

 PDF
PDF Citation
Citation Print
Print



Introduction
The definition of consciousness has evolved differently in the philosophy, psychology, and medicine fields [1]. Locke [2] defined consciousness as “the perception of what passes in a man’s own mind” in a modern conceptual style. Philosophers apply the term to four main topics (Routledge Encyclopedia of Philosophy, 1998): knowledge in general, intentionality, introspection and phenomenal experience. Perceptual experiences, such as tasting and seeing; bodily-sensational experiences, such as pains, tickles, and itches; imaginative experiences, like those of one’s own actions or perceptions; and streams of thought, as in the experience of thinking ‘in words’ or ‘in images’ are typical examples of phenomenal consciousness. According to the classical psychoanalytic theory of Sigmund Freud (APA Dictionary of Psychology, 2020), the region of the psyche contains thoughts, feelings, perceptions, and other aspects of mental life present in awareness. In medicine, the approach to consciousness is pragmatic and involves the treatment of individuals whose brain functions have been damaged by diseases, accidents, or drugs, and the determination of level consciousness by neurological examination [3, 4]. In this review, we define consciousness simply as “an organism’s awareness of something either internal or external to itself” and explore the softer question what brain areas and processes are associated with the emergence and maintenance of consciousness.
Consciousness has two-dimensions: awareness and wakefulness [4-6]. Awareness of consciousness refers to contents of consciousness such as sensory perception, self-reference, emotion, motivation, and social and goal-directed behaviors. Contents of the awareness can be further divided into two components: external vs internal awareness. External awareness involves perceptions formed from sensory information gleaned from the external world (i.e., what we see, hear, smell, and taste) and information derived from visual, auditory, somatosensory-motor, and attention networks. On the other hand, internal awareness involves stimulus-independent processes such as self-referential, emotional, motivational, social, and goal-directed behaviors. The default mode network (DMN) is mainly related to internal awareness.
Wakefulness is the state of vigilance or alertness. The separation of these dimensions is apparent in unresponsive wakefulness state (UWS) patients [7] and spatial neglect syndrome patients [8]. UWS patients show some signs of wakefulness but no sign of awareness—other than by neuroimaging, and spatial neglect syndrome patients show no conscious awareness of visual stimuli though they are awake. Although these two dimensions are different, neural processes associated with both have been shown to interact. The interaction between wakefulness processes, which are regulated by vertical connectivity (brain stem-thalamus-cortex), and awareness processes, which are regulated by horizontal connectivity (cortico-cortical communication), is introduced briefly at the end of this review.
Three components are required for conscious processes: structural connectivity, functional connectivity, and dynamic communications between these components [9]. Structural connectivity means physical connectivity, which can be revealed by neuroanatomical methods such as neural tracing and neuro-imaging methods like diffusion tensor imaging. Functional connectivity means a transient organization of clusters for the computation of specific psychological or cognitive processes. Although brain regions are anatomically connected (the connectome), all regions cannot be connected simultaneously for specific psychological or cognitive processes. Nonetheless, specific functional brain networks play critical roles in conscious processes. In this review, we mainly focus on functional connectivity.
Although neuroanatomical and neurophysiological approaches have added much to our understanding of the awareness of consciousness, recent progress in this field has been made mainly using neuroimaging techniques such as functional magnetic resonance imaging (fMRI) and PET. Together with graph theory-based analysis of neuro-imaging data, these techniques enable the brain mapping of many psychological and cognitive processes. Blood oxygen level dependent (BOLD) scans can be acquired by fMRI, and by using BOLD signals, the co-activation pattern of regions of interest (ROI) (path lengths and clustering coefficients) are calculated by graph theory analysis. The reservoir of fMRI data has been expanded; more than 3,800 functional neuroimaging papers were published in 2020 alone. Interestingly, a new software package called NeuroSynth (https://neurosynth.org/, Supplementary Information) has been developed for the large-scale collation and synthesis of human neuroimaging data [10]. This technique combines text-mining, meta-analysis, and machine-learning techniques to generate probabilistic mappings between cognitive and neural states. In this review, we mainly focus on progress made using neuro-imaging techniques but also address progress in the neuroanatomical and neurophysiological fields. In addition, we present the results of meta-analysis using the NeuroSynth web tool (Figs. 1 and 2).
