

 PDF
PDF Citation
Citation Print
Print



Introduction
The bony carotid canal in adults is a tube-like bone with a rough surface in contrast to smooth surfaces of the other parts of the temporal bone petrosal portion (below, abbreviated by “petrosa”): the morphology takes an impression of the additional, out-sourcing product after a complete development of the inner ear in the petrosa (Fig. 1). The slowly-developed tympanic bone also has a rough external surface. Without the carotid canal and tympanic bone, the anteroinferior aspect of the petrosa would simply reflect a slight convex of the cochlear wall. Our group incidentally demonstrated photos of the bony canal developing much later than any surface structures of the petrosa [1, 2] although it was out of their focus.
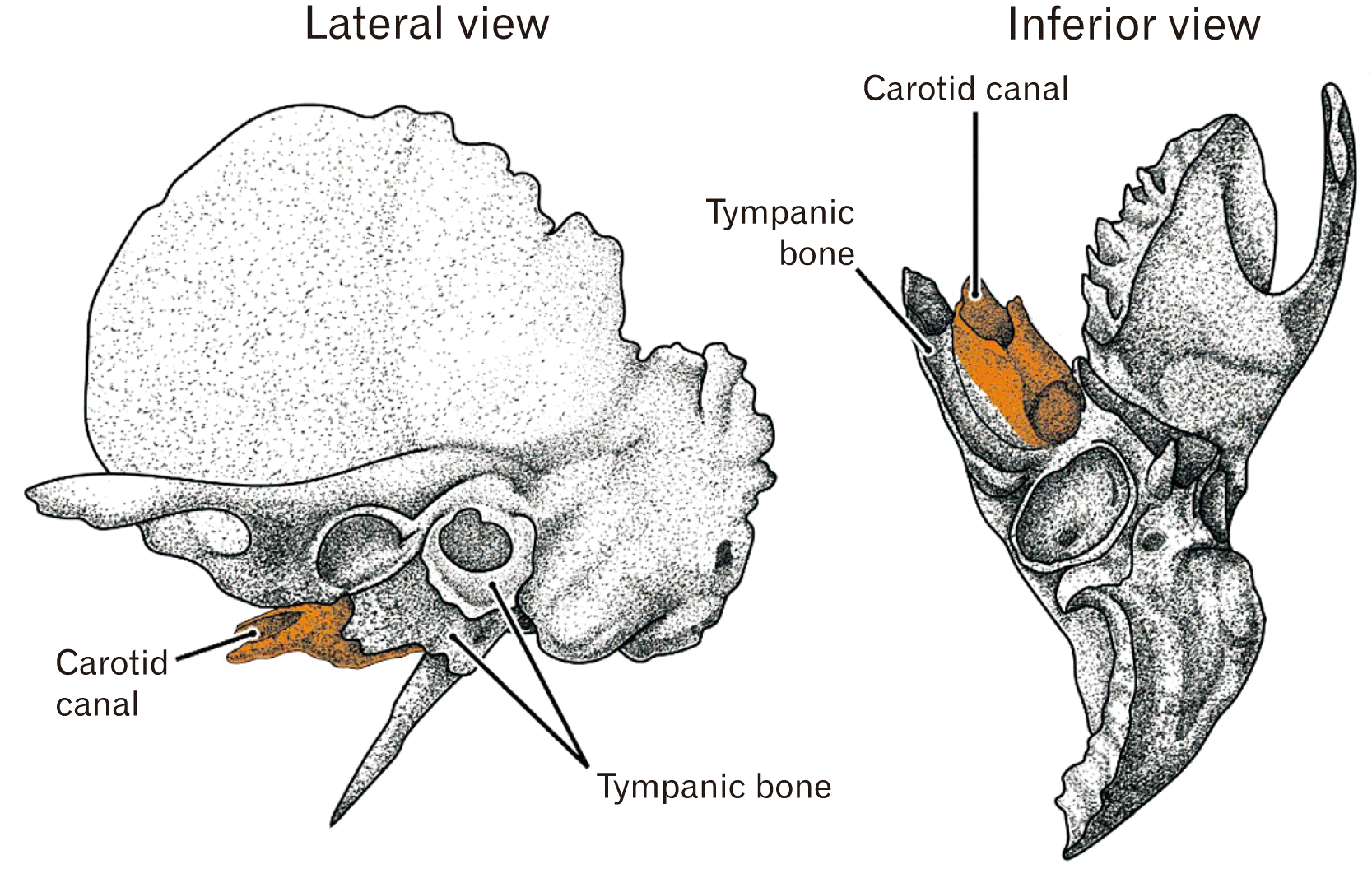
Usually, the skull base skeleton is simply divided into two parts: bony parts in which endochondral ossification occurs and the other parts with membranous ossification [3, 4]. However, in the skull base, as typically seen in the pterygoid of the sphenoid, membranous ossification simultaneously occurs at a site adjacent to endochondral ossification [5]. Also in the ear, Michaels et al. (2010) [6] described membranous ossification occurs to provide a lamellar bone along the endolymphatic duct after ossification of the otic capsule cartilage. Because endochondral ossification of the ear requires its cartilage model before midterm and occurs until 21–24 weeks [7-10], the carotid canal is most likely to be established by membranous ossification as in the facial nerve canal [11]. A delayed ossification is known well in the facial nerve canal because of the so-called dehiscence (reviewed by Spector and Ge [12], 1993).
Although the bony carotid canal may be simply believed as a part of the petrosa, no study had been conducted to evaluate a contribution of the adjacent sphenoid and pharyngotympanic tube in the fetal development and growth. In the anterior side of the ear, three elements of the sphenoid are located: 1) the ala temporalis and 2) the alar process, both of which take endochondral ossification and, 3) the pterygoid composed of multiple parts each of which takes endochondral or membranous ossification [5, 13, 14]. Is it likely for the sphenoid elements to contribute the closure of the carotid canal? Consequently, the aim of this study was to examine when and how the internal carotid artery (ICA) is involved into the petrosa without involvement into the sphenoid elements.
