제술을 하면 우측대장암 발생률이 증가하다가 에스트로젠 투여에 의해 감소함으로써 에스트로젠이 우측대장암 발생을 억제함을 알 수 있었다.91 이와 반대로, 수컷 AOM/DSS 대장암모델에서 정소 절제술을 시행하면 좌측대장암 발생이 의미 있게 감소하며, 테스토스테론을 주입하면 대장암 발생이 다시 증가함으로써 테스토스테론이 좌측대장암 발생에 관여함을 확인하고92 이러한 대장암의 성차를 나타내는 근저에 장내미생물이 역할을 하고 있음을 시사하는 결과를 발표한 바 있다.87
(2) 대장암에서의 장내미생물의 성차
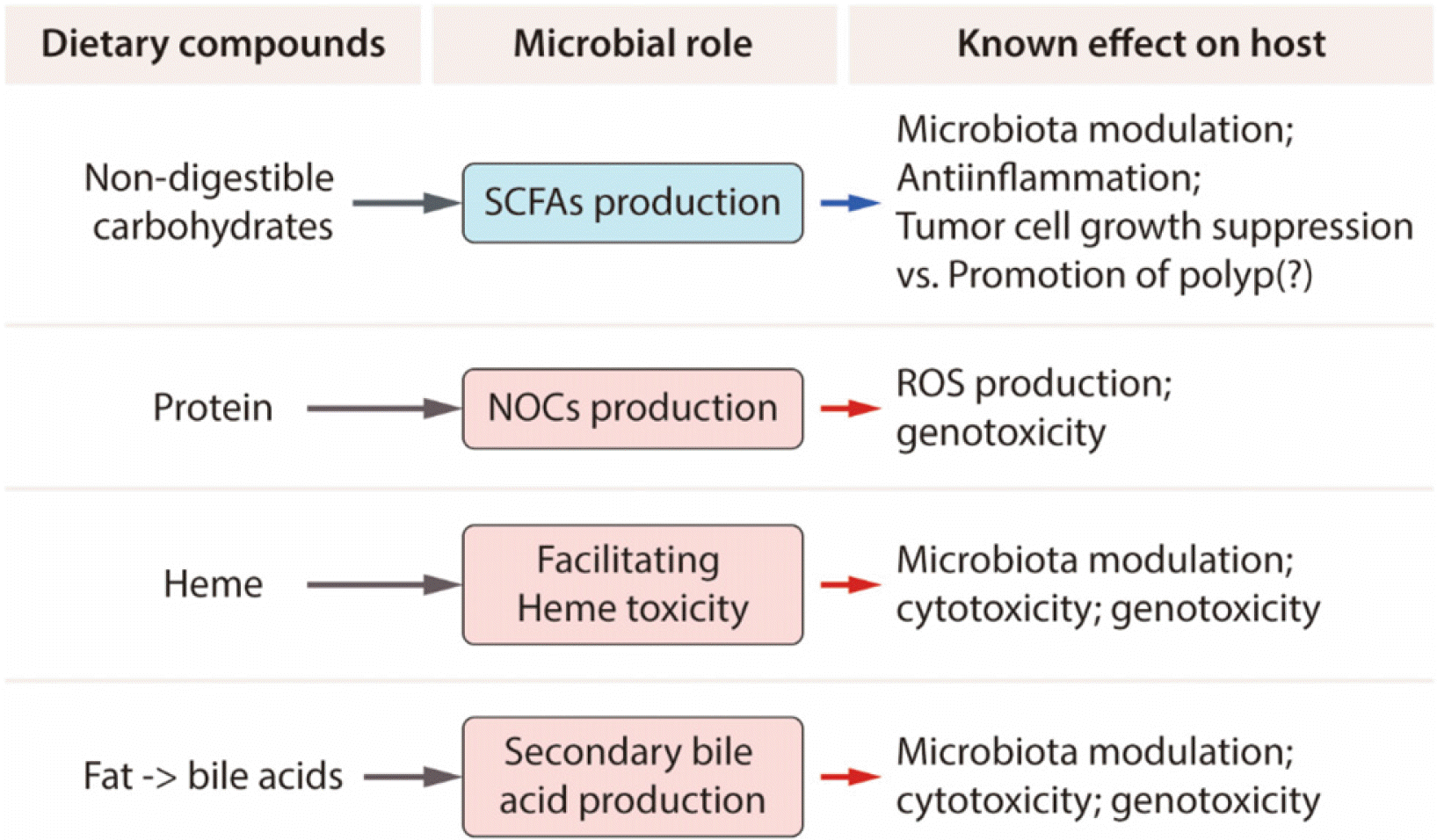
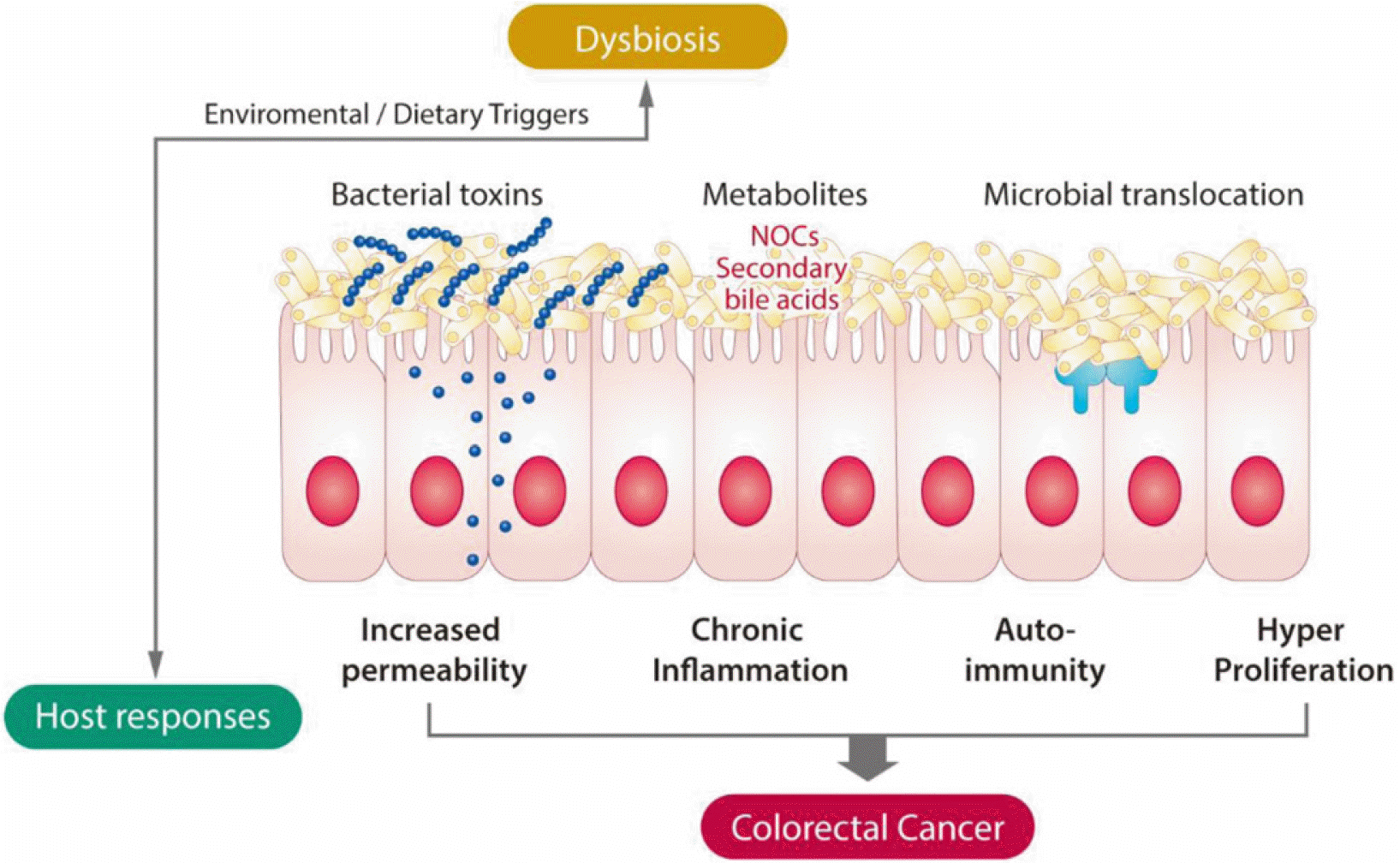
에스트로젠의 대장암 발생 억제효과는 에스트로젠에 의한 F/B ratio 감소 그리고 α-다양성을 나타내는 Simpson Index 의 증가와 같이 장내미생물의 변화도 일부 기여함이 시사되었다.87 남녀는 식이에 대한 선호가 다른데, 예를 들면 남자는여자에 비해 육식을 좋아하고 여자는 채소와 과일을 선호하는 것으로 나타나고 있고 이들이 남녀에서의 대장암 발생 차이를 유발하는 것으로 알려진 바 있다. 이러한 식이 선호도에 따라 장내미생물의 분포가 달라지고 이들 장내미생물이 분비하는 내용물이 달라진다. 장내미생물이 대장암 발생에 영향을 주는 기전으로는 에스트로젠 대사에 영향을 주는 β-glucuronidase 를 가지고 있는 장내미생물의 역할도 있지만 장내미생물의 분비물에 의하는 경우가 많음이 밝혀지고 있다(Fig. 10).92 최근 장내미생물의 분비물 중 가장 연구가 많이 되고 있는 SCFA 에는 아세트산(acetate), 프로피온산(propionate) 및 뷰티르산(butyrate) 이 있고 이들은 에너지원은 물론 염증조절자이기도 하고 장운동 및 상처 치유에 큰 역할을 하는 것으로 알려져 있으나 용종 형성을 증가시킨다는 