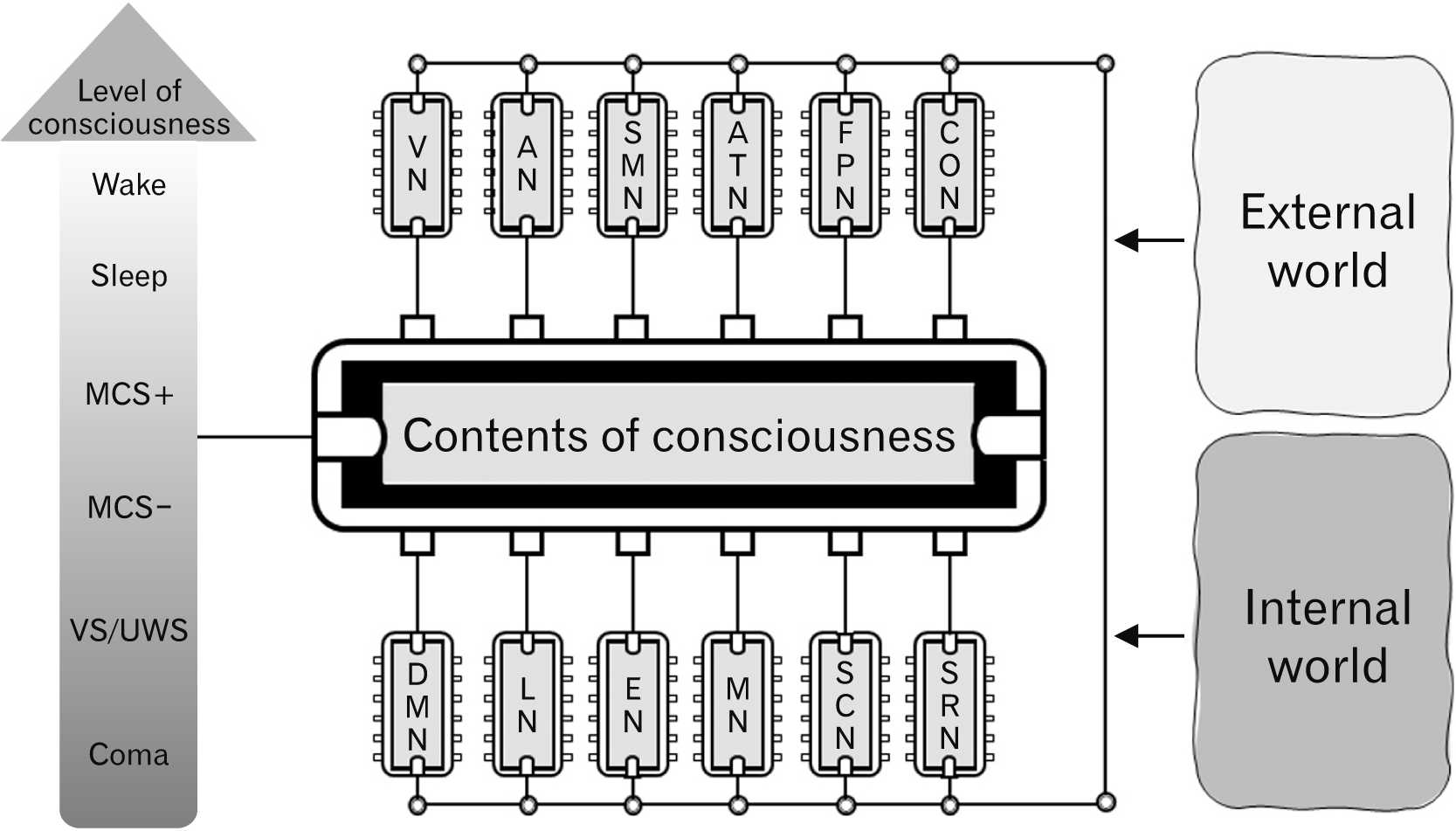
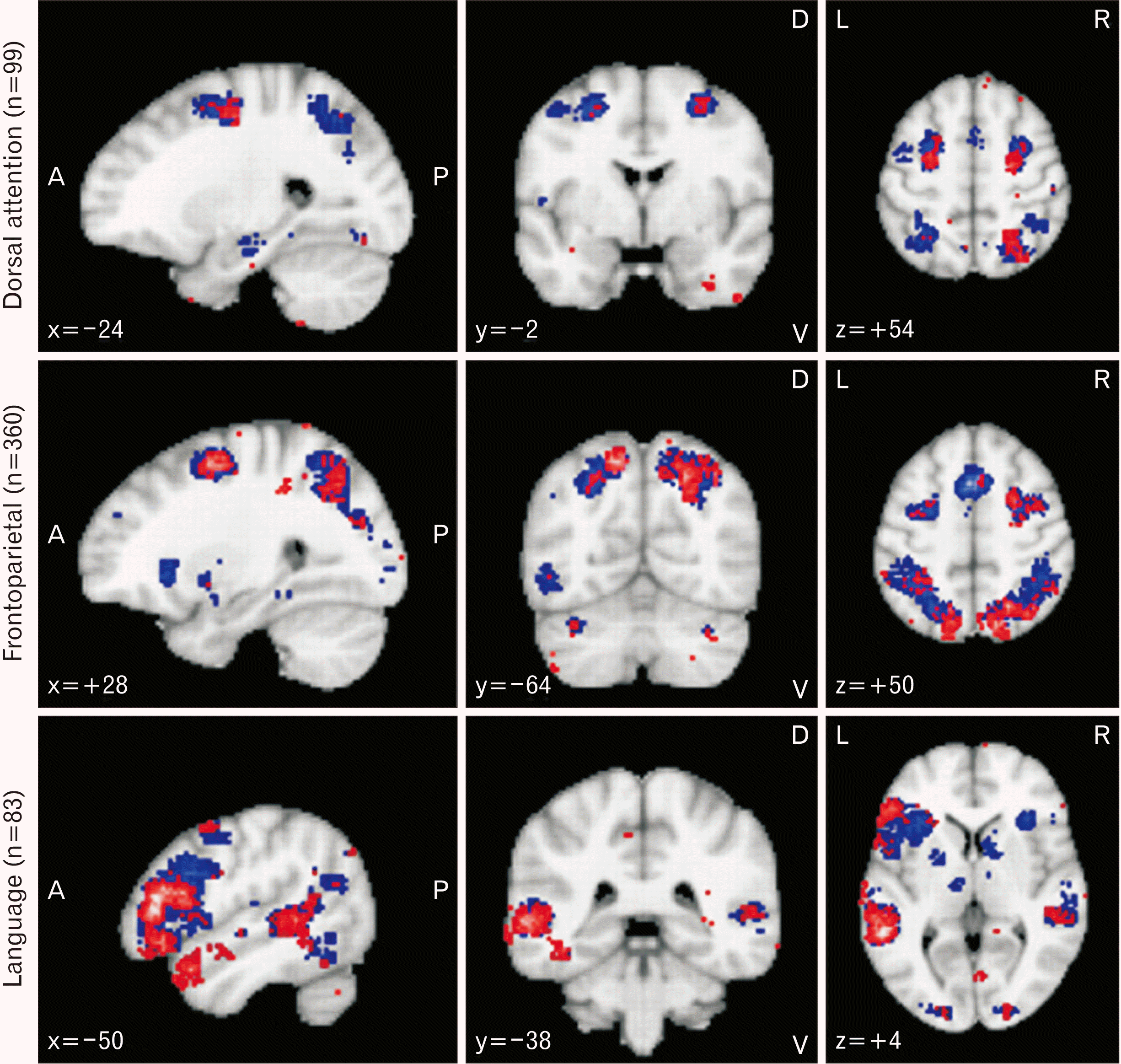
Fig. 1
The Structure of Consciousness. We are aware of external and internal worlds due to the activities of various large-scale functional brain networks. In addition, level of consciousness can affect the contents of consciousness. VN, visual network; AN, auditory network; SMN, somatosensory-motor network; ATN, attention network; FPN, frontoparietal network; CON, cingulo-opercular network; DMN, default mode network; LN, language network; EN, emotional network; MN, motivation network; SCN, social cognition network; SRN, self-referential network; Wake, wakefulness; MCS, minimally conscious state; VS, vegetative state; UWS, unresponsive wakefulness syndrome.
![]()
Fig. 2
Attention and language network. Results of meta-analyses on attention and language network obtained from neurosynth.org. The search terms used were “Dorsal Attention” (n=99 studies); “Frontoparietal Network” (n= 360 studies); and “Language Network” (n=83 studies). Association test and uniformity test map results are shown in red and blue; respectively. Only positive results are depicted. Results are false discovery rate (FDR) corrected at P<0.01. Further details are available at neurosynth.org.
