

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Lung cancer despite advancement in the medical field continues to be a major threat to human lives and accounts for a high proportion of fatalities caused by cancers globally [1]. Small cell and non-small cell lung cancers (NSCLC) respectively contribute 15% and 85% of the total pulmonary cancer patients diagnosed worldwide [2]. The rate of 5-year survival for patients with NSCLC is only 15% even after huge advancement in chemo- and radiation therapies for [3]. Development of more efficient treatment strategies for pulmonary cancer are thereby immediately required to improve survival of patients.
Most crucial way to arrest tumor development and inhibit carcinoma progression is via induction of apoptosis. Majority of the researchers are searching for molecules which activate pathways involved in apoptosis and therefore accelerate death of cells. Selective carcinoma cell death is activated by tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) which is an apoptosis inducing ligand related with tumor necrosis factor [4,5]. In activating death TRAIL interacts with receptors namely, DR4 and DR5 present in membranes via its ligands [6]. Moreover, TRAIL also influences decoy receptors including DcR-1 and DcR-2 in addition to osteoprotegerin which are devoid of transmembrane domain [7]. On interacting with DR4 and DR5, TRAIL causes induction of death signal via activation of both initiator as well as effector caspases leading to apoptosis [8,9]. Elevated decoy receptor levels and suppressed DRs are associated with cell resistance to TRAIL mediated apoptosis [9,10]. Besides, excessive FLICE-like inhibitory proteins (cFLIP) along with apoptosis inhibitor proteins (IAPs) also lead to resistance to TRAIL induced death [11].
Four-hydroxy-3-methoxybenzaldehyde (Fig. 1), commonly called as vanillin is present in the seeds of Vanilla planifolia plant. The compound initially used as flavoring agent was latter reported to possess anti-microbial, anti-mutagenic and anti-inflammatory properties in rodent and human models [12-15]. Vanillin targets nuclear factor-κB expression and activity of MMP’s to arrest migration and invasiveness of liver carcinoma cells [16]. In pulmonary cancer cells angiogenesis and development of lamellipodia is inhibited by vanillin by targeting PI3K activation [17-19]. Signals to induce apoptosis in breast and cervical carcinoma cells are initiated by vanillin to inhibit tumor growth [17-19]. The current study investigated vanillin oxime, a derivative of vanillin against lung cancer cells for development of treatment and explored mechanism.
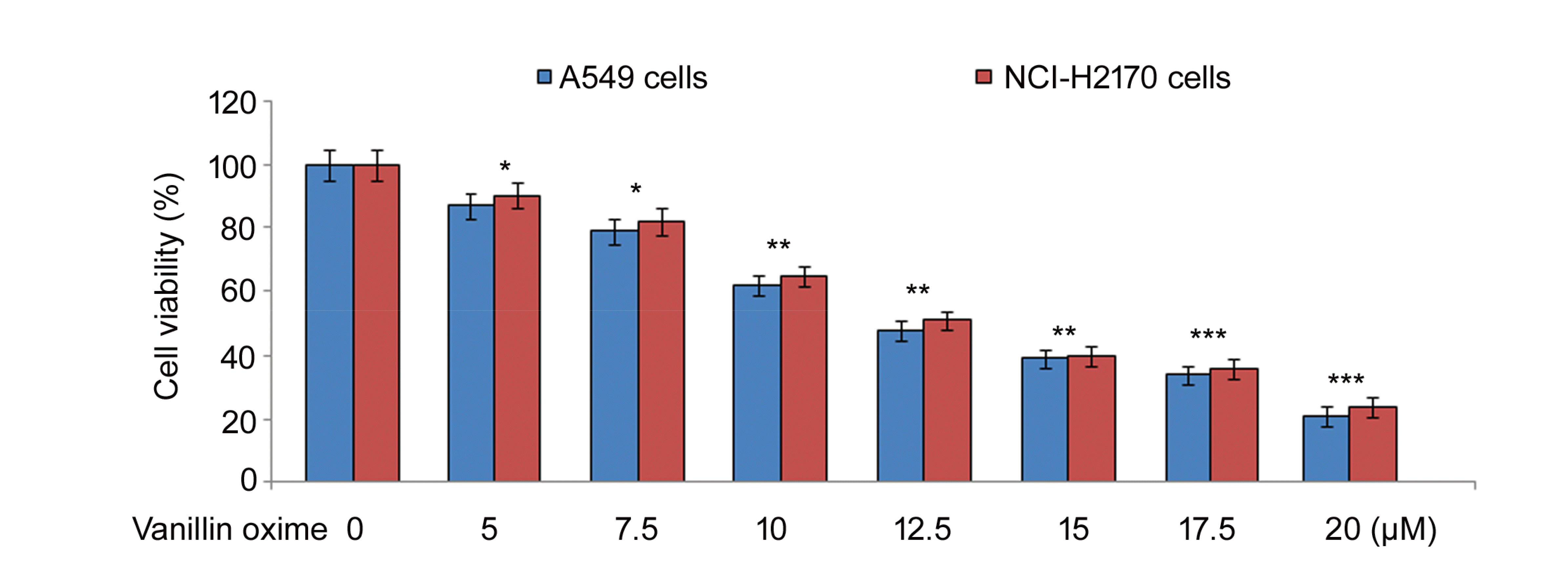 | Fig. 1Effect of vanillin oxime on A549 and NCI-H2170 cells.Vanillin oxime at 5, 7.5, 10, 12.5, 15, 17.5, and 20 µM concentrations was mixed in RPMI to determine A549 and NCI-H2170 cell viability changes at 48 h by MTT assay. MTT, 3-(4,5-dimethylthiazol-2-yl)2,5-diphenyltetrazolium bromide. *p < 0.05 and **p < 0.01 vs. control, ***p < 0.018.
