

 PDF
PDF Citation
Citation Print
Print



Introduction
Acquired or inherited photoreceptors loss initiates a chain of events, known as “retinal remodeling”, that ends in retinal ganglion cell (RGC) death [1]. As RGCs are the most relevant neurons in the inner retina as their axons form the optic nerve that sends the visual information to the visual cortex. Thus, therapies should be applied early during photoreceptors degeneration before the remodeling process reaches the inner retina [1].
N-Ethyl-N-nitrosourea (ENU) is a DNA alkylating agent that targets the retinal photoreceptor cells. It has been used to create photoreceptor degenerative diseases animal model via apoptosis [2], such as age-related macular degeneration and retinitis pigmentosa (RP) [3, 4].
Metformin is a well-tested drug used to treat type 2 diabetes, and recently it has been reported to have beneficial effects in neurodegenerative diseases [5]. It was demonstrated that metformin ameliorates the neuropathic effects of acrylamide in rats by decreasing caspase3 activity and up regulating bcl2 expression [6].
So, this study was carried out to evaluate the protective effect of metformin on the rat optic nerve following photoreceptors loss induced by ENU using light and electron microscope.
Materials and Methods
A total number of 18 adult 5 months old male Wistar rats (200 g body weight) were used in this study. They were purchased from the Central Animal House, Faculty of Medicine, Assiut University. All animal procedures were in accordance with the standards set forth in the guidelines for the care and use of experimental animals by the Committee for the Purpose of Supervision of Experiments on Animals and according to the National Institute of Health protocol and approved by the Institutional Ethics Committee of Assiut University.
Animal groups: The animals were divided into 2 groups
Group (I): (normal vehicle control) consisted of 6 rats that received a single intraperitoneal injection of 0.5 ml physiologic saline. Group (II): (ENU-induced photoreceptor degeneration) consisted of 12 rats that received a single intraperitoneal injection of ENU (purchased from Sigma Chemical Co., St. Louis, Mo, USA) at a dose of 600 mg/kg freshly dissolved in 0.5 ml physiologic saline [7]. Rats in group II were equally divided into two subgroups: Group (IIa) (photoreceptor degeneration group) consisted of 6 rats that received a daily intraperitoneal injection of 0.5 ml physiologic saline, for 7 days starting one day before ENU administration. Group (IIb) (metformin-treated group) consisted of 6 rats that received metformin (purchased from Nasr Company for Pharmaceutical Chemicals) by intraperitoneal injection once daily at a dose of 200 mg/kg dissolved in 0.5 ml physiologic saline, for 7 days. Metformin injection was started one day before ENU administration [8, 9].
The animals were anaesthetized with ether, their hearts were exposed, and then perfusion was done.
For a light microscope, three rats from each animal group were used and perfused intracardially with a 10% formaldehyde solution. The retina was separated, and the optic nerve (intraorbital part) was transected 1 to 2 mm behind the globe. Paraffin sections (5 μm) were cut using a microtome (Leica RM 2125RT, Leica microsystems, Wetzlar, Germany) and every 10th section was stained with hematoxylin-eosin [10]. Some sections were stained with Mallory’s phosphotungstic acid hematoxylin (PTAH) for demonstration of glial fibrils within astrocytes in the optic nerve [10].
For electron microscope, three rats from each group were perfused intracardially with 4% glutaraldehyde in cacodylate buffer (pH 7.4). Specimens from the retina and optic nerve were cut into thin slices (1×1 mm) and immersed in the fixative for 24 hours and post fixed in 1% osmium tetroxide in phosphate buffer for two hours [11]. Semi-thin sections (0.5–1 μm) were cut on the ultramicrotome (LKB Bromma 8800 UltratomeR III, 3518, Sweden) and stained with 1% toluidine blue (pH 7.3) for examination on a light microscope (Bx50. Model Bx50F-3, SC09160; Olympus, Tokyo, Japan). Ultrathin sections (50–80 nm) were cut from selected areas of the blocks on a Reichert ultramicrotome (Leica WILDM3Z, 89386, Wien, Austria) and contrasted with uranyl acetate and lead citrate. These sections were examined using the transmission electron microscope (Jeol E.M.-100 CX11; Japanese electron optic laboratory, Tokyo, Japan). All processes were done in Electron microscopic unit at Assiut University.