보고도 있는 등 연구 결과가 좀 더 다양해지고 있다. SCFA 외에도 대장암 발생에 관여하는 장내미생물 분비물에는 heme 내지 2차 담즙산(secondary bile acid) 등이 있는데(Fig. 10),93 이들이 대장암발생에 미치는 매개하는 인자로는 장투과성 증가, 염증 증가,자가면역성 및 증식 증가로 요약할 수 있다(Fig. 11).93 대장암환자에서 Fusobacterium nucleatum이 증가한 경우가 흔히 관찰되지만 대장암의 원인인지 아니면 대장암 발생의 결과인지 확실히 밝혀진 바 없는 등 아직까지 인체에서 대장암을 유발하는 장내미생물이 밝혀진 바 없다. 또한 이들의 성차에 대한 보고는 더욱 없으나 향후 연구가 좀 더 세분화 되면서 대장암을 유발하는 장내미생물의 상이성이 대장암의 성차에 기여한다는 결과가 나올 수 있지 않을까 기대된다.
 | Fig. 10General mechanisms for microbiota-related colon cancer. Bacteria and their products affect the gut permeability to antigen. Dietary substrates undergo bacterial metabolism to form potentially carcinogenic products such as NOCs and secondary bile acids. Accordingly, chronic inflammation and genotoxicity contribute to the colon tumorigenesis. NOCs, N-nitroso compounds. Adapted from Yoon and Kim93 with permission.
|