|
Go to : 
METHODS
Drugs and reagents
Vanillin oxime (Cas number S539570; purity 99%) was supplied by Merck Chemicals. Dimethyl sulfoxide (DMSO) and many other chemicals were provided by the Sigma-Aldrich (St Louis, MO, USA).
Cell lines
Lung cancer cell lines (A549 and NCI-H2170) and normal epithelial cells (BEAS-2B) were supplied by the ATCC, Rockville, MD, USA and maintained in RPMI-1640 medium provided with 10% FBS at 37°C in incubator with 5% CO2 atmosphere. Supplementation of medium was made with penicillin (100 units/ml) and streptomycin (100 µg/ml).
Cell viability assay
The A549, BEAS-2B and NCI-H2170 cell viability changes by vanillin oxime were measured using 3-(4,5-dimethylthiazol-2-yl)2,5-diphenyltetrazolium bromide (MTT) assay. Cells maintained for 24 h in RPMI-1640 medium at 2 × 106 cells/well density in 96-well plates were exposed for 48 h to 5, 7.5, 10, 12.5, 15, 17.5, and 20 µM concentrations of vanillin oxime. Medium was changed and then MTT (20 µl; 0.5 mg/ml) solution was poured in each well to incubate cells for 4 h. Afterwards, DMSO (120 µl) was added to the wells and optical density was recorded by Multiskan Spectrometer (Vantaa, Finland) at 574 nm.
Analysis of apoptosis
Vanillin oxime-mediated apoptosis was detected in A549 and NCI-H2170 cells at 48 h of exposure by flow cytometry. Exposure of cells to 12.5 and 15 µM concentrations of vanillin oxime was followed by treatment with annexin V-FITC reagent (5 µl) for 35 min at room temperature. Apoptotic cell fraction was analysed using FACS Calibur flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
Western blotting
The A549 and NCI-H2170 cells were exposed to 12.5 and 15 µM concentrations of vanillin oxime for 48 h. After exposure, incubation was carried with ice-cold (0.5 ml) lysate buffer (consisting of NaCl [5 M], NaVO4 [0.2 M], Nonidet P-40 [10%], EGTA [0.1 M], EDTA [0.5 M], PhCH2SO2F [0.1 M], NaF [1 M], HEPES [1 M] aprotinin [2 µg/ml] and leupeptin [2 µg/ml]) for 45 min. After centrifugation at 12,000 ×g at 4°C protein level in supernatant separated from the lysates was estimated by Bio-Rad assay (Bio-Rad, Hercules, CA, USA). Fractionation of 60 µg protein samples on SDS-PAGE was followed by transfer to PVDF membranes (Millipore, Bedford, MA, USA). Probing of proteins was carried out by over-night membrane incubation at 4°C with primary antibodies. Washing membranes with TBST was followed by incubating the blots for 1 h with anti-rabbit secondary antibodies conjugated to horseradish peroxidase. Detection as well as analysis of the bands was carried out using the ECL western blot analysing system (Biotech Inc., USA). The primary antibodies used were against JNK, p-JNK, caspase-3, -8, -9, CHOP, Bcl-xL, ERK1/2, p-ERK1/2, DR4, DR5, survivin, Bax, cFLIP, xIAP (Cell Signaling Technology, Danvers, MA, USA).
Determination of DR4 and DR5 levels
Mouse anti-human antibodies against DR4 as well as DR5 which are conjugated to the phycoerythrin (Minneapolis, MN, USA) were used for analysis death receptors in A549 and NCI-H2170 cells. The cells at 2 × 106 cells/well distribution were exposed to 12.5 and 15 µM concentrations of vanillin oxime for 48 h. Then incubation with antibodies was done for 50 min at 4°C to analyse receptor expression using flow cytometry.
