

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Alzheimer's disease (AD) is a neurodegenerative disease characterized by impairment of cognitive functions such as memory and learning [1]. AD is the most common cause of dementia in elderly people, making it a growing public health concern [2]. Due to its long asymptomatic latent period, the onset of AD is difficult to notice until the disease is advanced. Moreover, there is no effective treatment to prevent progression of the disease. These are the reasons why prevention of AD is considered a priority [3]. Diagnostic lesions of the AD brain include extracellular neuritic plaques and intracellular neurofibrillary tangles [4]. Neuritic plaques result in deposition of amyloid-β protein (Aβ) by hydrolyzing the Aβ precursor, also called the amyloid precursor protein (APP). The APP is processed by APP cleaving enzymes such as β- and γ-secretases to generate Aβ [2]. The accumulation of Aβ in synapses attenuates the functions of neurotransmitters, which result in cognitive impairment [5].
Zizyphus jujuba (Zj) is a fruit of Zizyphus jujuba belonging to the Rhamnaceae family [6]. It has been consumed as a medicinal crop for a long time, and used predominantly in Asian countries [7]. Recent studies have reported various biological activities of Zj, including antioxidant and anti-inflammatory functions [89]. It has been reported that Zj attenuates neurotoxicity induced by diabetic neuropathy in PC12 cells, indicating that Zj exerts a protective effect against neurotoxicity [10]. Zj contains several bioactive compounds such as polysaccharides, flavonoids, alkaloids, saponins and triterpenoids [611]. Since these active compounds possess neuroprotective effects, Zj is also expected to provide neuroprotection that emanates from those compounds [1213].
Fermentation is a traditional food processing technique applied for improving storage and functionality of foods. During fermentation, a range of secondary metabolites are produced, some of which possess health-promoting properties [14]. This process also enhances bioavailability of nutrients and bioactive components found in the product [15]. Yeast has been widely utilized for fermentation to manufacture alcoholic beverages and bread [1617]. Numerous studies have used yeast to elevate the functionality of foods by fermentation [1819]. Since Zj has bioactive compounds in the form of glycosides such as flavonol glycosides, absorption into the body can be limited [20]. Yeasts decompose highly polymerized compounds into lower ones during fermentation, thereby reinforcing the absorption of these compounds and increasing the bioactivity of Zj. However, the effects of the Zj and Zizyphus jujuba fermented by yeast (Zj-Y) on cognitive impairments associated with AD are yet to be studied. This study was therefore undertaken to investigate the ability of Zj and Zj-Y on cognitive function in the Aβ-injected AD mouse model.
Go to : 
MATERIALS AND METHODS
Plant materials and ethanoic extracts
Zj was purchased from Miryang, Korea. Totally, 1.15 kg of dried Zj was finely powdered after freeze-drying, followed by extraction with 70% EtOH (1:10, w/v) for 24 h. The extraction process was repeated thrice to obtain the Zj extract. The final yield of ethanoic extract was 721.5 g from the dried Zj. Fermentation of Zj was conducted at 30°C for 48 h by inoculating yeast (Saccharomyces cerevisiae, 106 CFU/mL) into a mixture of Zj powder and distilled water (DW; 1:99, v/v). Fermented Zj was subsequently extracted according to the same method mentioned above.
Regents
Aβ25-35 and Griess reagent were obtained from Sigma Aldrich (St. Louis, MO, USA). Phosphoric acid was procured from Samchun Pure Chemical (Pyeongtaek, Korea), thiobarbituric acid (TBA) from Acros Organics (Geel, Belgium), and 1-Butanol from Yakuri Pure Chemical (Kyoto, Japan). Aspartate aminotransferase (AST) and alanine aminotransferase (ALT) assay kits were purchased from Asan Pharmaceutical Co. (Eumseong, Korea).
