

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Cellular aging or senescence, a process which irreversible cell cycle arrest was drove, is a sort of cellular response to various types of stresses including oxidative stress, DNA damage, metabolic alterations and oncogene activation [1]. Oxidative stress is thought to be the most redundant source to trigger cellular aging or senescence in the point of the molecular basis on aging [2,3]. Chronic oxidative stress stems from an imbalance of redox status [4,5]. One of the characteristics of aging is the decline in the level of anti-oxidants during senescence [6], which indicates the physiological importance of oxidative stress in cellular senescence and aging.
DNA damage is associated with the modulation of Saccaromyces cerevisiae chromatin silencing factor Sir2 (silent information regulator-2), which is one of the main factors that determines life span. Mammalian sirtuin 1 (SIRT1), nicotinamide adenosine dinucleotide (NAD)-dependent protein deacetylase, highly conserved with Sir2, participates in regulation of many cellular events including cell cycle, senescence, apoptosis and metabolism by interacting with key molecules for those events like p53, signal transducer and activator of transcription (STAT3) and Peroxisome proliferator-activated receptor gamma coactivator (PGC1α) [7-9]. A previous study has shown that SIRT1 neutralizes progressive multifocal leukoencephalopathy (PML)-induced premature senescence by deacetylation of p53 in mouse embryo fibroblasts [7]. Recently many evidences have been reported that inhibition or downregulation of SIRT1 induces premature senescence-like phenotype through the augmentation of p53 acetylation in human cancer and endothelial cells [10]. Reversely, overexpression of SIRT1 makes cells more protective from senescence-related physiological alterations in cells [8]. Therefore, it would be potential target to be able to regulate redox and SIRT1 expression for deferring aging process.
Luteolin is a highly abundant natural flavonoid that exists in celery, green pepper and chamomile. Luteolin has been reported to have anti-inflammatory, anti-oxidant and anti-carcinogenic activities [11-13]. It has been reported as a potent inhibitor to be able to restrain tumor necrosis factor-α (TNF-α) and Interleukin 6 (IL-6), and nitric oxide expression induced by lipopolysaccharide [11]. It also has been considered as an anticancer agent through its inhibitory role for metastasis and angiogenesis, enhancing role of cell cycle arrest and apoptosis [14]. A latest study has indicated that luteolin ameliorates cisplatin-induced acute kidney injury in mice by regulating p53-dependent renal tubular cell apoptosis [15]. It has also been shown that luteolin inhibits microglia and alters hippocampal-dependent spatial working memory in aged mice and exhibits protective roles in neurodegenerative diseases [16,17]. However, there is a lack of direct evidence showing the association between luteolin and the observed anti-aging effect in vitro as well as the relevant molecular mechanisms. In this study, we investigated the inhibitory effect of luteolin against hydrogen peroxide (H2O2)-induced cellular senescence and the molecular mechanism of its inhibitory action with focus on modulation of SIRT1 and p53.
METHODS
Materials
Luteolin, 5-bromo-4-chloro-3-indolyl-beta-D-galactoside (X-Gal) and H2O2 were purchased from Sigma-Aldrich (St. Louis, MO, USA). Antibodies against the following proteins were used: phospho-p53 (p-p53, Cell Signaling Technology, 9284), p21 (Santa Cruz Biotechnology, sc-397), Sirt1 (Merck Millipore, 07-131), β-actin (Santa Cruz Biotechnology, sc-1616), phospho-JNK (p-JNK, Cell Signaling Technology, 9251), phospho-ERK (p-ERK, Cell Signaling Technology, 9101), and phospho-p38 (p-p38, Cell Signaling Technology, 9211). The specific siRNA for p53 (sc-29436) and SIRT1 (sc-40987) was purchased from Santa Cruz Biotechonology (Santa Cruz, CA, USA). Unless indicated otherwise, all other chemicals were obtained from Sigma-Aldrich.