RT-PCR analysis
To determine levels of CHOP and DR5 mRNA in A549 and NCI-H2170 cells after exposure to 12.5 and 15 µM vanillin oxime RT-PCR was used. From A549 and NCI-H2170 cells total RNA was isolated at 48 h of vanillin oxime exposure by TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Primer sequences used for PCR were as follows: CHOP, forward 5’CAACTGCAGAGATGGCAGC TGA-3’ and reverse 5’-CTGATGCTCCCAATT GTTCAT-3’; DR5, forward 5’-AAGACCCTTGTG CTCGTTGTC-3’ and reverse 5’-GACACATT CGATGTCACTCCA-3’; and GAPDH, forward 5’-GTCTTCACCACCATGGAG3’ and reverse 5’-CCACCCTGTTGCTGTAGC3’.
siRNA transfection
Effect of silencing CHOP and DR5 on A549 and NCI-H2170 cells followed by exposure to vanillin oxime was also studied. Transfection of 30 nM siRNA (oligonucleotides) to A549 and NCI-H2170 cells was performed using lipofectamine 2000 (Invitrogen) as per the manual procedure [20]. Then cells were exposed for 48 h to 12.5 and 15 µM vanillin oxime followed by western blotting to detect changes in CHOP and DR5 expression.
Statistical analysis
The data expressed are mean ± standard deviation of the measurements made in triplicates. To analyse data statistically SPSS software (IBM Corp.) was used. Differences between groups were statistically determined by one-way ANOVA and Duncan’s test (DMRT). The values at p < 0.05 were taken to be statistically significant.
Go to :
RESULTS
Vanillin oxime is cytotoxic
A549 and NCI-H2170 cells were exposed to vanillin oxime at 5, 7.5, 10, 12.5, 15, 17.5, and 20 µM concentrations (Fig. 1). Vanillin oxime in concentration dependent way suppressed A549 and NCI-H2170 cell viabilities at 48 h. At 5 and 20 µM concentrations vanillin oxime exposure reduced A549 cell viability to 87 and 21%, respectively. Viabilities of NCI-H2170 cells were suppressed to 90 and 24%, respectively on exposure to 5 and 20 µM concentrations vanillin oxime. As viabilities of A549 and NCI-H2170 cells were reduced to intermediate level at 12.5 and 20 µM therefore only these two concentrations were chosen for further studies.