Go to : 
Materials and Methods
This study was conducted in accordance with the Declaration of Helsinki 1995 (as revised in 2013). We examined paraffin-embedded histological sections from 32 fetuses (approximately 12 to 37 weeks of gestational age; 28 to 310 mm of crown-rump length (CRL). The specimens were categorized into two groups according to age and size: 1) 20 midterm fetuses at 10 to 18 weeks of gestational age (CRL, 19–120 mm) and 2) 17 late-stage fetuses at 25 to 37 weeks (CRL, 200–310 mm).
Serial histological sections of 20 midterm fetuses were part of the large collection kept at the Department of Anatomy of the Complutense University of Madrid, Madrid, and were the results of miscarriages and ectopic pregnancies from the Department of Obstetrics of the University. They had been prepared serially (sagittal, 15 fetuses; horizontal, five fetuses) and stained with hematoxylin and eosin (H&E), azan or orange G. This study was approved by the Ethics Committee of Complutense University of Madrid (B08/374). Photographs were taken with a Nikon Eclipse 80 (Nikon, Tokyo, Japan).
From 17 late-stage fetuses, we prepared paraffin-embedded sections at 50 or 100 micron interval. The sectional planes were sagittal (12 fetuses) and horizontal (5 fetuses) and, all sections were stained with H&E. These specimens were parts of the collection of the Department of Anatomy, Akita University, Akita, Japan. These specimens had been donated by their families to the Department from 1975–1985 and preserved in 10% (w/w) neutral formalin solution for more than 30 years. The available data were limited to the date of donation and gestational age, but there was no information on family name, the name of the obstetrician or hospital and the reason for abortion. The use of these specimens for research was approved by the ethics committee of Akita University (No. 1428). Photographs were taken with a Nikon Eclipse 80.
Most photographs were taken with a Nikon Eclipse 80, whereas photographs at ultra-low magnification (objective lens less than ×1) were obtained using a high-grade flat scanner with translucent illumination (Epson scanner GTX970; Epson, Nagano, Japan).