Cell culture
House Ear Institute-Organ of Corti 1 (HEI-OC1) cells, derived from long-term cultures of Immortomouse cochlea, are extremely sensitive to ototoxic drugs. HEI-OC1 cells possess hair cell-like properties, express several specific markers, including Math 1 and Myosin 7a [18]. HEI-OC1 cells obtained from Center for Metabolic Function Regulation, Department of Microbiology, Wonkwang University (Iksan, Korea). Cells were cultured with high-glucose DMEM containing 10% FBS and stored in a humidified incubator (33 degree, 5% CO2) without antibiotics.
Cell viability and cell proliferation by MTT assay
Cell viability and cell proliferation was performed as Yu et al. [19] and Lee et al. [20] described, respectively. Briefly, Cells (4 × 104/well) were seeded in 96-well plates and treated with luteolin and H2O2 as described in figure legends (Fig. 1A, D). Then 20 μl of MTT (3-[4,5-dimethythiazol-2-yl]-2,5-diphenyltetrazolium bromide, a tetrazolium salt) test solution was added and incubated for 4 h at 37°C in a humidified incubator with 5% CO2. The absorbance was measured at a wavelength of 490 nm using a SpectraMax M3 instrument (Molecular Devices, Sunnyvale, CA, USA).
Fig. 1
Luteolin reduces hydrogen peroxide (H2O2)-induced cellular senescence in House Ear Institute-Organ of Corti 1 (HEI-OC1) cells.
(A–C) Cells were pretreated with luteolin for 12 h, then incubated in 30 μM H2O2 for 3 days. Cell viability was determined by MTT assay (A). The percentage of SA-β-gal positive cells out of total cells was counted and the average data was obtained from three independent experiments (B). The senescent phenotype of HEI-OC1 cells was detected by the SA-β-gal assay (×100) (C). (D) Cell growth was evaluated by MTT assay at various time points indicated in the figure after addition of H2O2. Data represents means values of triple experiments. *p < 0.05 vs. control, #p < 0.05 vs. H2O2. Piperine was used as a positive control.
![]()
Senescence-associated β–galactosidase (SA-β-gal) staining
The proportion of SA-β-gal positive cells was determined as described by Dimri et al. [21]. HEI-OC1 cells were washed twice with phosphate-buffered saline (PBS) and fixed with 3.7% formaldehyde in PBS for 10 min at room temperature. After washing with PBS, cells were incubated with β-galactosidase reagent (1 mg/ml X-gal, 40 mM citric acid/sodium phosphate buffer, pH 6.0, 5 mM potassium ferrocyanide/potassium ferricyanide, 150 mM NaCl, 2 mM MgCl2) at 37°C. The cells were washed twice with PBS, then we counted old cells (blue) and total cells under fluorescence microscopy using a Zeiss microscope.
Western blot analysis
Western blot analysis was performed as Cheng et al. [22]. HEI-OC1 cells were harvested, washed with ice cold 1×PBS, and resuspended in pre-cold 50 mM Tris-HCL buffer (pH 7.4) containing protease inhibitors (2 mM phenylmethanesulfonyl fluoride, 5 mM protease inhibitor cocktail, 1 mM Na orthovanadate). Protein concentration was measured at 595 nm using a SpectraMax M3 instrument. Sample (50 μg) were separated by 12% sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and were transferred to Polyvinylidene fluoride (PVDF) membranes. The membranes incubated in first antibody overnight and then incubated in secondary antibody at 4°C for 2 h. Proteins bands were detected using enhanced chemiluminescence system (Amersham Pharmacia Biotech, Piscataway, NJ, USA).
SIRT1 activity assay
HEI-OC1 cells were harvested, washed with ice-cold 1×PBS, and lysed with NETN buffer (1 mM EDTA, 20 mM Tris-HCl, pH 8.0, 100 mM NaCl, 0.5% NP-40). A SIRT1 fluorometric drug discovery kit (BML-AK511; Enzo Life Sciences, Plymouth Meeting, PA, USA) was used to detect SIRT1 activity. Briefly, Lysates incubated with SIRT1 assay buffer (45 min, 37°C), and then stopped by SIRT1 Developer (15 min, room temperature). At last, the sample was analysed by a Spectra M3 instrument at 355 nm/460 nm.