Animal care and experiments
ICR mice (5 weeks old, male) were purchased from Orient Inc. (Seongnam, Korea). All mice were housed in plastic cages kept in a controlled environment at 20 ± 2°C temperature, 50 ± 10% humidity, and 12 h light-dark cycle. Totally, 28 mice were divided into 4 groups (n = 7 per group) separated in different cages: normal, control, Zj, and Zj-Y group. Zj and Zj-Y groups were administered with extracts of Zj and Zj-Y at 200 mg/kg/day, respectively. The administration that started after 5 days injection of Aβ25-35 was carried out for 14 days. Normal and control groups were administered with saline. Aβ25-35 was dissolved in diluted DW to achieve a concentration of 5 nM, and incubated at 37°C for 72 h. After aggregation, mice were injected with Aβ25-35 according to the procedure of Laursen and Belknap [21]. Briefly, using 10 μL Hamilton microliter syringe with 26-gauge needle, 5 μL aggregated Aβ25-35 was injected 0.5 mm posterior to the bregma, 1.0 mm rightward to the midline, and at 2.2 mm depth to the surface of the brain. Normal group was injected with saline instead of aggregated Aβ25-35. On day 5 after administering Aβ25-35, Zj and Zj-Y groups were administered extracts of Zj and Zj-Y at 200 mg/kg/day, respectively, for 14 days, whereas normal and control groups received saline. Learning and memory ability of mice was evaluated by behavioral experiments including T-maze test, novel object recognition test, and Morris water maze test. At the end of the experiments, all mice were fasted for 12 h and sacrificed using CO2. Organs such as brain, kidney, and liver were harvested, and all tissue samples were stored at −80°C until analysis. All animal protocols were reviewed and approved by the Pusan National University Institutional Animal Care and Use Committee (PNU-IACUC, approval number: PNU-2010-00056) guidelines for the care of laboratory animals and ethical procedures.
T-maze test
T-maze test was conducted by the method established by Montgomery [22]. The maze is shaped like a ‘T’, having 2 arms at the end of an I-shaped starting box. Walls separating the starting box and arms can be removed during the experiment. In the training period, each mouse was placed in the starting box with a wall separating the right arm. The mouse moved around repeatedly, finding the correct route of the maze. After 24 h, the mouse is capable of moving freely around the maze, and both walls of the right and left arms were eliminated. The percentage of space perceptive ability was evaluated by determining the ratio of the number of left or right route entries to the number of total route entries multiplied by 100.
Novel object recognition test
According to the novel object recognition test procedure published by Bevins and Besheer [23], a black-square box was a field that mice performed the novel object recognition test. The difference between training and test period is the kind of object used. In the training period, 2 identical objects were located in the box, apart from each other. A mouse was placed in the box and allowed to touch objects for 10 min, and the number of touches of each object was counted. In test period, one of the objects was changed into a novel object. Again, the mouse went around and contacted the objects during a 10 min, and number of contacts with each object was recorded simultaneously. The calculation for object cognitive ability percentage is the number of contacts with both objects over the number of contacts with the original object or novel object.
Morris water maze test
The Morris water maze test was performed according to the method suggested by Morris [24], conducted in a round pool 150 cm in diameter and 30 cm in height. A platform was located in one of the 4 quadrants that were distributed equally by imaginary cross lines. The platform was hidden by placing it 1 cm below the water, which was opacified with non-toxic white color paint, and maintained at 22 ± 2°C. Each quadrant had a visual cue that helped the mouse find the hidden platform. The Morris water maze test consisted of a training period for 3 days and testing period for 1 day; 3 trials per day were performed during the training period. During training, the trial was initiated by placing a mouse on the water surface in a random quadrant, except the one containing the platform. The mouse found the platform by swimming freely for a maximum of 60 s. If the mouse failed to find the platform in time, it was led to it and allowed to stay on the platform for 15 s. The following day, the same trial was conducted.