Go to :
Results
Observations of the midterm fetuses
Sagittal sections from 15 midterm fetuses (12–18 weeks) always demonstrated a longitudinal course of the ICA along a slight curvature or convex provided by the cartilaginous anterior cochlear wall (Fig. 2). The ICA ran superiorly along the vertebral column and, near the inferior end of the inner ear cartilage, the ICA turned anteriorly. Thus, the anterior turn of the ICA at the skull base was established until midterm. The ICA passed through a wide gap or loose mesenchymal space between the cochlear wall and sphenoid. The anteroposterior length of the gap was almost same as the short diameter of the cochlear turn or spiral.
 | Fig. 2Initial carotid canal in sagittal sections: a wide gap or loose space between the cochlear wall and the sphenoid. Azan staining (A–D) or H&E staining (E, F). (A, B) A fetus of crown-rump length (CRL) 97 mm (approximately 13 weeks); (C, D) CRL 100 mm (13 weeks); (E, F) CRL 105 mm (14 weeks). (B) and (D) are higher magnification view of the center of (A) and (C), respectively. (F) is a higher magnification view of a square in (E). The left-hand side of each panel corresponds to the anterior side of the head. The ala temporalis of the sphenoid (ala) is ossified in (B) and (D), while it is cartilaginous in (E). In (D), an inferior part of the ala (ala with asterisk) looks like a pterygoid. In contrast, most parts of the pterygoid of the sphenoid is bony (B, E). (F) Shows the pterygoid containing two modes of the ossification. (B, D, E) Stars indicate a linear mesenchymal condensation corresponding to the future antero-inferior margin of the petrosal part of the temporal bone. Scale bars: (A, C) 5 mm; (B, D, E) 1 mm; (F) 0.1 mm. ABD, abducant nerve; AP, alar process; DPN, deep petrosal nerve; ICA, internal carotid artery; IR, inferior rectus of the eye; PPG, pterygopalatine ganglion; PTT, pharyngotympanic tube; tent, tentorium cerebelli; TG, trigeminal ganglion.
|
In the anterior side of the longitudinal course of the ICA, a single sagittal section was likely to contain three elements of the sphenoid: the ala temporalis, the alar process and the pterygoid (Fig. 2E). The pterygoid was always ossified except for the hamulus. However, whether the ala temporalis and alar process were cartilaginous or bony varied between specimens even in specimens of almost same sizes. Thus, three combinations were seen: 1) the ala and alar process were bony (Fig. 2B), 2) the alar process was bony and the ala temporalis was cartilaginous (Fig. 2D), 3) both were cartilaginous (Fig. 2E). The third pattern was seen in specimens of CRL 75 mm, 80 mm, and 105 mm. The cartilaginous ala looked like the pterygoid in shape (Fig. 2D). The pterygoid contained two types of ossification: 1) endochondral ossification that was characterized by balloon-like cartilage cells adjacent to eosinophilic calcified zone and 2) membranous ossification without cartilage cells at and near the calcified zone (Fig. 2F). Posterior parts of the pterygoid, those faced the ICA, took endochondral ossification.
Although it did not depend on sizes of fetuses, topographical anatomy of the pharyngotympanic tube (PTT) varied between specimens: the PTT was closely to or distant from the ala temporalis and ICA (Fig. 2B vs. Fig. 2D, E). Notably, a linear mesenchymal condensation, crossing the ICA course, extended between the PTT cartilage and the cochlear wall (Fig. 2B, D, E). Being different from the adult morphology, a posterolateral part of the trigeminal ganglion was attached to the cochlear wall (Fig. 2B, C).