Comet assay
Cellular DNA damage was performed using a OxiSelect Comet Assay Kit according to the manufacturer’s protocol (Cell Biolabs, Inc, San Diego, CA, USA) as previously described [23]. Images were obtained and analyzed by 15 V. Komet 5.5 (Comet Assay Software; ANDOR technology, Belfast, Northern Ireland).
RESULTS
Luteolin inhibits H2O2-induced cellular senescence in HEI-OC1 cells
In this study, we evaluated the effect of luteolin on the viability of HEI-OC1 cells to determine its cytotoxic potential. Up to a concentration of 2 μM luteolin, no cytotoxic effects could be detected using the MTT assay (Fig. 1A). Piperine is a major plant alkaloid presented in pepper and was used as a positive control in present study. To investigate the effect of luteolin on the HEI-OC1 cells with senescent phenotype, cells were treated with 30 μM of H2O2 (Supplementary Fig. 1) for induction of cellular senescence. We found that pretreatment with luteolin inhibited the senescent phenotype as shown by the results of the SA-β-gal assay and appearance of enlarged cell morphologies at 3 days. After treatment with H2O2, 65% of the cells were found to be SA-β-gal-positive, which was reduced to only 17% in the presence of luteolin (Fig. 1B). The SA-β-gal-positive cells also exhibited senescent morphological changes such as enlarged and flattened shapes (Fig. 1C). Next, we monitored cell growth for 5 days under same condition. As expected, H2O2 significantly reduced the cell growth rate but the cells treated with luteolin are rescued more than 50% (Fig. 1D). In this setting, we did not find major differences on the portion of propidium iodide-stained cells, indicating that the reduced proliferation was not interfered by an increase in apoptosis (data not shown). These results indicated that luteolin effectively protected the HEI-OC1 cells against H2O2-induced cellular senescence.
Protective effect of luteolin against H2O2-induced cellular senescence is dependent on p53
As p53 and p21 are known to be major players in the induction of senescence [24], we investigated whether p53 played a role in mediating the protective effect of luteolin against H2O2-induced cellular senescence in HEI-OC1 cells. As shown in Fig. 2A, we observed that after H2O2 treatment, there was increased phosphorylation of p53 and expression of p21, both of which were decreased after luteolin treatment. Meanwhile, 80 nM p53 siRNA efficiently inhibited H2O2-induced p-p53 expression (Fig. 2B). We also found that blocking p53 using siRNA reversed luteolin effects on H2O2-treated cells as shown by the SA-β-gal activity assay results and senescent morphological changes on 3 days (Fig. 2C, D). These data suggested that luteolin protect cells from H2O2-induced cellular senescence via modulation of p53 phosphorylation and its downstream molecule, p21 expression.

Fig. 2
Protective effect of luteolin against hydrogen peroxide (H2O2)-induced cellular senescence is dependent on p53.
(A) Cells were pretreated with 2 μM luteolin for 12 h, then incubated in 30 μM H2O2 for 3 days. (B) Cells were transfected with various concentrations siRNAs of control or p53, then incubated with 30 μM H2O2 for 3 days. The protein levels were detected by Western blotting and quantified by densitometry based on immunoblot images (A, B). (C) Representative images of SA-β-gal staining of HEI-OC1 (×100). (D) The percentage of senescent cells were calculated in (D). Data represents means values of triple experiments. *p < 0.05 vs. control; #p < 0.05 vs. H2O2-treated cells.
![]()
Luteolin increases SIRT1 expression in HEI-OC1 cells
As shown in Fig. 1, luteolin protects against H2O2-induced cellular senescence and hence, we considered that luteolin might promote the expression of the longevity gene, SIRT1. HEI-OC1 cells were treated with various concentrations of luteolin (0–4 μM) for 12 h. The effect of luteolin on SIRT1 expression is shown in Fig. 3A and B. As shown in Fig. 3A, luteolin induced the expression of SIRT1 in a dose-dependent manner. The maximal induction of SIRT1 was achieved at a dose of 2 μM luteolin (Fig. 3A). The induction of SIRT1 expression by luteolin (2 μM) was detected as early as 3 h following luteolin treatment; the augmentation lasted for an additional 9 h (Fig. 3B). SIRT1 activity was found to increase over the same luteolin dose range (Fig. 3C) and to reach a maximum level after 12 h of luteolin treatment (Fig. 3D).