Measurement of malondialdehyde (MDA)
The measurement of MDA in tissues was conducted by applying the procedure described by Mihara and Uchiyama [25]. Tissues obtained by dissecting mice were homogenized with saline and mixed with 1% phosphoric acid and 0.67% TBA solution. Following 45 min boiling and subsequent cooling on ice, 2 mL of 1-butanol was added to the mixture, and the solution was centrifuged at 3,000 rpm for 10 min. The absorbance value was measured at excitation 520 nm and emission 535 nm. The level of lipid peroxidation was calculated with a standard curve derived from MDA.
Measurement of nitric oxide (NO)
NO levels in tissues were measured by the method of Green et al. [26] and Schmidt et al. [27]. Tissues homogenized in saline were mixed with DW. Griess reagent was added to the mixture, and the solution was incubated at room temperature for 30 min. Absorbance was measured at 540 nm. A standard curve was used for calculating the NaNO2 production yield.
Measurement of AST and ALT
The AST and ALT concentrations were determined according to the method proposed by Reitman and Frankel [28]. The reagent for AST and ALT was preincubated at 37°C, and subsequently mixed with liver tissue homogenized in saline. After incubating the mixtures at 37°C for 60 min and 30 min for AST and ALT, respectively, 2,4-dinitrophenylhydrazine was added to induce a color reaction, after which 10 mL of 0.4 N NaOH was added and incubated for 10 min at room temperature. Enzyme activities of AST and ALT were measured at absorbance 490 nm, and quantified based on a standard curve.
Statistical analysis
Data in this study were analyzed using the SPSS software (IBM Corporation, Armonk, NY, USA), and are expressed as means ± SD. Statistical differences between the normal, control, Zj, and Zj-Y groups were determined by one-way analysis of variance followed by Duncan's post hoc test. In the T-maze and novel object recognition test, the perceptive ability between training and test sessions were compared by Student's t-test. Values of P < 0.05 are considered significant.
Go to :
RESULTS
Effects of Zj and Zj-Y on spatial cognition ability in T-maze test
We examined the effects of Zj and Zj-Y on Aβ25-35-induced memory impairment of mice by applying the T-maze test; data is presented in Fig. 1. The Aβ25-35-injected control group showed no significant difference in their mode of entering the new and old routes, whereas the normal group showed a preference for the new route over the old route. This suggests that injecting Aβ25-35 deteriorates the short-term memory in mice. Conversely, the Zj and Zj-Y groups administrated Zj or fermented Zj showed significantly higher preference for utilizing the new route. Furthermore, preferences for the new route were significantly increased in the Zj and Zj-Y groups, as compared to the normal group mice. Our results indicate that both Zj and Zj-Y administered groups have improved spatial cognition, as compared to the Aβ25-35-injected group.
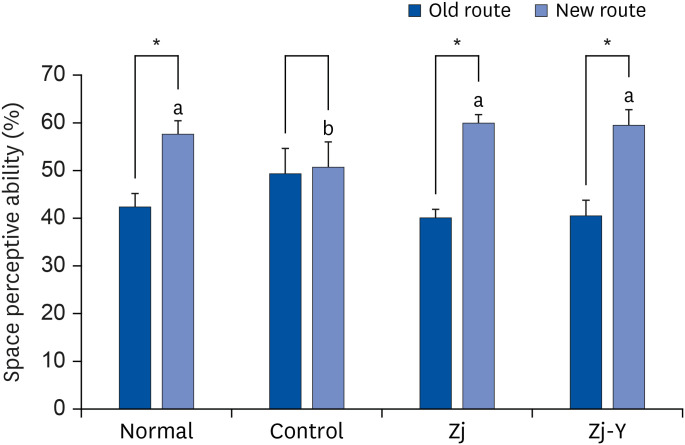 | Fig. 1Effects of Zj and Zj-Y on spontaneous alternation behavior in the T-maze test. Normal: saline injection + oral administration of saline for 14 days. Control: Aβ25-35 injection + oral administration of saline for 14 days. Zj: Aβ25-35 injection + oral administration of Zj (200 mg/kg/day) for 14 days. Zj-Y: Aβ25-35 injection + oral administration of Zj-Y (200 mg/kg/day) for 14 days. Values are mean ± SD.Zj, Zizyphus jujuba; Zj-Y, Zizyphus jujuba fermented by yeast; Aβ25-35, amyloid beta25-35.