According to horizontal sections from five midterm fetuses (Fig. 3), the ICA was surrounded by the petrosa or cochlear wall posteriorly, the alar process anteriorly and the basisphenoid (body of the sphenoid) medially. The basisphenoid is a large cartilaginous mass that contains both of the hypophysial fossa (Rathke’s pouch remnant) and the notochord rostral end [15, 16]. The basisphenoid connected to the petrosal cochlear wall. A slit between the alar process and patrosa became narrower in larger specimens (Fig. 3D vs. Fig. 3F). Likewise, the ICA course was also constricted by another slit of the sphenoid between the basisphenoid and the orbital cartilage (future ala minor) (Fig. 3A). As shown in sagittal sections, the trigeminal ganglion was attached to the cochlear wall (Fig. 3A). Taken together with observations of sagittal sections, at midterm, the ICA did not face areas of membranous ossification but it was completely surrounded by endochondral ossification sites.
 | Fig. 3Initial carotid canal in horizontal sections: a slit surrounded by the cochlear wall, alar process (AP) and basisphenoid. Azan staining. (A–C) A fetus of crown-rump length (CRL) 49 mm (approximately 10 weeks); (D) CRL 52 mm (10 weeks); (E) CRL 95 mm (13 weeks); (F) CRL 100 mm (14 weeks). The upper side of each panel corresponds to the anterior side of the head. The basisphenoid, a large cartilage mass, contains both the notochord (NC) and hypophysial fossa (fossa). The internal carotid artery is surrounded by the petrosa or cochlear wall posteriorly, the AP anteriorly and the basisphenoid medially. A slit between the petrosa and sphenoid appears to be narrower in larger specimens. All scale bars: 1 mm. chiasma, optic chiasma; HP, hypophysis; ICA, internal carotid artery; ON, optic nerve; SPS, superior petrosal sinus; TG, trigeminal ganglion.
|
Observations of the late-stage fetuses
We had an impression that the ICA consistently passed through a narrow gap between the bony alar process and the bony cochlear wall for entering a parasellar area of the future cavernous sinus (Figs. 4–6). However, this “narrow gap” had the almost same anteroposterior length as the short diameter of the cochlear turn or spiral. Thus, the gap appeared not to be narrower than that at midterm. The alar process and ala temporalis of the sphenoid became large in combination with the thickening cochlear wall. In contrast to the bony alar process, the ala temporalis still contained a large cartilage mass in the center (Figs. 4D, 5A, 6A), whereas the pterygoid was composed of multiple bony bars (Fig. 5A). Below the gap between the sphenoid and cochlear wall, the ICA was contained in a dense mesenchymal tissue of which inferior margin was connected between the sphenoid (the alar process, ala temporalis and/or pterygoid) and the anterior cochlear wall (Figs. 5A, B, 6D). In addition, as in midterm fetuses, the trigeminal ganglion was always attached to the cochlear wall (Figs. 4B, C, 5D, 6F).
 | Fig. 4Wide and dense mesenchymal tissue containing the internal carotid artery and extending between the cochlear wall and sphenoid. Horizontal sections. A fetus of crown-rump length (CRL) 228 mm (approximately 28 weeks). H&E staining. (A) or (D) displays the most superior (or inferior) site in the figure. (E–G) are higher magnification views of squares in (A), (B), and (D), respectively. The left-hand side of each panel corresponds to the anterior side of the head. In these superior sections, no structure delineates the future carotid canal except for the bony alar process (AP) of the sphenoid. Instead, a dense mesenchymal tissue extends between the cochlear wall and sphenoid and it contains abundant veins. (A–D) or (E–G) were prepared at the same magnification; (A, E) Scale bar: 1 mm. ala, ala temporalis of the sphenoid; GPN, greater petrosal nerve; ICA, internal carotid artery; ICN, internal carotid nerve; MXN, maxillary nerve (intracranial course); TG, trigeminal ganglion.
|
 | Fig. 5Initial ossification of the carotid canal starts in the dense mesenchymal tissue connecting to the anterosuperior end of the cochlear wall. Sagittal sections. A fetus of crown-rump length (CRL) 276 mm (approximately 32 weeks). H&E staining. Panel (A) containing the future cavernous sinus, displays the most medial site in the figure. (C) and (E) are higher magnification views of squares in (B) and (D), respectively. Panel (F) corresponding to a square in (E), exhibits the membranous ossification starts in the in the dense mesenchymal tissue connecting to the anterior end of the cochlear wall. The left-hand side of each panel corresponds to the anterior side of the head. Note an inferior margin (stars in A–E) of the dense mesenchymal tissue extends between the cochlear wall and the sphenoid (the alar process [AP], ala temporalis [ala] and pterygoid): it connects with the pharyngotympanic tube cartilage in (B) and (D). The palatine bone (PB) looks like a part of the pterygoid (A). Scale bars: (A–E) 1 mm; (F) 0.1 mm. ala, ala temporalis of the sphenoid; ABN, abducant nerve; ICA, internal carotid artery; IR, inferior rectus of the eye; MXN, maxillary nerve in the round foramen; OCN, oculomotor nerve; PPG, pterygopalatine ganglion; PTT, pharyngotympanic tube; SPS, superior petrosal sinus; TG, trigeminal ganglion.