![]()
Initially, we review major large-scale functional brain networks associated with the horizontal connectivity (cortico-cortical communication) that underlies awareness of consciousness (attention/language/default mode networks). However, primary sensory and motor networks, including visual, auditory, and somatosensory-motor networks, are not reviewed. This is followed by a review of the wakefulness network that is associated with vertical connectivity (subcortical-cortical communication). Finally, we introduce the interaction between horizontal and vertical connectivity.
Attention Network
The attention system can be classified into orienting and executive networks [11]. The orienting attention network is involved in the orientation of external (vision or other) stimuli. This network can prioritize inputs by selecting a modality or location. The brain areas involved are divided into dorsal and ventral attention networks (VAN). The dorsal attention network (DAN) is associated with top-down visuospatial orientation [12, 13] and enables the orientation to external stimuli and directs motor or eye movements [14]. The brain areas involved (Fig. 1) include the frontal eye field (FEF) and the superior parietal lobe (SPL)/intraparietal sulcus (IPS) [15]. The VAN is associated with switch and bottom-up reorientation, and the involved brain areas include the ventral frontal cortex (middle and inferior frontal gyrus [IFG]) and the temporoparietal junction (TPJ) [16]. Interestingly, synchronization between the orienting network and primary sensory brain areas, such as the visual area, enhance the sensory sensitivity [17].
The executive network handles target selection and focal attention [18-21]. In task experiments, this network showed greater activity for targets than nontargets, for conflict than for non-conflict trials, and for incorrect rather than correct trials. This network is also associated with top-down regulation, self-control, problem-solving, planning, and goal implementation. The executive network can be divided into the frontoparietal and cingulo-opercular networks. Task experiments have shown that the frontoparietal network controls task switching after a start-cue and the initiation of adjustments within trials. This network mainly involves lateral frontal and parietal regions (Fig. 1). On the other hand, the cingulo-opercular network controls the background maintenance of task performance across trials and performance-monitoring. The medial frontal/cingulate cortex and anterior insula (AI) are involved in the cingulo-opercular network, and interestingly Von Economo neurons, which are found in higher mammals, are located in this network [22].
Language Network
Visual stimuli such as words and sentences can evoke a language network (LAN), and the existence of this network is suggested by several neurological symptoms such as sensory, motor and conduction aphasia and by the results of fMRI studies [23-26]. Furthermore, left lateralization of the LAN has been reported [27, 28]. Its main components include many regions located in frontal, parietal, and temporal lobes (Fig. 1). Connection pathways between its components can be classified into two groups [29-31]. The ventral pathway is mainly involved in processing visual information into meaning (semantics), and the other, the dorsal pathway is mainly associated with visuo-phonology.
The inferior fronto-occipital fasciculus (IFOF), inferior longitudinal fasciculus (ILF) and uncinated fasciculus (UF) constitute the ventral sematic pathway, in which the IFOF connects the occipital cortex, superior parietal lobule, and fusiform gyrus with the IFG and dorsolateral prefrontal cortex (DLPFC). The ventral sematic pathway is critical for verbal and nonverbal semantic procession, and electrical stimulation of this pathway has been reported to cause semantic paraphasia [29, 32, 33]. The ILF links the occipital cortex with the anterior temporal lobe and provides critical pluri-modal integration of multiple semantic-related signals [34]. The UF, which plays a role in proper name retrieval [35], connects the temporal pole (TP) with the pars orbitalis of the IFG [36, 37].
The arcuate fasciculus (AF) constitutes the dorsal pathway. Its deep component connects posterior parts of the middle temporal gyrus (MTG) and inferior temporal gyrus (ITG) with the IFG, pars triangularis, and opercularis [38]. The AF is involved in phonological processing and its lesions can induce conduction aphasia [39, 40]. The superficial component (superior longitudinal fasciculus [SLF] III or the horizontal component of the AF) connects the supramarginal gyrus (SMG) and posterior part of the superior temporal gyrus (STG) with the ventral premotor cortex and contributes to the translation of phonological data into articulatory motor programs [38, 41].