*The space perceptive abilities for old and new routes are significantly different as determined by Student's t-test (P < 0.05); a,bMeans with the different letters are significantly different among groups by Duncan's multiple range test (P < 0.05).
|
Effects of Zj and Zj-Y on object recognition ability in novel object recognition test
To evaluate the effects of Zj and Zj-Y on impaired object recognition ability, we carried out the novel object recognition test; results are presented in Fig. 2. There was no significant difference in the recognition ability for old and novel objects in the Aβ25-35-treated control group. However, the groups administered Zj and Zj-Y spent more time around the novel objects, and touched the more than familiar object, with the Zj-Y administered group paying significantly more attention to novel objects than the Zj administered mice. This indicates that administration of Zj and Zj-Y recovers impairment of the object recognition ability in the AD mouse model, and Zj-Y is more effective in increasing object recognition ability compared to Zj.
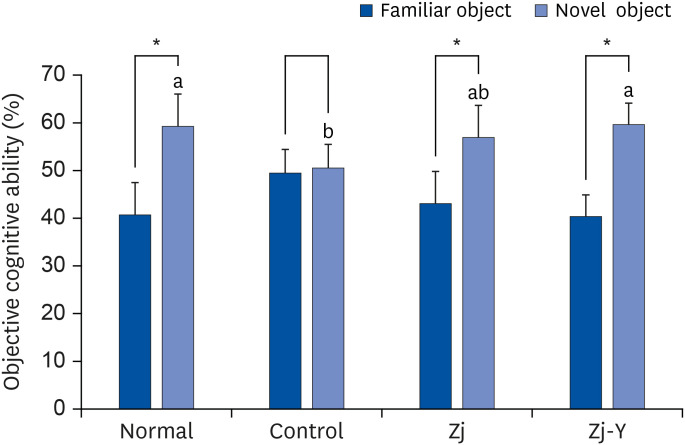 | Fig. 2Effects of Zj and Zj-Y on recognition memory in the novel object recognition test. Normal: saline injection + oral administration of saline for 14 days. Control: Aβ25-35 injection + oral administration of saline for 14 days. Zj: Aβ25-35 injection + oral administration of Zj (200 mg/kg/day) for 14 days. Zj-Y: Aβ25-35 injection + oral administration of Zj-Y (200 mg/kg/day) for 14 days. Values are mean ± SD.Zj, Zizyphus jujuba; Zj-Y, Zizyphus jujuba fermented by yeast; Aβ25-35, amyloid beta25-35.
*The space perceptive abilities for old and new routes are significantly different as determined by Student's t-test (P < 0.05); a,bMeans with the different letters are significantly different among groups by Duncan's multiple range test (P < 0.05).
|
Effects of Zj and Zj-Y on reference working memory in Morris water maze test
Morris water maze test was conducted to investigate whether Zj and Zj-Y attenuate the Aβ25-35-induced long-term memory functional damage. As shown in Figs. 3 and 4, the Aβ25-35-injected mice take longest time to reach the hidden platform, whereas groups administrated Zj and Zj-Y (especially Zj-Y) take considerably less time to reach the hidden platform, as compared to the control group. This indicates that mice injected with Aβ25-35 have damaged learning and memory functions, but administration of Zj and Zj-Y alleviates the impaired functions. Especially, Zj group tended to spend less time finding the platform than the normal and Zj-Y groups.