Immunohistochemical study
Expression of Caspase-3 (apoptotic marker), CD68 (macrophage marker), and inducible nitric oxide synthase (iNOS) (oxidative stress marker) was detected in formalin-fixed paraffin-embedded sections. The Caspase-3 (CPP32) Ab-4, Rabbit Polyclonal Antibody (Thermo Fisher Scientific, Fremont, CA 94538-7310, USA), the (iNOS) Rabbit Polyclonal Antibody (Thermo Fisher Scientific, Fremont, CA 94538-6406, USA), and the CD68 (Monoclonal mouse antibody) Ab-3, (Clone KP1 Lab Vision Co., Neo Markers Inc/Lab Vision, Fremont, CA, USA) were used at 1:100 dilution for 30 minutes at room temperature.
Sections were processed according to the manufacture instructions using the universal kit (Ecno Tek HRP Anti-Polyvalent, DAB) (ScyTek Laboratories, Inc., 205 South 600 West Logan, UT 84321 USA). After completion of the reaction, counterstaining was done using Mayer’s hematoxylin, dehydrated and cover-slipped using DPX (Oxford laboratory reagents, Bombay, India).
Morphometric study
Using the Image J software program, the total retinal thickness (from the internal limiting membrane to the pigment epithelium) was measured in 5 micrographs of toluidine blue stained semithin sections (×400 magnification) from three different animals for each group. The number of astrocytes per mm2 was counted in optic nerve in 5 micrographs of PTAH-stained paraffin sections (×400 magnification) from three different animals for each group. The number of Caspase-3, iNOS and CD68 positive cells per mm2 was counted in optic nerve in 5 micrographs of immunostained paraffin sections (×400 magnification) from three different animals for each group. The diameter of optic nerve fibers (μm) and thickness of the myelin sheath of optic nerve fibers (μm) were measures in 5 micrographs of ultrathin sections (×3,600 magnification) from three different animals for each group.
Statistical analysis
The morphometric data of each animal group were statistically analyzed using the computer statistics Prism-5.0 package (GraphPad Software, Inc., San Diego, CA, USA). One-way analysis of variance (ANOVA) followed by Turkey’s test as a post-test was employed to compare the studied animal group. The results were expressed as mean±standard deviation (SD). A P-value<0.05 was considered significant.
Results
Light microscopy
Histological results
In group I, the retinal layers were well defined (Fig. 1A). The mean total retinal thickness was 232.53±40.81 μm, ranged from 200.8 to 286.0 μm (Fig. 1D). The optic nerve was surrounded by the pia mater and divided into fascicles by thin connective tissue septa (Fig. 2A). It is formed of RGCs axons and glial cells. The axons appeared myelinated with variable sizes (Fig. 2D). Oligodendrocytes had small dark nuclei while astrocytes were characterized by large lightly stained nuclei (Fig. 2D). Astrocytes and their long processes appeared dark blue in color with PTAH stain (Fig. 2G). Their mean number per mm2 was 16±2.943, ranged from 13 to 20 (Fig. 2J).
In group IIa, ENU treated retina revealed photoreceptor degenerative changes in the form of vacuolations and loss of photoreceptor nuclei. Also, some ganglion cells were lost leaving empty spaces (Fig. 1B). The mean total retinal thickness was significantly decreased (P-value<0.000) compared to group I. It was 121.30±11.10 μm, ranged from 106.4 to 131.3 μm (Fig. 1D). The optic nerve showed less organized connective tissue septa (Fig. 2B). There was a reduction in the myelinated nerve axons compared to group I (Fig. 2E). Using PTAH stain, the optic nerve showed astrocytes with negatively stained processes (Fig. 2H). Their mean number per mm2 was significantly decreased (P-value<0.0003) compared to group I. It was 5.75±1.707, ranged from 4 to 8 (Fig. 2J).