Default Mode Network
The existence of the DMN was suggested by Shulman et al. (1997) [42]. This network subserves internal mentation including self-reference, emotion, and motivation (Fig. 3). The DMN can be deactivated by focusing on the external world [43-45]. Although the topic remains under debate, some core cortical areas of the DMN usually include the ventral medial prefrontal cortex (VMPFC), the dorsal medial prefrontal cortex (DMPFC), the posterior cingulate/retrosplenial cortex (PCC/Rsp), the inferior parietal lobule (IPL), the lateral temporal cortex (LTC), and the hippocampal formation (HF) (Fig. 2). Also, most researchers agree there are from one to nine subnetworks [46, 47]. In a recent study that used the within-individual approach instead of the group-average approach, it was suggested that the DMN consists of nine subnetworks, which include the parietal, ventromedial, pregenual, retrosplenial, posterior medial temporal lobe, and four lateral networks [47]. Interestingly, these four lateral networks work as connecting hubs that link to other large-scale networks such as the language and frontoparietal networks. The parietal network functions as a central coordinator among these nine subnetworks and is related to internal mentation and social cognition [45, 48] and includes brain areas such as the medial parietal cortex, middle angular gyrus, anterior VMPFC, anterior medial superior frontal gyrus, middle lateral superior frontal gyrus, anterior superior temporal sulcus, ventral posterior cerebellar lobe, and vermis. The ventromedial subnetwork has been associated with fear and anxiety [49, 50] and contains brain areas such as the VMPFC, amygdala, and anterior hippocampus. The pregenual subnetwork has been associated with reward, expected value, decision-related positive emotion, and salience [50-55] and contains brain areas such as the pregenual MPFC, anterior striatum, and ventral AI. The retrosplenial subnetwork has been reported to be involved in the processing of contextual and scene information [56] and contains the Rsp, parieto-occipital fissure, and parahippocampal cortex. The posterior medial temporal lobe subnetwork is critical for long-term memory [57-61] and includes the posterior medial temporal cortex and caudal hippocampus.
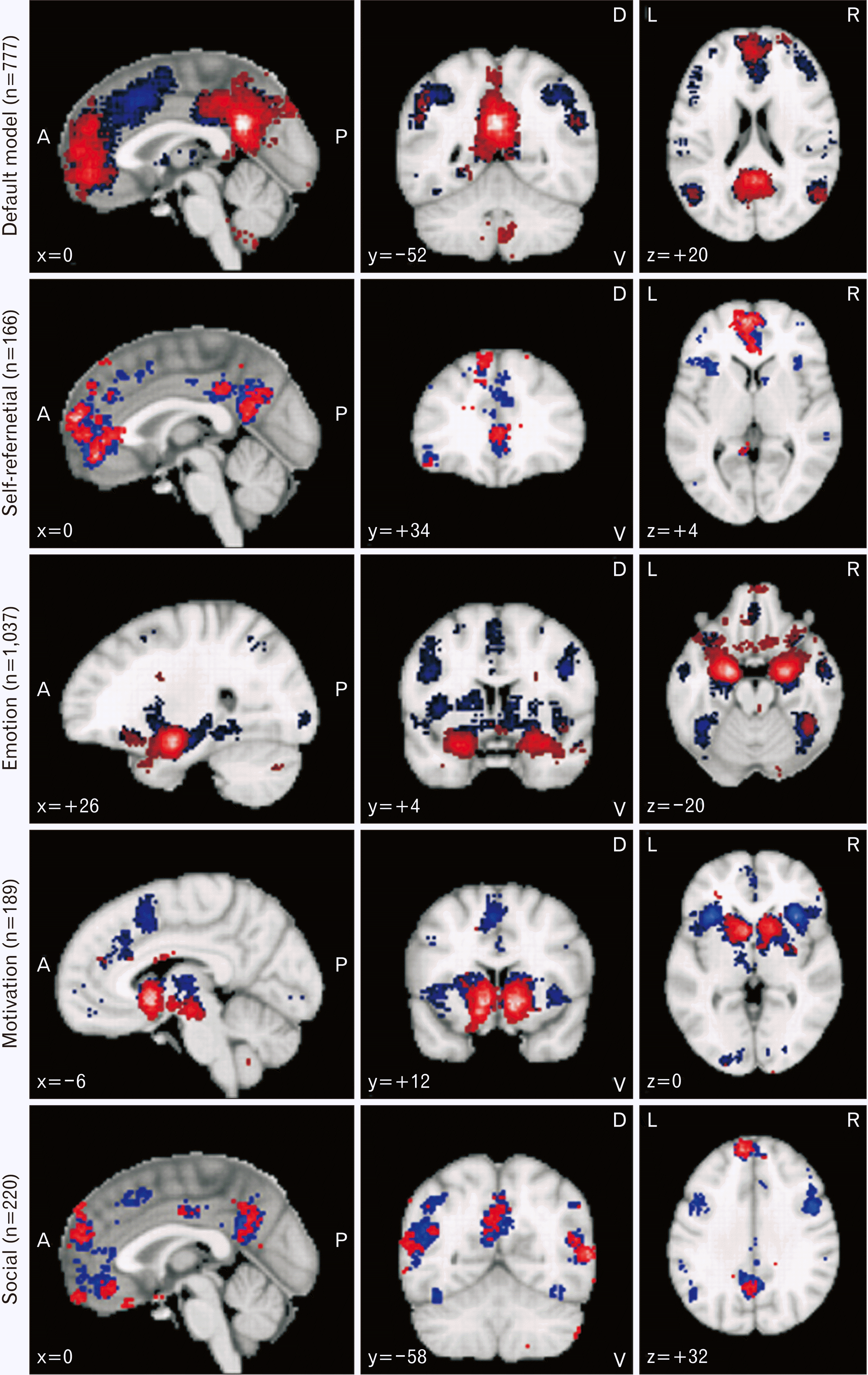
Fig. 3
Default mode network. Results of meta-analysis for the default mode network obtained from neurosynth.org. The search terms used were “Default Mode” (n=777 studies); “Self-referential” (n=166 studies); “Emotion” (n=1,037 studies); “Motivation” (n=189 studies) and “Social Cognition” (n=220 studies). Association test and uniformity test map results are shown in red and blue; respectively. Only positive results are shown. Results are false discovery rate (FDR) corrected at P<0.01. Further details are available at neurosynth.org.
![]()
Self-referential network