Fig. 3
Effects of luteolin on sirtuin 1 (SIRT1) expression in House Ear Institute-Organ of Corti 1 cells (HEI-OC1).
(A, C) HEI-OC1 cells were treated with luteolin (0–4 μM, 12 h). (B, D) Cells were treated with luteolin (2 μM, 0–12 h). The proteins level (A, B) and SIRT1 activity (C, D) were performed with Western blotting and SIRT1 fluorometric kit, respectively. The proteins were quantified by densitometry based on immunoblot images. Data represents means values of triple experiments. Significance vs. control: *p < 0.05.
![]()
Inhibition of SIRT1 abrogates the protective effect of luteolin against H2O2-induced cellular senescence
To determine the role of endogenous SIRT1 in cellular senescence, HEI-OC1 cells were treated with SIRT1 siRNA. As shown in Fig. 4A and B, p-p53 and p21 expression were down-regulated by treatment of luteolin, while p-p53 and p21 expression were restored by silencing of SIRT1. In addition, SIRT1 inhibition abrogated the effect of luteolin on senescence-specific morphological changes and SA-β-gal activity. Under SIRT1 siRNA transfection, luteolin treatment did not increase the activity and expression of SIRT1 (Fig. 4C, D). These results demonstrated that luteolin inhibited oxidative stress-induced cellular senescence, which might be mediated by modulation of p53 and SIRT1.
Fig. 4
Luteolin protects against hydrogen peroxide (H2O2)-induced p53 phosphorylation via sirtuin 1 (SIRT1) protein expression.
House Ear Institute-Organ of Corti 1 (HEI-OC1) cells were transfected with 80 nM siRNAs of control or SIRT1, then incubated in 2 μM luteolin for 12 h, followed by 30 μM H2O2 treatment for additional 3 days. (A, B) The protein levels were detected by Western blotting and quantified by densitometry based on immunoblot images. (C) Representative images of SA-β-gal staining of HEI-OC1 (×100). (D) The percentage of senescent cells were calculated in (D). Data represents means values of triple experiments. *p < 0.05 vs. control; #p < 0.05 vs. H2O2-treated cells.
![]()
Luteolin prevents H2O2-induced DNA damage in HEI-OC1 cells
There is a significantly increase in oxidative DNA damage in senescent cells, and the levels are similar to that seen in aging and age-related diseases [25,26]. Therefore, we examined whether luteolin has any protective role against the DNA damage caused by H2O2. We performed single cell gel electrophoresis assay, comet assay, to detect the DNA damage at individual cell level. As shown in Fig. 5A and B, the cells were treated with 2 μM luteolin for 12 h after exposure to 30 μM of H2O2 for 3 days show much shorter comet tails and smaller comet tail areas than the H2O2-only exposed cells (Fig. 5A). This significant difference in the tails between the groups is shown quantitatively in Fig. 5B, clearly indicating that luteolin markedly protects the cells against DNA damage under H2O2. To verify our hypothesis, we repeated this experiment in the SIRT1 or p53 silenced cells. As shown in Fig. 5A and B, SIRT1 siRNA efficiently abolishes the protective effect of luteolin on H2O2-induced DNA damage, whereas p53 siRNA did not show any significant changes. These results further underline the contribution of the accumulating DNA damage to cellular senescence and suggest that the SIRT1 plays a role in the prevention of DNA damage while p53 does not.