In group IIb, metformin treated retina revealed decreased evidence of photoreceptor degenerative changes induced by ENU (Fig. 1C). The mean total retinal thickness was significantly increased (P-value<0.000) compared to group IIa. It was 195.22±9.19, ranged from 186.4–208.8 (Fig. 1D). The optic nerve connective tissue septa were still less organized compared to group I (Fig. 2C). The myelinated nerve axons were increased compared to group IIa (Fig. 2F). Astrocytic cell bodies and processes appeared dark blue in color with PTAH (Fig. 2I). Their mean number per mm2 was significantly increased (P-value<0.0007) compared to group IIa. It was 14.75±1.707, ranged from 13 to 17 (Fig. 2J).
Immunohistochemical results
Group I optic nerve showed mild positive caspase-3 reaction (Fig. 3A). The mean caspase positive cell number per mm2 was 1±0.8165, ranged from 0 to 2 (Fig. 3D). While in group IIa, caspase-3 expression was strong positive in glial cells (Fig. 3B). The mean caspase positive cell number per mm2 was significantly increased (P-value<0.0027) compared to group I. It was 5±1.826, ranged from 3 to 7 (Fig. 3D). The optic nerve of group IIb showed decreased caspase-3 immunoreactivity compared to group IIa (Fig. 3C). The mean caspase positive cell number per mm2 was significantly decreased (P-value<0.0097) compared to group IIa. It was 1.75±0.5, ranged from 1 to 2 (Fig. 3D).
The expression of iNOS immunoreactivity was mildly positive in group I optic nerve (Fig. 4A). The mean iNOS positive cell number per mm2 was 7±1.0, ranged from 6 to 8 (Fig. 4D). While group IIa showed strong positive iNOS reaction in glial cells (Fig. 4B). The mean number of iNOS positive cells per mm2 was significantly increased (P-value<0.0004) compared to group I. It was 20.3±2.5166, ranged from 18 to 23 (Fig. 4D). Group IIb optic nerve showed decreased iNOS immunoreactivity compared to group IIa (Fig. 4C). The mean number of iNOS positive cells per mm2 was significantly decreased (P-value<0.0004) compared to group IIa. It was 6.7±2.0816, ranged from 5 to 9 (Fig. 4D).
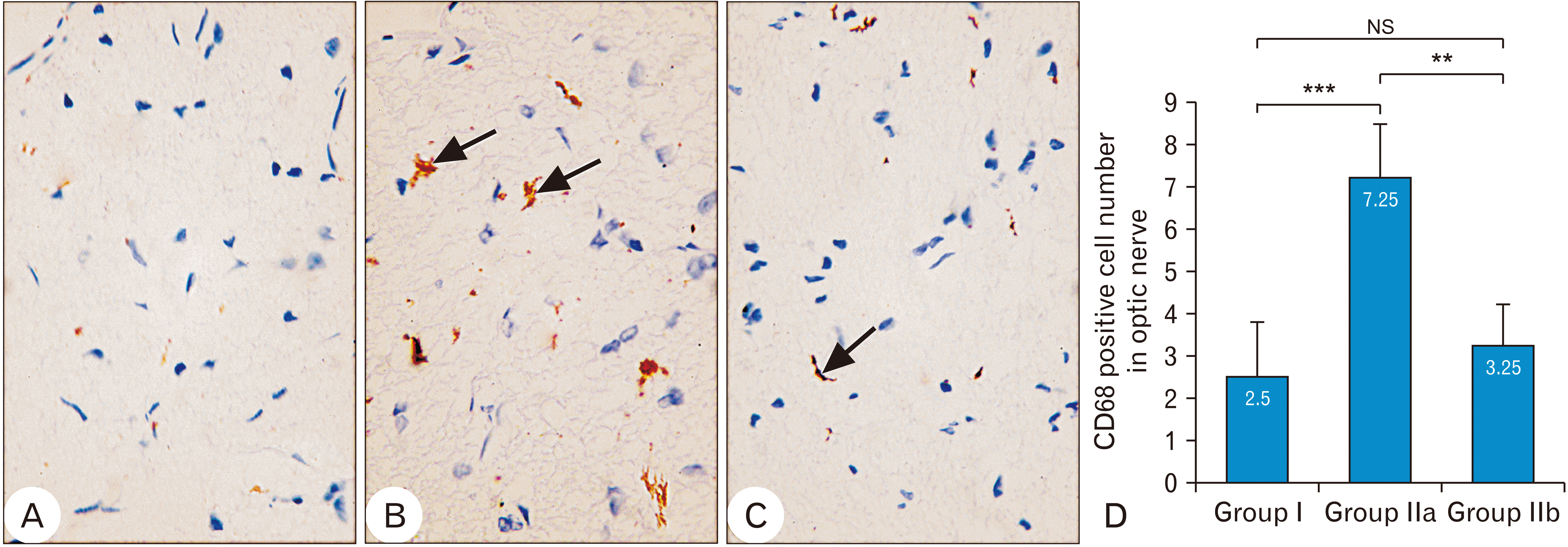
CD68 immunoreactivity was mild positive in group I optic nerve (Fig. 5A). The mean CD68 positive cell number per mm2 was 2.5±1.290994, ranged from 1 to 4 (Fig. 5D). While in group IIa, the optic nerve showed strong positive CD68 reaction in glial cells (Fig. 5B). The mean number of CD68 positive cells per mm2 was significantly increased (P-value<0.0008) compared to group I. It was 7.25±1.258306, ranged from 6 to 9 (Fig. 5D). The optic nerve of group IIb showed decreased CD68 immunoreactivity compared to group IIa (Fig. 5C). The mean number of CD68 positive cells per mm2 was significantly decreased (P-value<0.0025) compared to group IIa. It was 3.25±0.957427, ranged from 2 to 4 (Fig. 5D).
Electron microscopy
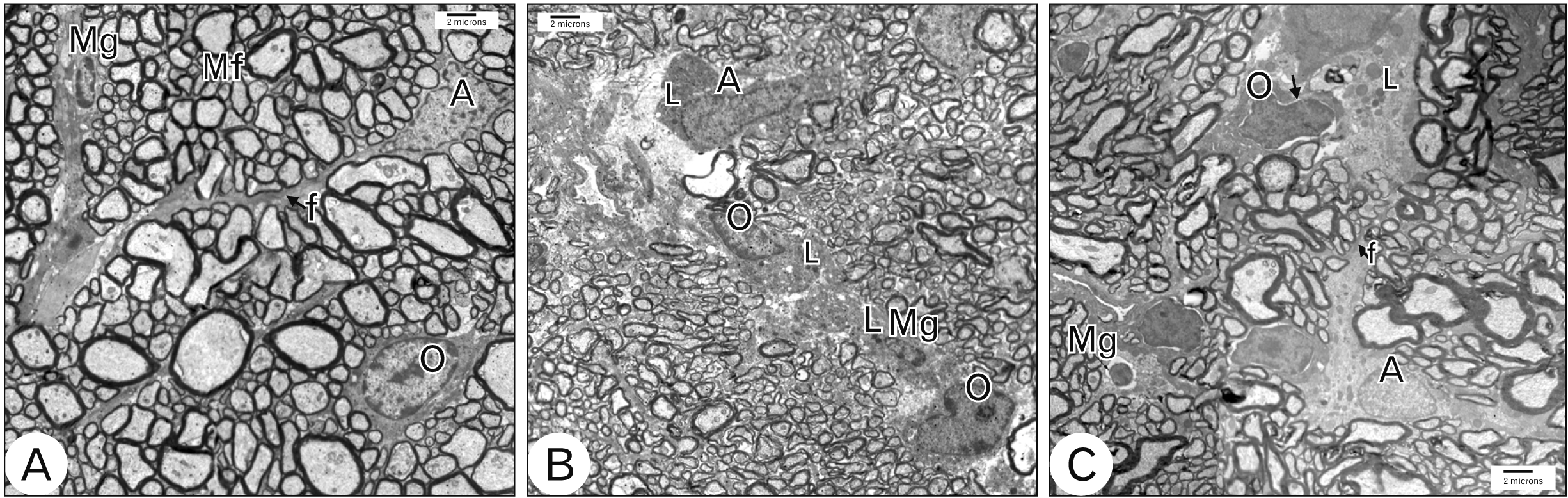
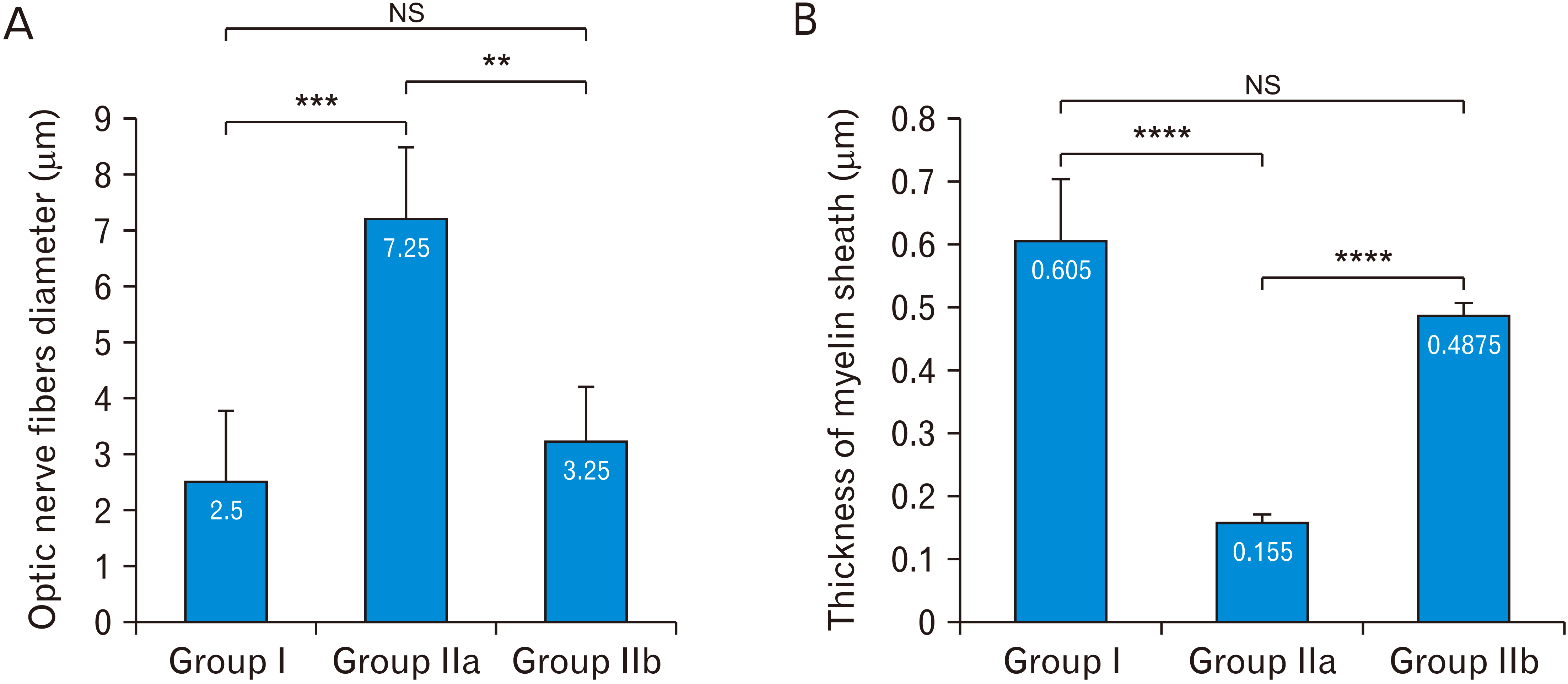
In group I, the myelinated optic nerve fibers were arranged in fascicles surrounded by astrocytic processes (Fig. 6A). The mean diameter of optic nerve fibers was 4.825±0.81802, ranged from 3.8 to 5.7 (Fig. 7A). The mean thickness of myelin sheath of optic nerve fibers was 0.605±0.099, ranged from 0.53 to 0.75 (Fig. 7B). Astrocytes had large euchromatic nucleus with thin peripheral heterochromatin and long processes containing bundles of filaments (Fig. 6A). Oligodendrocytes had ovoid nucleus with peripheral clumped chromatin and moderate electron-dense cytoplasm (Fig. 6A). Microglia had small oval heterochromatic nucleus (Fig. 6A).