The self-referential network is one of the most critical networks in DMN because thoughts about self predominate during the resting state. In contrast with other referential processes, self-referential processes can be defined from various perspectives and handled by several kinds of self such as executive self, observing self, neutral self, emotional self, and others [62]. Executive self is associated with the frontoparietal and DAN, whereas emotional self is related to the affective network. Observing self means observing self as an object. Unlike emotional self, neutral self is unaffected by emotion and occurs spontaneously in the resting state compared, whereas observing self usually requires active attention to observe self.
The MPFC is the most critical brain area of the self-referential network and can be divided into four areas (V-, M-, D1-, D2-PFC) [62-64]. V- (Area 11) and D2- (Area 8) are associated with emotional and executive processes, respectively [65, 66], and M- (Area 10) and D1-(Area 9) are associated with neutral and observing selfs, respectively [67, 68]. For self-referential processes, other areas such as the anterior cingulate cortex (ACC), PCC, precuneus (PrC), insula and IPL/TPJ are also involved (Fig. 2). Based on analyses of the effective network, PCC, and PrC areas send signals continuously to the MPFC, which then presents selected signals to consciousness [69-71]. In this model, posterior midline structures are critical for the integration of information from interoception and exteroception.
Emotion and motivation network
Emotions can be defined as organized affective reactions to significant internal and external events [72, 73]. Extensive studies have been conducted to explain the neurobiological basis of emotions, which range from the Papez circuit to the theory of constructed emotion. Because the emotional network processes various emotions (e.g., fear, anxiety, anger, sadness, and happiness), it may consist of heterogeneous subnetworks. Recently, three new theories encapsulating concepts of emotion were suggested; affective neuroscience theory [73-76], reward sensitivity theory [77], and the theory of constructed emotions [78]. Affective neuroscience theory suggests that basic emotions are evolutionarily conserved in non-human animals. In several studies, ultrasonic vocalizations (USVs) were used to detect the affective statuses of non-human animals [79-81]. Furthermore, efforts have been made to localize seven types of emotion (seeking, lust, care, play, fear, anger/rage, panic/sadness) in the brain map [82]. Although basic emotions are evolutionarily conserved across species, results from non-human animals have yet to be confirmed in man. Reward sensitivity theory describes three systems in brain that link stimuli and responses, i.e., the behavioral activation system (BAS), the fight-flight-freezing system (FFFS), and the behavioral inhibition system (BIS). The BAS and FFFS involve appetitive stimuli and threats, respectively, whereas the BIS promotes exploratory behaviors in unfamiliar situations. The theory of constructed emotion suggests that emotions are constructed by the brain to aid understanding of incoming environmental information.