Vanillin oxime is apoptotic agent
Vanillin oxime mediated apoptosis in A549 and NCI-H2170 cells at 48 h was detected by Annexin V staining (Fig. 2). Exposure to 12.5 and 15 µM concentrations of vanillin oxime increased A549 and NCI-H2170 cell apoptosis significantly (p < 0.05). Vanillin oxime at 12.5 and 15 µM led to apoptosis in 48.23 and 63.91% A549 cells, respectively. In NCI-H2170 cells apoptosis reached to 45.14 and 59.37%, respectively on exposure to vanillin oxime at 12.5 and 15 µM concentrations.
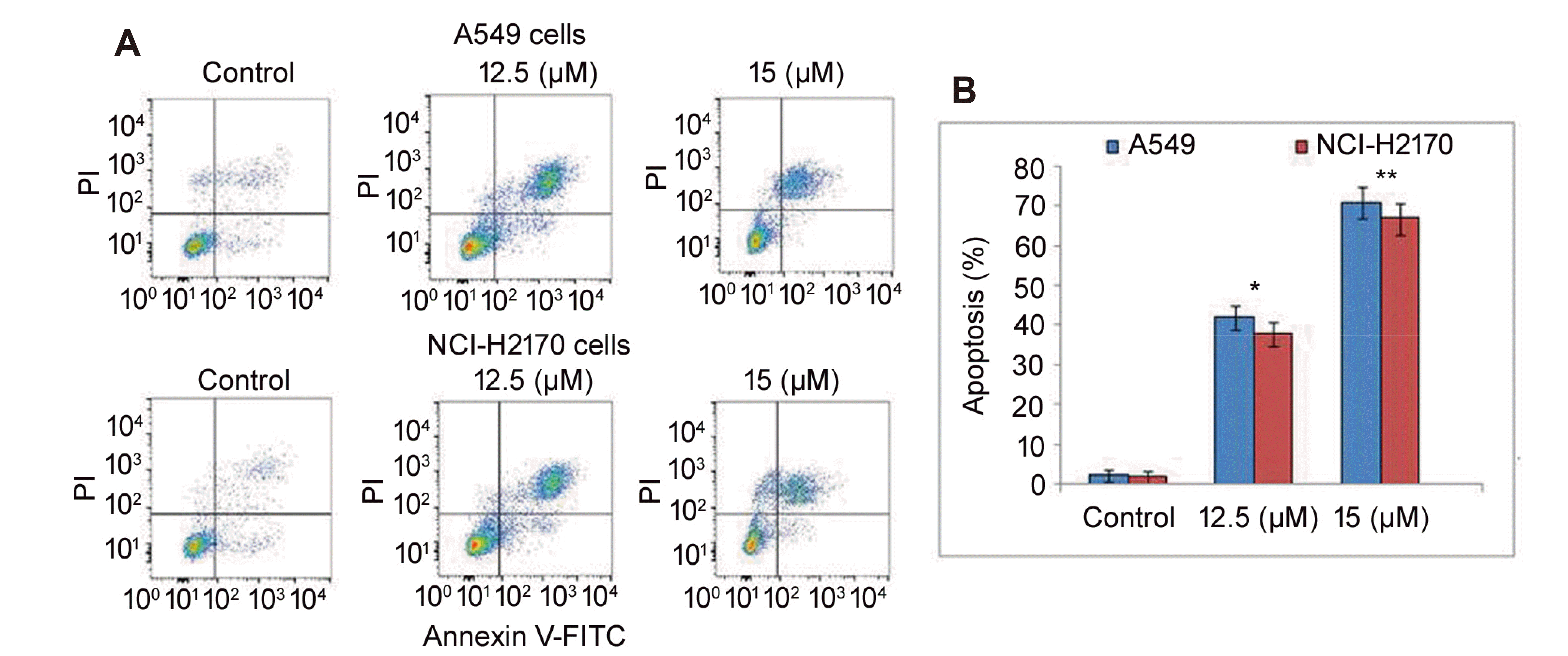 | Fig. 2Effect of vanillin oxime on A549 and NCI-H2170 cell apoptosis.(A) Vanillin oxime at 12.5 and 15 µM concentrations was mixed in RPMI to determine A549 and NCI-H2170 cell apoptosis induction at 48 h by flow cytometry. (B) Quantification of the data obtained from flow cytometry. PI, propidium iodide. *p < 0.05 and **p < 0.01 vs. control.
|
Vanillin oxime promoted pro-apoptotic proteins
In vanillin oxime exposed A549 and NCI-H2170 cells pro-apoptotic protein levels were assayed at 48 h by western blotting (Fig. 3). On exposure to 12.5 and 15 µM concentrations of vanillin oxime elevated cleaved caspase-3 and -9 levels in A549 and NCI-H2170 cells were observed. In vanillin oxime (12.5 and 15 µM) exposed A549 and NCI-H2170 cells Bax level was also higher relative to untreated cells.
 | Fig. 3Effect of vanillin oxime on pro-apoptotic proteins.(A) Vanillin oxime at 12.5 and 15 µM concentrations was mixed in RPMI to determine cleaved caspase-3, -9 and Bax expression in A549 and NCI-H2170 cells at 48 h by Western blotting. (B) Proteins bands were quantified. *p < 0.05 and **p < 0.01 vs. control.