In group IIa, there was a reduction in the diameter of the optic nerve fibers compared to group I with thinning of myelin sheath. The mean diameter of optic nerve fibers was significantly decreased (P-value<0.0002) compared to group I. It was 1.625±0.15, ranged from 1.5 to 1.8 (Fig. 7A). The mean thickness of myelin sheath of optic nerve fibers was significantly decreased (P-value<0.0001) compared to group I. It was 0.155±0.013, ranged from 0.14 to 0.17 (Fig. 7B). Astrocytes were more electron-dense compared to group I with disrupted processes and their cytoplasm contained numerous lysosomes (Fig. 6B). Oligodendrocytes were more electron-dense compared to group I and their cytoplasm contained numerous lysosomes (Fig. 6B). Activated microglia with numerous lysosomes were observed (Fig. 6B).
In group IIb, there was an increase in the diameter of the optic nerve fibers compared to group IIa. The mean diameter of optic nerve fibers was significantly increased (P-value<0.0012) compared to group IIa. It was 4.1575±0.8045, ranged from 3.03 to 4.9 (Fig. 7A). The mean thickness of myelin sheath of optic nerve fibers was significantly increased (P-value<0.0001) compared to group IIa. It was 0.4875±0.021, ranged from 0.46 to 0.51 (Fig. 7B). Astrocytes were less electron-dense compared to group IIa. They had large nucleus with thin peripheral heterochromatin, long processes containing bundles of filaments and lysosomes (Fig. 6C). Oligodendrocytes were electron dense with dilated perinuclear cisternae (Fig. 6C). Less activated microglia with small oval heterochromatic nucleus and occasional lysosomes (Fig. 6C).
Discussion
In this work, we evaluated the protective effect of metformin on the rat optic nerve following photoreceptors loss induced by ENU. Photoreceptor loss initiates retinal remodeling, which ends in RGC death, the neurons that transmit the information to the brain via the optic nerve [1]. It was reported that ENU induced apoptosis of the photoreceptors in rodents 7 days after a single intraperitoneal injection via the downregulation of Bcl-2, upregulation of Bax and activation of the caspase families [7, 12]. The retinal degenerative changes following photoreceptors loss induced by ENU will be discussed in another paper (in progress).
Group IIa (ENU-induced photoreceptor degeneration) of this study revealed a reduction in the diameter of the optic nerve fibers with thinning of myelin sheath confirmed by morphometric analysis. In agreement, axonal and myelin damage in optic nerve was detected in patients with inherited photoreceptor loss [13-15] Also, Similar findings were reported in transgenic rabbit model of inherited photoreceptor loss (RP) by Lahav et al. [16] and Asakawa et al. [17]. These authors hypothesized that the degenerative changes of optic nerve occurred as consequence to outer retinal degeneration.
On the other hand, statistical analysis of group IIb (metformin treated group) of this study revealed preservation of the diameter of optic nerve fibers and thickness of myelin sheath. This could correlate to the decreased oxidative stress and apoptosis detected by immunohistochemical study of iNOS and caspase-3 expression. In agreement, Oda [6] reported that metformin could protect against central and peripheral neuropathy in rat by decreasing oxidative stress and apoptosis via activation of adenosine monophosphate-activated protein kinase (AMPK) pathway. Also, Xu et al. [18] demonstrated that metformin could protect the optic nerve fibers by upregulation of crystallin proteins that enhance the survival of RGCs in transgenic rd1 mice (a model of photoreceptor degeneration).