Brain areas involved in the emotional network usually include the VMPFC, amygdala, ACC, PCC, insula, thalamus, and periaqueductal gray matter [72] (Fig. 2), and activations of these areas depending on emotion type. For example, a proximal threat can activate periaqueductal gray matter and central nuclei of the amygdala [83], but a distant threat activates the VMPFC. A recent study that used multivariate pattern classification showed that distinct subnetworks representing different emotions are observed in the DMN [84].
Motivation can be defined as a process that determines direction and energization [72]. Although motivation is more related to behavior-production and emotion to experiential processes, the motivation network is associated with the emotional network in several respects. Many emotional states promote behaviors that are essential components of motivation [85], for example, anger can provoke offensive behaviors, and in the conditioned place preference test, animals prefer a location associated with reward [86]. Moreover, neuroanatomical areas associated with emotion or motivation tend to overlap. Kim et al. proposed a model to explain motivational behavior [87, 88]. According to this model, the motivation process consists of three phases: generation (anticipation of reward), maintenance (value-based decision making), and modulation (cognitive regulatory control). The motivation network usually includes the ventral tegmental area, nucleus accumbens (NAcc), ventral striatum (VS), amygdala, and MPFC (Fig. 2), and the NAcc and mesolimbic pathways are critical for the generation of motivation [89, 90]. The activity of the NAcc is correlated with degree of excitement, and connectivity between the MPFC and VS is critical for grit [72]. Connectivity between the dorsal anterior cingulate cortex (dACC) and dorsal and VS is associated with growth mindset [91], and the orbitofrontal cortex (OFC) is important for value evaluation [87].
Social cognition network
Like other social animals, humans can ‘sense’ the mental states of others [92], that is, we can reasonably predict the emotional statuses (empathy), intentions, and perspectives (Theory of Mind) of others, and based on our perceptions, we perform prosocial activities that reward these others. Furthermore, these prosocial activities are critical for eliciting healthy social interactions. Empathy can be defined as the ability to understand and share the affective states of others, and the anterior cingulate gyrus (ACCg) and subgenual anterior cingulate gyrus (sgACC) are usually involved in the process of empathy [92, 93]. The Theory of Mind concerns ability to assess the mental states of others [94-96], and several kinds of tests such as the Sally-Ann test, the dot perspective task, and the ambiguous figure perception test support this theory [97, 98]. Perspective-taking mainly involves the TPJ, DMPFC, and PrC (Theory of Mind) [99-101]. Prosocial activity refers to actions undertaken to obtain a reward for another person despite the personal effort/cost involved, and this activity largely involves the sgACC and TPJ [92, 102-104].
Goal-directed behaviors derived from social or non-social motivation are based on subjective valuation, which computes physical and cognitive efforts and rewards. Neuronal activities in the dACC, AI, VS, basolateral amygdala (BLA), VMPFC, and DLPFC have been shown by fMRI studies to be correlated with efforts and rewards [105-107]. Moreover, single-cell recordings have shown that a subset of neurons in the dACC signal the amount of reward on offer, and another neuron subset in the same area signals the amount of effort required [108, 109].
Wakefulness Network
Wakefulness refers to levels of arousal and behavioral states during which an animal exhibits voluntary motor activation and is responsive to internal and external stimuli [110]. Wakefulness states can be described as: comatose, vegetative (unresponsive wakefulness syndrome), or minimally conscious [3]. During the comatose state, patients do not respond to extensive stimuli. In the vegetative state patients exhibit spontaneous breathing, spontaneous or stimulus-induced eye opening, blinking to a visual threat, auditory startle response, orient eyes or head to stimuli, stereotyped posturing, and show normal flexion withdrawal to noxious stimuli and gag/deglutination/oral reflex. The minimally conscious state (MCS) can be further divided into two states: MCS+ and MCS–. Patients in the MCS– state show simple non-reflex movements (visual pursuit, orientation to pain or non-contingent behavior [smiling]), and those in the MCS+ state respond to simple commands. These behaviors suggest MCS patients have some degree of cortico-cortical communication but that communication is poor in UWS patients. Level of wakefulness is also naturally regulated by circadian rhythm (sleep cycle) [110]. Polysomnography (EEG, electrooculography, and electromyography) results show natural sleep can be divided into non-rapid eye movement (NREM) sleep (N1, relaxed wakefulness; N2, light sleep; N3, deep sleep), and REM sleep.