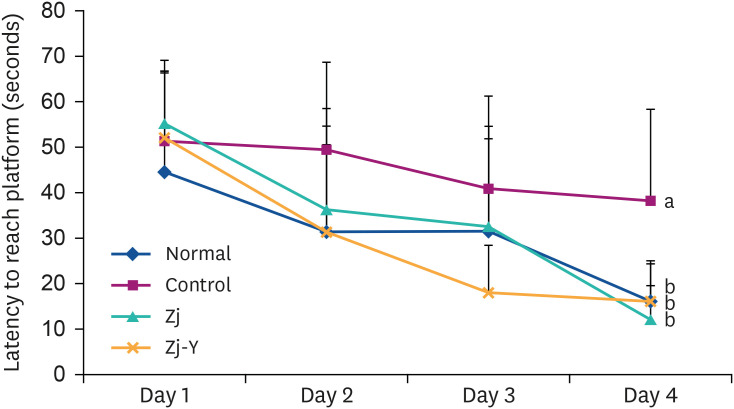 | Fig. 3Effects of Zj and Zj-Y on escape latency to platform in the Morris water maze test. Normal: saline injection + oral administration of saline for 14 days. Control: Aβ25-35 injection + oral administration of saline for 14 days. Zj: Aβ25-35 injection + oral administration of Zj (200 mg/kg/day) for 14 days. Zj-Y: Aβ25-35 injection + oral administration of Zj-Y (200 mg/kg/day) for 14 days. Values are mean ± SD.Zj, Zizyphus jujuba; Zj-Y, Zizyphus jujuba fermented by yeast; Aβ25-35, amyloid beta25-35.
a,bMeans with the different letters are significantly different among groups by Duncan's multiple test (P < 0.05).
|
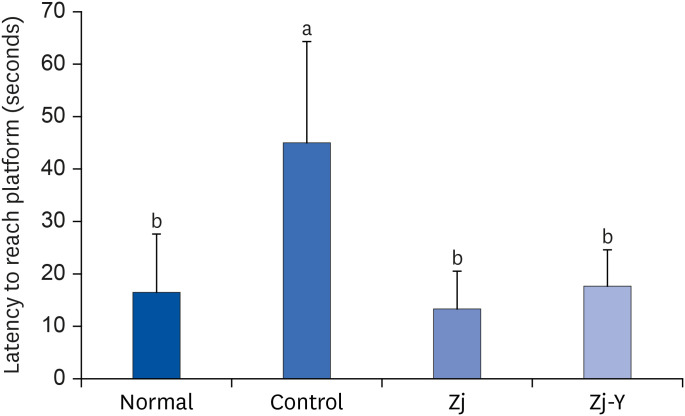 | Fig. 4Effects of Zj and Zj-Y on latency to reach hidden platform in the Morris water maze test. Normal: saline injection + oral administration of saline for 14 days. Control: Aβ25-35 injection + oral administration of saline for 14 days. Zj: Aβ25-35 injection + oral administration of Zj (200 mg/kg/day) for 14 days. Zj-Y: Aβ25-35 injection + oral administration of Zj-Y (200 mg/kg/day) for 14 days. Values are mean ± SD.Zj, Zizyphus jujuba; Zj-Y, Zizyphus jujuba fermented by yeast; Aβ25-35, amyloid beta25-35.
a,bMeans with the different letters are significantly different by Duncan's multiple range test (P < 0.05).
|
Effects of Zj and Zj-Y on MDA generation in mice tissues
MDA levels of the brain, liver, and kidney in mice were measured to evaluate the effects of Zj and Zj-Y against lipid peroxidation (Fig. 5). The control group showed significantly higher MDA levels in the brain, liver, and kidney, as compared to levels obtained in the normal group. However, administrations of Zj or Zj-Y resulted in decreased MDA levels in all 3 tissues, as compared to the control group. In particular, Zj-Y exerted a strong inhibitory effect against MDA generation, thus indicating that fermentation of Zj leads to increased antioxidative activity.