The changes observed in optic nerve fibers in group IIa of this study were concomitant with morphological changes in oligodendrocytes suggested an impaired oligodendrocyte function. The progressive oligodendrocyte stress is followed by demyelination of the optic nerve [19, 20]. It was suggested that myelin repair could be performed by metformin via up-regulation of factors that promote oligodendrocyte precursor cells differentiation and maturation, recruitment of oligodendrocyte precursor cells to the lesion site and induction of their maturation and down-regulation of myelin-associated neurite outgrowth inhibitor marker [21]. Also, Metformin attenuated oxidative stress with corresponding induction of antioxidative defenses in oligodendrocytes exposed to cytokines via AMPK activation [5, 22, 23]. Moreover, Kim et al. [24] reported that αB-crystallin protein was expressed in peripapillary glial cells and some oligodendrocytes. The crystallin proteins which might be upregulated by metformin were proved to be protective in nerve injury [25].
Activated microglial cells, characterized by small heterochromatic nucleus and scanty cytoplasm containing numerous lysosomes, were observed in group IIa. In addition, the number of CD68 positive cells was markedly increased in this group. It is known that CD68 marker is associated with phagocytic microglia [26]. In agreement, Gupta et al. [27] suggested that microglia are activated in response to photoreceptor death. In addition, immunohistochemical and morphometric results of this study revealed increased iNOS expression in the optic nerve of group IIa. It is known that phagocytic microglia (also known as M1 microglia) release proinflammatory cytokines such as tumour necrosis factor alpha (TNF-α) and nitric oxide synthase‐2 (NOS-2). TNF‐α binds to cell membrane and triggers apoptotic pathways while NOS‐2 induces oxidative stress leading to caspase activation and mitochondrial dysfunction resulting in cell death [28, 29].
On the other hand, less activated microglial cells were observed in metformin-treated group of this study (group IIb). Statistically, the number of CD68 positive cells was significantly decreased compared to group IIa. There was an evidence that metformin inhibits microglia reactivity through different mechanisms including AMPK signaling, thereby reducing typical features of pro-inflammatory microglia including NADPH-dependent production of free radicals leading to polarization of microglia from proinflammatory phenotype (M1) to neuroprotective phenotype (M2) [30, 31].
Statistically, the number of astrocytes was decreased in group IIa optic nerve. Moreover, astrocytic processes were negatively stained with PTAH. The increased iNOS expression observed in this group indicated oxidative stress which might involve in the death of astrocytes [32]. Astrocytic clasmatodendrosis (lose their processes) was also observed in a previous study by Eltony and Abdelhameed [33] in optic nerve of Viagra treated rats. It was suggested that astrocytic clasmatodendrosis is related to excessive acidosis concomitant with hyperglycemia [34]. The hyperglycemia induced by ENU is thought to result from mutations in genes involved in glucose homeostasis [35, 36].
It is known that the optic nerve is subdivided into fascicles by connective tissue and glial septa [37]. The less organized septa observed in group IIa of this study was reported also by Asakawa et al. [17] in rabbit with photoreceptor degeneration. They attributed this disorganization to the enlargement of interstitial spaces due to the degeneration of myelinated axons.
Ultrastructurally, astrocytes of group IIa contained numerous lysosomes and lipofuscin granules. It has been reported that astrocytes participate in phagocytosis of cerebral white matter myelin in adult rabbit [38], in Wallerian degeneration of the opossum’s optic nerve [39] and following brain trauma [40].
On the other hand, this study revealed that there was no statistically significant difference in the mean number of PTAH-stained astrocytes between metformin-treated group and the control group. Metformin was found to inhibit caspase-3 through activation of AMPK pathway. Thus, protect astrocytes from apoptosis induced by stress stimuli [41, 42].
In conclusion, metformin ameliorates the histological changes of the rat optic nerve following photoreceptors loss induced by ENU. This suggests the protective role of metformin in preventing optic nerve damage secondary to retinal photoreceptor loss.



 XML Download
XML Download