Level of wakefulness is regulated by vertical connectivity from the brain stem (reticular formation system, locus ceruleus, ventral tegmental nucleus, raphe nucleus, and others) through hypothalamus and thalamus to cortex [110, 111], and damage to this vertical connectiveness can alter the status of wakefulness [112, 113]. During slow-wave sleep, low frequency synchronized activity between the cortex and thalamus blocks sensory input to the cortex [114]. However, when awake, thalamocortical activity is attenuated. Moreover, contents of awareness can alter depending on the level of wakefulness.
The suprachiasmatic nucleus is the central pacemaker of circadian rhythm and regulates wake-promoting, sleep-promoting, and REM-producing circuits in a time-dependent manner [115, 116]. Orexin-positive neurons in the lateral hypothalamus [117-119], cholinergic neurons in pedunculopontine and laterodorsal tegmental nuclei [120, 121], monoaminergic neurons in locus coeruleus [122, 123], dorsal and median raphe nuclei [124, 125], and ventral tegmental nuclei [126, 127] and histaminergic neurons [128, 129] constitute wake-promoting circuits. These circuits might be divided into two pathways, i.e., the ventral and dorsal pathways [110]. The former links brain stem areas with hypothalamus, basal brain, and cortex, and regulates the behavioral state of wakefulness, whereas the dorsal pathway links brain stem areas with thalamus and regulates contents of consciousness. GABAergic neurons in ventrolateral and median preoptic nuclei regulate sleep-promoting circuits [130, 131], and glutamatergic neurons in the sublaterodorsal nucleus [132, 133] and cholinergic neurons in the pedunculopontine and laterodorsal tegmental nuclei are important for REM-promoting circuits [134, 135].
Interaction between Horizontal and Vertical Connectivity
The activities of large-scale brain networks have been reported to depend on level of wakefulness. A recent study analyzed the dependence of spatiotemporal dynamics of large-scale brain networks on sleep stages [136] and showed that up to 10 brain networks can co-occur. The average number of co-occurring networks found was 2–5, and the mean duration of each network was 7.3 seconds. Activities of all brain networks are maintained through sleep stages and are maximally activated during the N2 stage. Notably functional interrelations between networks decrease in the deep sleep stage. During this stage, connectivity between the MPFC and PCC in the DMN and activity of the attention network decrease, whereas the activity of the posterior DMN increases. In addition, the duration of brain network activity decreases in the deep sleep stage, which suggests cortico-cortical connectivity breakdown.
Spatiotemporal dynamics of brain networks have also been examined in drug-induced (ketamine and propofol) states of altered wakefulness and in the presence of neuropathologies (UWS) [137]. This study showed reduced brain network entropy and uniformity of the occurrence rates of different brain networks in unresponsive states. Interestingly, it was also shown that preferred transition trajectories exist between brain networks. In the conscious state, accessibility to different brain networks is well maintained, but in unresponsive states, visual (VIS) and VAN networks are monopolized, and the negative correlation between DMN and DAN is attenuated. Accordingly, sleep stage studies show the persistence probabilities of brain networks are reduced in unresponsive states.
Future Direction
In the present study, we reviewed major large-scale functional brain networks related to consciousness. Brain mapping of networks has been largely performed using neuro-imaging data. However, because the brain is composed of neurons and glial cells, further studies are needed to explore the cellular-scale mechanisms subserving consciousness to identify its neural correlates. Cellular-scale mechanisms should explain how distant brain regions are connected or coupled transiently to achieve functional connectivity. Because one specific stimulus should not activate all anatomically connected brain regions for specific psychological or cognitive processes, unnecessary coupling needs to be inhibited to establish specific neural networks. Recently micro- and macro-circuits have been suggested for long-distance connections between brain areas. Glutamate release from distant pyramidal neurons can activate target pyramidal neurons and vasoactive intestinal peptide (VIP)-positive GABAergic interneurons [138-141], and thus, modulate the feedback loop between GABAergic interneurons and pyramidal interneurons. This disynaptic disinhibition generates gamma oscillations in distant pyramidal neurons and stabilizes local information. Moreover, the selective activations of subnetworks and inactivations of other subnetworks can be achieved by inhibiting α oscillation-induced gating [142-145]. More detailed mechanistic studies are needed to characterize the micro- and macro-circuits underlying consciousness.
 XML Download
XML Download