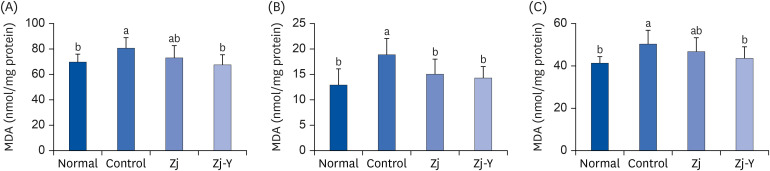 | Fig. 5Effects of Zj and Zj-Y against MDA generation in the brain (A), liver (B), and kidney (C) induced by Aβ25-35. Normal: saline injection + oral administration of saline for 14 days. Control: Aβ25-35 injection + oral administration of saline for 14 days. Zj: Aβ25-35 injection + oral administration of Zj (200 mg/kg/day) for 14 days. Zj-Y: Aβ25-35 injection + oral administration of Zj-Y (200 mg/kg/day) for 14 days. Values are mean ± SD.Zj, Zizyphus jujuba; Zj-Y, Zizyphus jujuba fermented by yeast; Aβ25-35, amyloid beta25-35; MDA, malondialdehyde.
a,bMeans with the different letters are significantly different by Duncan's multiple test (P < 0.05).
|
Effects of Zj and Zj-Y on NO generation in mice tissues
Fig. 6 presents the NO levels in the brain, liver, and kidney of mice. As compared to the normal group, significantly higher levels were obtained in the Aβ25-35-injected group. However, administration of Zj or Zj-Y resulted in suppressed NO levels in the Aβ25-35 treated mice. Moreover, decreased NO formation in the brain, liver, and kidney was more prominent after Zj-Y administration as compared to Zj administration.
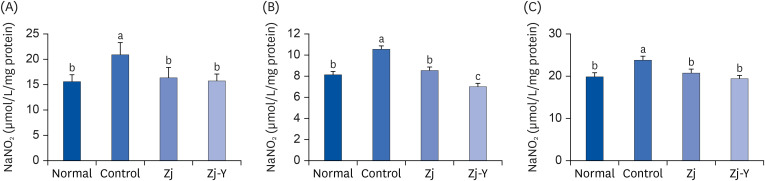 | Fig. 6Effects of oral Zj and Zj-Y on Aβ25-35-induced nitric oxide formation in the brain (A), liver (B), and kidney (C). Normal: saline injection + oral administration of saline for 14 days. Control: Aβ25-35 injection + oral administration of saline for 14 days. Zj: Aβ25-35 injection + oral administration of Zj (200 mg/kg/day) for 14 days. Zj-Y: Aβ25-35 injection + oral administration of Zj-Y (200 mg/kg/day) for 14 days. Values are mean ± SD.Zj, Zizyphus jujuba; Zj-Y, Zizyphus jujuba fermented by yeast; Aβ25-35, amyloid beta25-35.
a-cMeans with the different letters are significantly different by Duncan's multiple test (P < 0.05).
|
Effects of Zj and Zj-Y on ALT and AST
As shown in Table 1, administration of Aβ25-35 significantly elevated the ALT and AST activities in serum, as compared with normal group. However, Zj and Zj-Y exposure resulted in degradation of the ALT and AST activity in the Aβ25-35-exposed mice. The values of AST and ALT observed in the present study were lower than those reported by a previous study using Aβ25-35 [29]. Furthermore, it has been reported that pretreatment with Zj followed by hepatic injury ameliorates hepatic function in mice [30]. These results indicate that Zj and Zj-Y induce no hepatoxic effects against Aβ25-35.