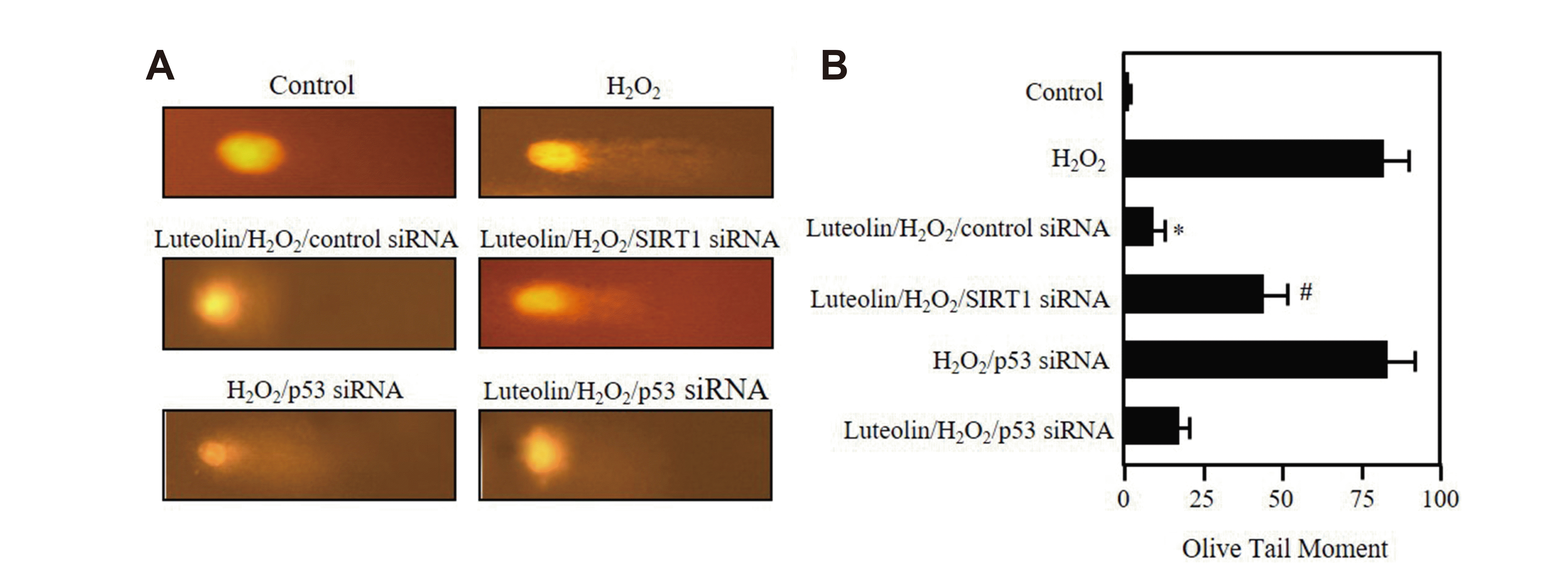
Fig. 5
Luteolin prevents DNA from hydrogen peroxide (H2O2)-induced damage.
(A, B) House Ear Institute-Organ of Corti 1 (HEI-OC1) cells were transfected with 80 nM siRNAs of sirtuin 1 (SIRT1) or p53, then incubated in 2 μM luteolin for 12 h, followed by 30 μM H2O2 treatment for additional 3 days. Comet assay was performed as described in Methods. Data show the results of one of three independent experiments. *p < 0.05 vs. H2O2, #p < 0.05 vs. luteolin/H2O2/control siRNA.
![]()
DISCUSSION
It has revealed that luteolin has protective roles in several disease models [11-14]. However, as far as we know, the role of luteolin in aging is still unknown. Here, we showed that luteolin inhibits H2O2-induced cellular senescence and enhances SIRT1 expression concomitantly, which played a crucial role in inhibiting p53 as well as suppressing the senescent phenotypes in auditory cells (Fig. 6).