|
Vanillin oxime targeted survival protein expression
Vanillin oxime exposure at 12.5 and 15 µM concentrations suppressed levels of Bcl-2, survivin, Bcl-xL, cFLIP and IAPs proteins in A549 and NCI-H2170 cells (Fig. 4). In control A549 and NCI-H2170 cells Bcl-2, survivin, Bcl-xL, cFLIP and IAPs protein levels were observed to be markedly higher compared to vanillin oxime treated cells. In A549 and NCI-H2170 cells protein expression was determined at 48 h using Western blotting assay.
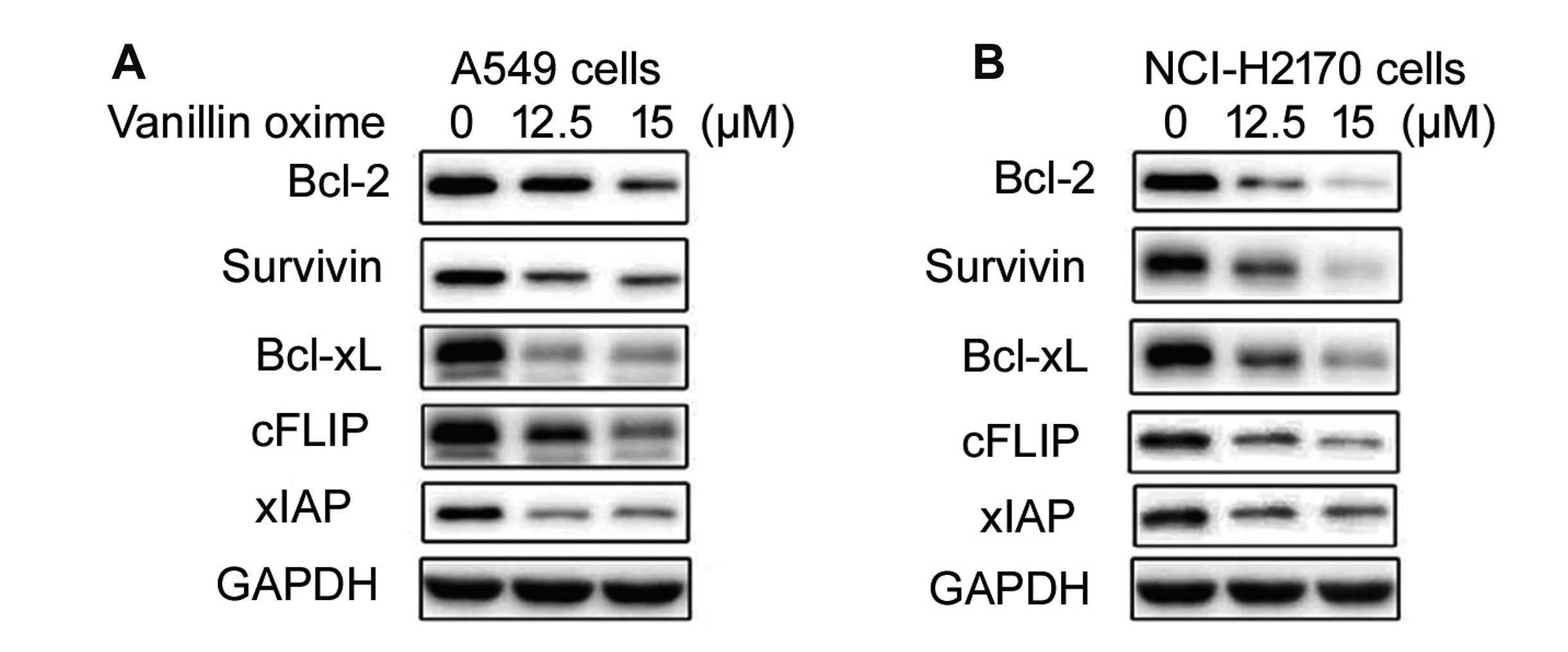 | Fig. 4Effect of vanillin oxime on survival proteins.Vanillin oxime at 12.5 and 15 µM concentrations was mixed in RPMI to assess survival proteins levels in (A) A549 and (B) NCI-H2170 cells by western blotting. In vanillin oxime treated cells protein expression was determined at 48 h using Western blotting assay.
|
Vanillin oxime up-regulated DR4 and DR5
In A549 and NCI-H2170 cells exposure to 12.5 and 15 µM concentrations of vanillin oxime was followed by DR4 and DR5 expression assessment (Fig. 5). Vanillin oxime exposure stimulated significant elevation in DR4 and DR5 levels in A549 and NCI-H2170 cells at 48 h. Promotion in DR4 and DR5 levels was higher on treatment with 15 µM vanillin oxime compared to 12.5 µM concentration.
Vanillin oxime promoted CHOP expression
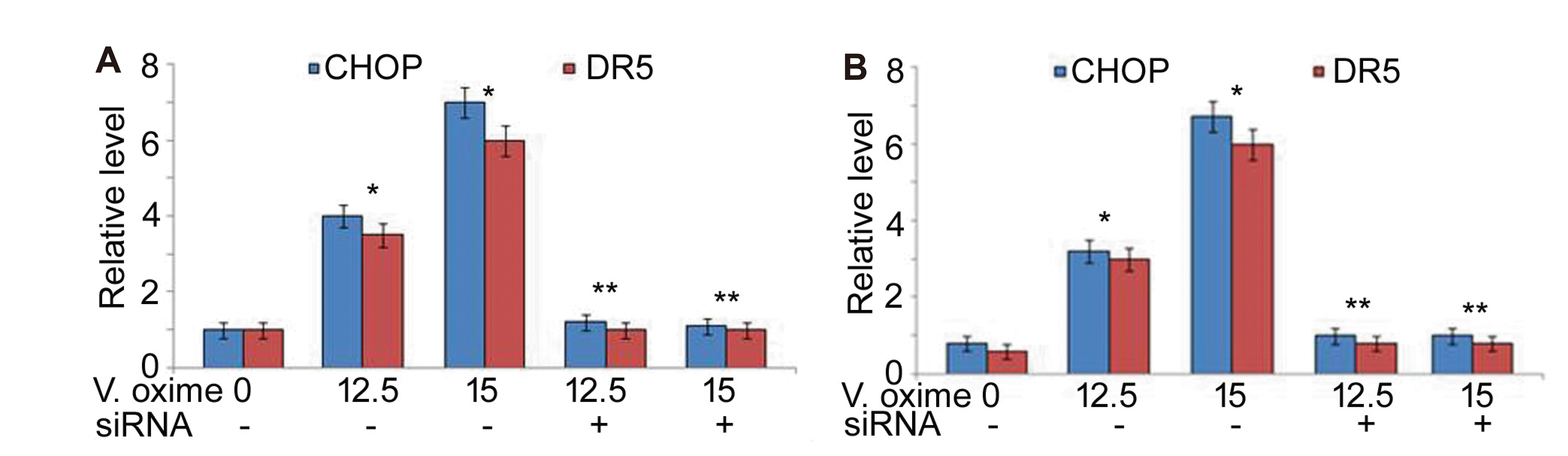
In A549 and NCI-H2170 cells vanillin oxime exposure at 12.5 and 15 µM doses caused significant (p < 0.05) enhancement in CHOP and DR5 expression (Fig. 6). Vanillin oxime exposure at 12.5 and 15 µM doses also elevated CHOP protein level in A549 and NCI-H2170 cells. However, in vanillin oxime exposed A549 and NCI-H2170 cells CHOP and DR5 expression was significantly lowered on CHOP-siRNA transfection (Fig. 7).