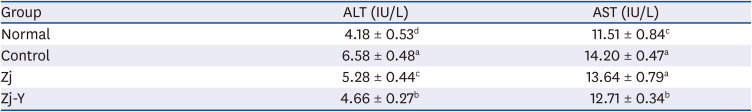
Table 1
Effects of Zj and Zj-Y on ALT and AST
| Group | ALT (IU/L) | AST (IU/L) |
|---|---|---|
| Normal | 4.18 ± 0.53d | 11.51 ± 0.84c |
| Control | 6.58 ± 0.48a | 14.20 ± 0.47a |
| Zj | 5.28 ± 0.44c | 13.64 ± 0.79a |
| Zj-Y | 4.66 ± 0.27b | 12.71 ± 0.34b |
Values are mean ± SD. Normal: saline injection + oral administration of saline for 14 days. Control: Aβ25-35 injection + oral administration of saline for 14 days. Zj: Aβ25-35 injection + oral administration of Zj (200 mg/kg/day) for 14 days. Zj-Y: Aβ25-35 injection + oral administration of Zj-Y (200 mg/kg/day) for 14 days.
Zj, Zizyphus jujuba; Zj-Y, Zizyphus jujuba fermented by yeast; ALT, alanine aminotransferase; AST, aspartate aminotransferase; Aβ25-35, amyloid beta25-35.
a-dMeans with the different letters are significantly different (P < 0.05) by Duncan's multiple test.
![]()
Go to :
DISCUSSION
Aβ accumulation in the brain is a neuropathological hallmark in AD patients. Ageing and environmental risk factors cause an imbalance between production and clearance of Aβ, resulting in Aβ generation and deposition. The deposition of Aβ results in oxidative stress and neuronal dysfunction [31]. Aβ25-35 is the core fragment of full-length Aβ, possessing numerous characteristics of the full-length Aβ. Aβ25-35 reveals cytotoxic properties similar to full-length Aβ, therefore it is constituted to represent a biologically active region of Aβ [32]. Previous studies report that injecting Aβ25-35 in mice triggers cognitive dysfunction [3334]. Hence, the Aβ25-35-injected mouse model is extensively used for studying AD, and has been applied in the current study.
Food fermentation plays an important role in food preservation. Fermented foods such as wines and ciders have a low pH and high alcohol content. This environment is capable of hindering microbial growth (including molds and bacteria) which contribute to spoilage of foods [35]. Furthermore, fermentation is known to improve biological activities of foods. For example, fermentation of soybean extract with yeast increases the antioxidant properties [36]. Bokbunja fermented by yeast also imparts superior antioxidant properties and contains more total phenolic content than the unfermented material [18]. Yeast-fermented aged black garlic has increased effective bioactivity against high-fat diet mice, as compared to the non-fermented aged black garlic [19]. Similarly, Zj has been utilized for fermented food products including alcoholic beverages, vinegar, and bread [37]. Addition of Zj extract stimulates acid production of lactic acid bacteria in yoghurt, indicating that Zj imparts beneficial effects to human intestine [38]. Zj fermented wines have increased antioxidant characteristics as well as overall acceptability [39]. These studies indicate that Zj can be a highly useful fermented product with improved functionality. However, no studies have reported the protective effect of fermented Zj against Aβ25-35-induced AD. Therefore, we first carried out total phenol analysis and free radical scavenging assays to compare the effects of various microorganisms on Zj fermentation, and found out that Zj-Y showed highest total polyphenol contents and radical scavenging activity [40]. Consequently, we selected yeast as a microbe to ferment Zj in the current study. Based on our previous results, we evaluated whether Zj and Zj-Y alleviate cognitive and memory deficits in the Aβ25-35-injected AD mouse model. Zj or Zj-Y were orally administrated to Aβ25-35-injected mice for 14 days. Previous studies have determined the beneficial effect of Zj in carbon tetrachloride-induced hepatic injury in mice after oral administration of 200 mg/kg/day [30]. Moreover, doses up to 500 mg/kg/day did not show any toxicity [41]. In this study, we administered Zj-Y with the same concentration as Zj (200 mg/kg/day) to assess whether fermentation affects the beneficial effect of Zj in an AD model, by evaluating behavior tests including T-maze, novel object recognition, and Morris water maze test, as well as their effects on lipid peroxidation and NO production.