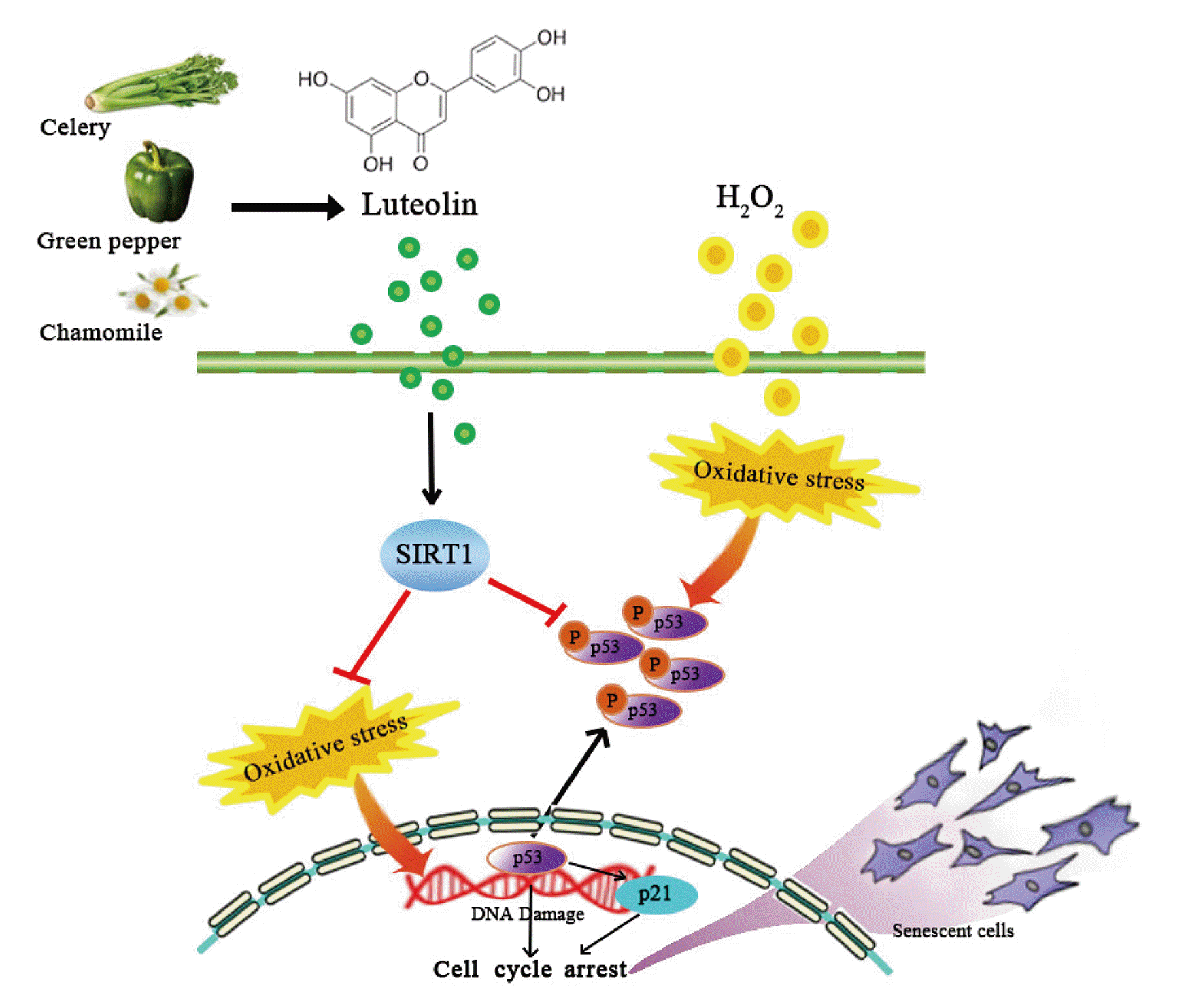
The anti-oxidant activity of luteolin has been associated with its ability to scavenge reactive oxygen species by inhibiting pro-oxidant enzymes and inducing anti-oxidant enzymes [27]. It has also been found to protect the microglia and alter hippocampal-dependent spatial working memory in aged mice [16,17]. As anticipated, luteolin prevented H2O2-induced cellular senescence as shown by attenuation of SA-β-gal signal and restring of cell growth (Fig. 1). In our observation, luteolin protects H2O2-induced cellular senescence by decreasing p53 phosphorylation (Fig. 2). Considering p53 is a key molecule that mediates multiple pathways involved in tumor suppression and senescence [28], several evidences that protective effect of Salidroside from premature senescence by regulation of redox and p53/p21 expression [29] and elevated level of p53 in H2O2 induced-prematurely senescent human diploid fibroblasts [30] raised us to investigate whether luteolin alters DNA damage under oxidative stress condition. Consistent with our expectation, luteolin ameliorated the H2O2-induced DNA damage (Fig. 5). As previous observation [29], luteolin downregulated p53 expression (Fig. 2).