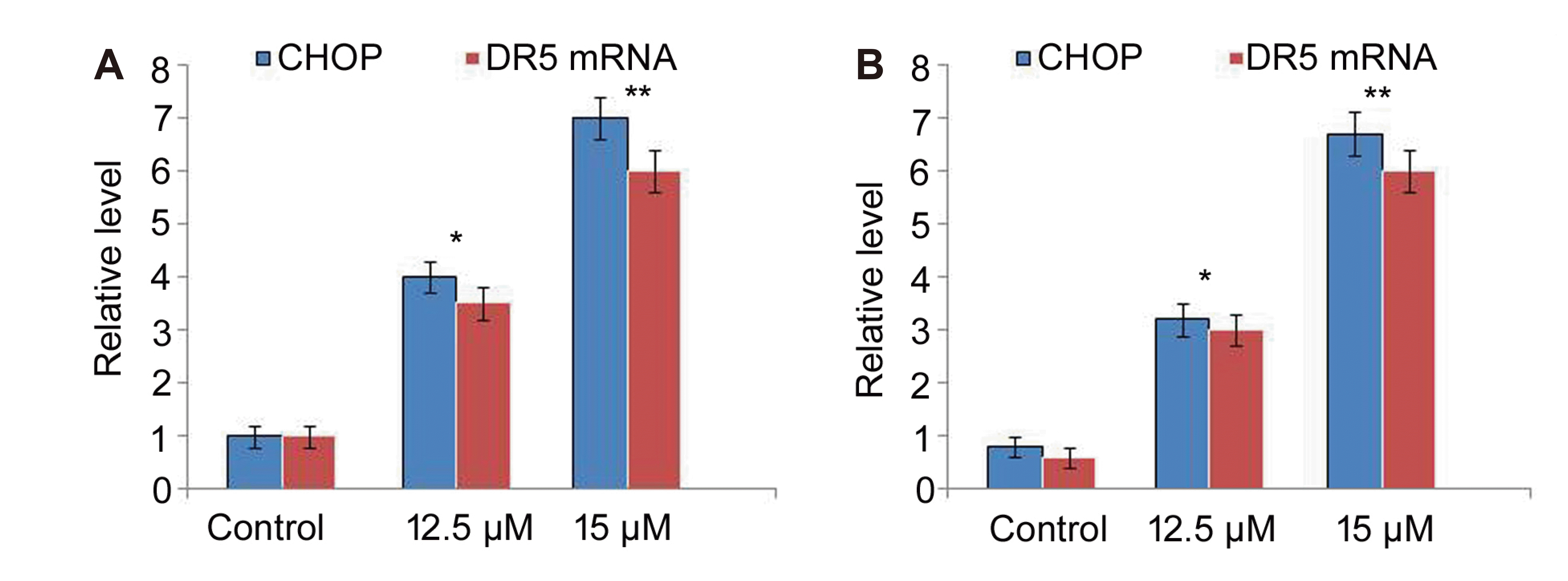 | Fig. 6Effect of vanillin oxime on CHOP and DR5 levels.(A) Vanillin oxime at 12.5 and 15 µM concentrations was mixed in RPMI to assess CHOP and DR5 levels in A549 and NCI-H2170 cells at 48 h by RT-PCR assay. (B) CHOP protein bands were assayed by Western blotting. *p < 0.05 and **p < 0.01 vs. control.