 | Fig. 11Dietary compounds and the role of microbiota in colon carcinogenesis. Specific food compounds that are related to microbiota, and have a role in colon carcinogenesis, are shown. SCFAs, short chain fatty acids; NOCs, N-nitroso compounds; ROS, reactive oxygen species. Adapted from Yoon and Kim93 with permission.
|
Go to :
요 약
장내미생물은 식이, 인종, 나이, 항생제, 스트레스, 심리적요인, 산모의 건강, 자연분만 등의 출생 방법, 환경 요인과 운동 등에 영향을 받는다. 최근 장내미생물이 남녀 차이를 보이며 에스트로젠과 안드로젠과 같은 성호르몬의 역할이 부각되고 있다.50 예를 들어 폐경 전후의 장내미생물 조성이 다르고, 폐경이 되면 F/B의 비율이 높아지는데, 이는 남성보다 더 높았다. 성호르몬과 장내세균에 대한 관계에 대한 보고도 많은데 ERβ가 장내세균의 대사를 직접 조절하기도 하지만 역으로 장내세균이 에스트로젠 대사에 적극 개입함이 밝혀지고 있다. 즉 장내미생물 β-glucuronidase가 결합에스트로젠을 비결합형으로 변화시키면 이들이 장간순환으로 들어가서 에스트로젠 수용체 ERα 및 ERβ에 작용한다. 다만 이러한 현상은 여성폐경 전에는 나타나지 않아 비난소 에스트로젠의 경우에 국한되는 것으로 추정되고 있다. 최근 과민성 장증후군, 염증성장질환 및 대장암 등 여러 위장관 질환이 장내미생물과 관련이 있음이 밝혀지고 있는데, 이들은 다시 성호르몬과 밀접한 관련성이 있다. 하지만 각각에 대한 연구는 아직 미진한 바이에 대한 다양한 연구가 진행할 필요가 있다. 향후 성호르몬과 연계된 장내미생물 차이를 잘 연구함으로써 새로운 신약개발에 도움을 받을 수 있을 것으로 생각된다.
Go to :
 XML Download
XML Download