SIRT1 plays an important role in protecting the cells against cellular oxidative stress and DNA damage [31]. Our results also showed that luteolin induces SIRT1 expression in HEI-OC1 cells and the upregulated SIRT1 expression prevented cellular senescence (Fig. 3). Previous studies have shown that SIRT1 overexpression has a protective role against oxidative stress-induced premature senescence in human endothelial cells [9] and reversely, SIRT1-deficient mouse embryonic fibroblasts extends replicative lifespan under sub-lethal oxidative stress [32]. Ota et al. [10] observed that senescence could be induced in WI38 and IMR90 cells using SIRT1 inhibitor. These effects could be reversed by inhibiting the transfection with SIRT1 siRNA. As shown in Fig. 4, SIRT1 siRNA efficiently abolishes the protective effect of luteolin on H2O2-induced DNA damage. Based on our data, luteolin influences SIRT expression (Fig. 5), which is indirectly implies its effect on DNA damage. Therefore, our observations along with ROS induced senescent pathway mediated by p53 are consistent with the concept that SIRT1 regulates important processes in cellular senescence. In addition, nicotinamide, a SIRT1 inhibitor, has been remarked as an important modulator to extend the replicative lifespan of primary human fibroblasts [33]. Taken together, SIRT1 has been considered as a key player to be able to make the resistance and modulate oxidative stress-induced premature senescence although still it is elusive whether it directly regulates lifespan. Interestingly, several reports have highlighted that dietary supplementation of flavonoids has a protective role against neurodegenerative, cardiovascular, inflammatory, metabolic diseases, and cancer by enhancing SIRT1 activity [34] and moreover, it has been reported that flavonoids, such as resveratrol, quercetin, and catechins, activate SIRT1 in vitro and in vivo [34-37]. Therefore, it would be promising to regulate SIRT1 activity by dietary flavonoids as a therapeutic application against oxidative stress-induced cellular senescence.
A previous study proposed that SIRT1 might inhibit the ability of p53 to promote endothelial senescence [9]. In our study, we observed that the increase in the phosphorylation of p53 was higher in senescent cells compared to non-senescent cells, which was significantly decreased after treatment with luteolin (Fig. 2). This decrease was abolished by blocking SIRT1 expression using SIRT1 siRNA (Fig. 4), suggesting that the downregulation of p53 phosphorylation by luteolin is at least partially mediated by SIRT1. These findings are quite consistent with previous reports demonstrating that inhibition of SIRT1 by siRNA increased the acetylation of p53 in endothelial cells [9]. The SIRT1/p53 pathway in cellular proliferation and senescence is tightly linked with transcriptional alterations of downstream cell cycle inhibitors [32]. In the same manner, our data showed that regulation of p53 phosphorylation determines p21 expression level in HEI-OC1 cells (Figs. 2A, 4A).
Taken together, this study provides relevant evidences supporting that luteolin exerts protective effects against oxidative stress-induced cellular senescence which is derived by activation of p53. Subsequent enhancement of SIRT1 expression by luteolin plays a crucial role in inhibiting p53 activation as well as suppressing the senescent phenotype in auditory cells. These findings present luteolin as a potential and bio-safe natural compound to be able to impede aging process and attenuate age-related diseases incidence.
SUPPLEMENTARY MATERIALS
Supplementary data including one figure can be found with this article online at https://doi.org/10.4196/kjpp.2021.25.4.297.
 XML Download
XML Download