|
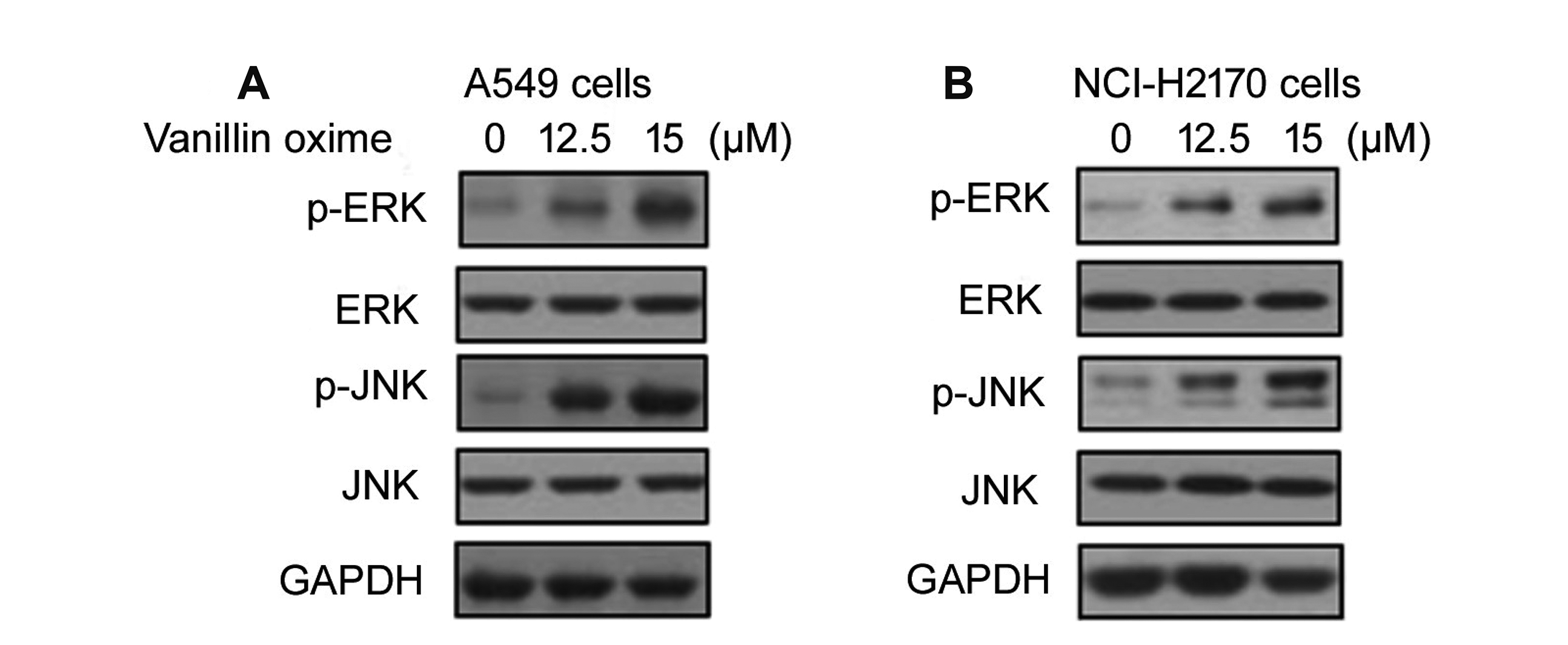
Vanillin oxime induced ERK and JNK activation
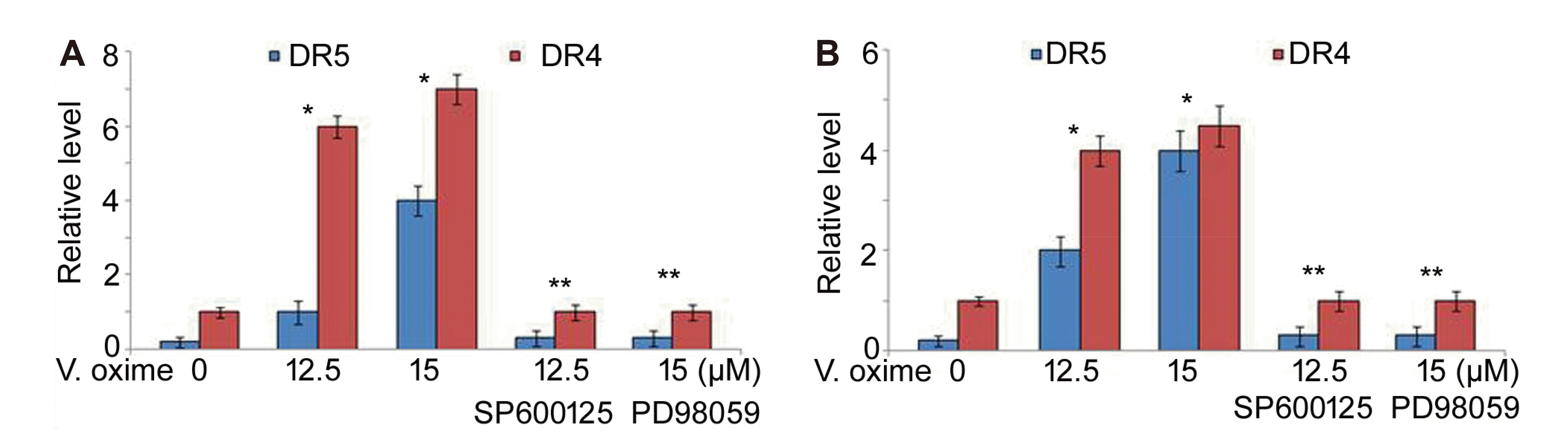
Vanillin oxime exposure of A549 and NCI-H2170 cells at 12.5 and 15 µM doses led to significant (p < 0.05) enhancement in levels of phosphorylated ERK and JNK (Fig. 8). Enhancement in p-ERK and p-JNK levels by vanillin oxime was higher at 15 µM doses relative to 12.5 µM concentration. Treatment of A549 and NCI-H2170 cells with JNK inhibitor (SP600125) as well as ERK1/2 inhibitor (PD98059) led to prominent reduction in vanillin oxime induced DR5 and DR4 levels (Fig. 9).