|
 | Fig. 6Carotid canal is involved into the bony cochlear wall. Sagittal sections. A fetus of crown-rump length (CRL) 274 mm (approximately 31 weeks). H&E staining. The left-hand side of each panel corresponds to the superior side of the head. Panel (A) containing the future cavernous sinus, displays the most medial site in the figure. (B) or (C) is a higher magnification view of a square in (A) or (B). The inferior side of each panel corresponds to the anterior side of the head (long arrow in A). In the medial part of the carotid canal (A–C), a membranous ossification is on-going. In contrast, in the lateral part, a bony process of the cochlear wall (arrow in D, E) appears to surround the carotid canal from the inferior side rather than from the superior side (Fig. 3D). Stars in (D) indicate an inferior margin of the dense mesenchymal tissue (corresponding to that in Fig. 3A, B) extends between the cochlear wall and sphenoid (ala temporalis or ala). Asterisk in (F) indicates the medial end of the growing tympanic cavity. Scale bars: (A, B, D–F) 1 mm; (C) 0.1 mm. AP, alar process of the sphenoid; ICA, internal carotid artery; ICN, internal carotid nerve; MHT, meningohypophysial trunk of the internal carotid artery; OPN, ophthalmic nerve; PTT, pharyngotympanic tube; SPS, superior petrosal sinus; TG, trigeminal ganglion.
|
Irrespective of the fetus sizes, below the alar process of the sphenoid, a structure anterior to the ICA course was different between specimens and it was classified into three morphologies: 1) the dense mesenchymal tissue contained abundant veins (Fig. 4; CRL, 228 mm); 2) membranous ossification occurred in the dense mesenchymal tissue (Fig. 5; CRL, 276 mm) and; 3) a bony process became surrounding the ICA in contrast to the highly reduced size of dense mesenchymal tissue (Fig. 6; CRL, 274 mm). The dense mesenchymal tissue with ossification spots was connected with the cochlear wall rather than the sphenoid because the PTT interposed between the dense tissue and the sphenoid (Fig. 5D). The bony process surrounding the ICA was not originated from the sphenoid but the cochlear wall (Fig. 6B, E) and it continued to an anteromedial end of the tympanic cavity wall (Fig. 6F). Thus, conversely, the ICA became positioned in the supero-antero-lateral side of the tympanic cavity (Fig. 7A, B). A fatty tissue occupied a large loose space below the dense mesenchymal tissue (Fig. 5A), i.e., the prestyloid compartment of the parapharyngeal space [17, 18] and it also contained muscles from the stylopharyngeus and levator veli palatini muscles.
 | Fig. 7Carotid and facial nerve canals adjacent to the tympanic cavity: membranous ossification in these septa. Sagittal sections. A fetus of crown-rump length (CRL) 272 mm (approximately 31 weeks). H&E staining. The left-hand side of each panel corresponds to the posterior side of the head. Panel (A) displays a plane 1.2 mm medial to (B). (A, B) Contain the carotid and facial nerve canals: the septum is developing for the internal carotid artery (ICA), while the latter canal still opens widely to the tympanic cavity (arrows in B). (C) or (E) is a higher magnification view of a square in panel (A) or (D). The left-hand side of each panel corresponds to the anterior side of the head. Near the facial nerve, membranous ossification (upper insert of C) coexist endochondral ossification indicated by a tidemark (arrowheads; lower insert). (D) A section near (B), exhibits a site corresponding to a square in (B). (E) A higher magnification view of a square in (D), shows membranous ossification in a septum between the carotid canal and tympanic cavity. Scale bars: (A–D) 1 mm; (E) 0.1 mm. Magnification of inserts, same as (E). ant, anterior; infe, inferior; ICN, internal carotid nerve; LSC, lateral semicircular canal.