Go to :
DISCUSSION
Vanillin was shown to possess anticarcinogenic activity in rats and as antimutagen in several in vitro models [13,21]. It potentiates cytotoxicity of many DNA-damaging agents such as cisplatin [22] and mitomycin C [23]. Moreover, vanillin demonstrated anti-metastatic activity for 4T1-mouse mammary carcinoma and inhibited invasiveness and migration of tumor cells in vitro [18]. Resistance to death induced by TRAIL is linked with the higher survival protein levels including, cFLIP, Bcl-2, survivin and Bcl-xL in carcinoma cells [24,25]. Targeting expression of these proteins which are associated with resistance to death induced by TRAIL has prominent potential in cancer treatment strategies. In the present study vanillin oxime suppressed A549 and NCI-H2170 cell viabilities in concentration dependent way. Thus, vanillin oxime was found to be cytotoxic agent for A549 and NCI-H2170 cells from initial MTT studies. Further studies demonstrated that exposure to vanillin oxime increased A549 and NCI-H2170 cell apoptosis significantly (p < 0.05). In vanillin oxime exposed A549 and NCI-H2170 cells caspase-3 and -9 levels were enhanced excessively relative to untreated cells. Moreover, vanillin oxime exposure of A549 and NCI-H2170 cells also raised Bax level significantly. Vanillin oxime exposure suppressed levels of cFLIP, Bcl-2, survivin and Bcl-xL proteins in dose-based way in A549 and NCI-H2170 cells. Therefore, vanillin oxime exhibited toxicity for A549 and NCI-H2170 cells by inducing apoptosis through targeting anti-apoptotic proteins.
Plant derived compounds promote death of carcinoma cells via TRAIL-mediated pathway through targeting survival proteins and elevation of caspases [26,27]. Resistance of cancers to TRAIL is related with dysregulated DR4 and DR5 levels and thereby up-regulation induces cell death [28,29]. In the present study vanillin oxime exposure stimulated significant elevation in DR4 and DR5 levels in A549 and NCI-H2170 cells. Moreover, elevation of DR4 and DR5 levels was higher on exposure to higher doses of vanillin oxime compared to lower concentrations.
The DR5 level is regulated by a transcriptional factor known as C/EBP homologous protein (CHOP) in various cells [30]. The expression of DR5 is promoted by CHOP on biding to the promotor region of DR5 [30]. The present study found that A549 and NCI-H2170 cells expressed significantly (p < 0.05) enhanced CHOP and DR5 mRNA expression on vanillin oxime exposure. Vanillin oxime exposure also elevated CHOP protein level in A549 and NCI-H2170 cells. Moreover, CHOP and DR5 expression was significantly lowered in vanillin oxime exposed A549 and NCI-H2170 cells on CHOP-siRNA transfection.
Stress is the major regulator for CHOP expression which is demonstrated by several studies investigating chemotherapeutic agents [26,27]. Up-regulated levels of CHOP protein and enhanced DR5 expression by oxidative stress has been demonstrated to induce TRAIL mediated apoptosis [26,27]. Among vital factors acting as downstream effectors of oxidative stress in tumor cells include MAPKs. During apoptosis mediated by TRAIL the levels of activated ERK and JNK are prominently higher [26,29]. The current study demonstrated that vanillin oxime exposure of A549 and NCI-H2170 cells led to significant (p < 0.05) enhancement in levels of phosphorylated ERK and JNK. However, treatment of A549 and NCI-H2170 cells with JNK inhibitor (SP600125) as well as ERK1/2 inhibitor (PD98059) led to prominent reduction in vanillin oxime induced DR5 and DR4 levels.
In summary, vanillin oxime inhibits pulmonary cell proliferation via induction of apoptosis through TRAIL mediated pathway. Moreover, ERK/JNK pathway was activated whereas CHOP and DR5 expression was promoted in A549 and NCI-H2170 cells by vanillin oxime treatment. Therefore, vanillin oxime may be studied further to develop treatment for the lung cancer.
Go to :
 XML Download
XML Download