|
Even after the anterior cochlear wall involved the ICA inside (the aforementioned, the third morphology), a septum was underdeveloped between the tympanic cavity and the future carotid canal: membranous ossification was on-going in the septum (Figs. 6F, 7D, E). Likewise, a separation of the facial nerve from the tympanic cavity by a membranous bone was always incomplete (Fig. 7C), i.e., the so-called facial canal dehiscence (see the Introduction). In contrast to these membranous bones, a tidemark (a border between the calcified and uncalcified bones) was sometimes seen in bones near and along the membranous labyrinth (two inserts in Fig. 7C).
Go to :
Discussion
The present study demonstrated that membranous ossification occurred in a dense mesenchymal tissue along the ICA. This mother tissue of the bony carotid canal, containing abundant veins, was delineated anteriorly and posteriorly by the cartilaginous petrosa and sphenoid, the latter was composed of the basisphenoid medially, the alar process supero-anteriorly, the ala temporalis infero-anteriorly. The pterygoid of the sphenoid also makes an infero-anterior margin. Notably, all these sphenoid elements facing the ICA took endochondral ossification. Thus, any sites of membranous ossification did not face the mother tissue of the bony carotid canal. Therefore, at the initial stage, membranous ossification along the ICA seemed to be likely in both or either mesenchymal tissues derived from the petrosa and sphenoid. The ICA comes from the parapharyngeal space far posterior to the sphenoid [17, 18]. The ICA seemed to be pulled posteriorly at the cranial base: this traction might be one reason of no or little contribution of a mesenchymal tissue from the sphenoid for the carotid canal ossification.
A narrow gap between the alar process and cochlear wall early determined the superior end of the future bony carotid canal. However, not the alar process but the basisphenoid contributes to a connection between the petrosa and sphenoid. Being different from an imagination suggested by the name, the ala temporalis of the sphenoid does not connect with the petrosa but builds a part of the skull base for a base of the inferiorly extending pterygoid. Inferiorly, the mother tissue of the bony canal was delineated by the PTT cartilage at midterm. The PTT cartilage appears late and finally in all cartilages of the body [1]. Near term, the growing pterygoid pushed the PTT upward and, in turn, the PTT pushed the ICA upward toward the petrosa. The pterygoid enlarged to provide large attachments of pterygoid muscles. Simultaneously, the PTT separated the mother tissue of the bony canal from the pterygoid. Therefore, the newly-built membranous bone along the ICA was liable to attach to the petrosa, not to the sphenoid. Depending on involvement of the PTT into the bony tympanic cavity (or depending on growth of the tympanic bone), the ICA was also involved into the petrosa. The bony carotid canal developed without a clear correlation with size of fetuses after 25 weeks. The final process, ossification of the septum between the carotid canal and tympanic cavity, was much delayed and possibly finished after birth.
At any stage examined, the trigeminal ganglion attached to the cochleal wall in contrast to the adult morphology and the ICA did not show the siphon-like course alongside the sphenoid. The former morphology suggested a drastic topographical change or an anterior sliding of the ganglion after birth. However, magnetic resonance imaging of fetuses and infants did not state about the change [19-22]. The latter evoked a question as to when and how multiple turns or a siphon-like course of the ICA is established: 1) an anteromedial turn at the base of the petrosa (an external aperture of the carotid canal); 2) an anterior bending at the entrance to the cavernous sinus; 3) a strong posterior bending in the sinus; 4) a superior turn at the exit from the cavernous sinus. However, Weninger and Müller [23, 24] demonstrated that, in infants, the intracavernous ICA shows an individual and age-dependent variations from the almost straight course (25%) to an adult-like siphon. Our group recently ensured the straight course in near-term fetuses [25].
Go to :
 XML Download
XML Download