The T-maze and novel object recognition tests evaluate the working memory, and they are defined as short-term memory for an object, stimulus, or location and that is trial dependent [4243]. These tests assess the natural preference of the mouse, which implies that they spend more time to explore the novel route/object than the familiar one without any reinforcement [44]. In the both tests, the Aβ25-35-injected control group showed no significant difference between the number of entries into the new or old route/object. It indicates that exposure to Aβ25-35 induces impairment of short-term working memory function. However, in T-maze test, the groups administered Zj and Zj-Y showed a higher tendency to explore the novel route rather than the old route. In the novel object recognition test, Zj, and especially Zj-Y, induced a significantly higher preference for the novel object, compared to the Aβ25-35-injected control group. These results indicate that Zj and Zj-Y improve the object recognition ability impaired by injection of Aβ25-35 in mice.
The Morris water maze test is widely applied to examine the spatial learning and reference memory by assessing repeated trials and preference for the platform area [4546]. In the current study, the Aβ25-35-injected control group did not decrease escape latency by trials. These results evidence that exposure to Aβ25-35 impairs spatial learning and memory in mice. However, the Zj and Zj-Y administered groups had a shorter time for reaching the platform to escape the water maze. A previous study reported that Zj improves the spatial learning and memory in Morris water maze test that was impaired in rats by bilateral electric lesions of the nucleus basalis of Meynert [47]. Thus, our results indicate that Zj and Zj-Y impart a protective effect on learning and memory functions against Aβ25-35.
Polyunsaturated fatty acids are abundant in the brain; as a result, the brain is vulnerable to free radical attack [48]. Oxidative stress on polyunsaturated fatty acids cause lipid peroxidation, generating reactive aldehydes such as MDA [4849]. Since MDA is toxic to neurons, they have been in focus as a biomarker of lipid peroxidation in the kidney and liver as well as in the brain [5051]. For this reason, MDA is also considered an index of lipid peroxidation in the AD model [51]. Our data demonstrates that Aβ25-35 induces lipid peroxidation in the brain, liver, and kidney of mice, but administration of Zj and Zj-Y results in significantly decreased MDA levels. These results indicate that Zj, and especially Zj-Y, exert a protective effect against lipid peroxidation that is induced by injection of Aβ in mice.
NO acts as a modulator of neural signal. However, at higher concentrations, it leads to neurotoxic effects causing oxidative damage [5253]. Aβ induces NO production by modulating the Ca2+ concentration or interacting with glial cells. Peroxynitrite results from responding NO and reactive oxygen species, leading to neuronal damage [54]. In this study, NO was overproduced in the brain, liver and kidney subsequent to Aβ25-35 injection, but administration of Zj and Zj-Y significantly prevented NO generation. In our previous study, we found that Zj and Zj-Y inhibit NO production in RAW 264.7 cells treated with sodium nitroprusside or lipopolysaccharide and interferon-δ [55]. Thus, we conclude that Zj and Zj-Y act as antioxidants against oxidative damage in the Aβ25-35-injected mouse model.
AST and ALT are hepatic enzymes found in the cytosol of the hepatocyte. Injury to hepatocytes results in leakage of these enzymes, resulting in increased serum AST and ALT levels [5657]. Due to their differing plasma half-life, both are used for evaluating hepatic disorders [57]. Our data reveals that exposure to Aβ25-35 resulted in hepatotoxicity, but administration of Zj and Zj-Y reduced the serum AST and ALT levels. It has been reported that Zj significantly decreases AST and ALT levels in the carbon tetrachloride-induced hepatic injury mice model [30]. These results indicate that Zj and Zj-Y have no toxicity, but improve hepatic damage.
In the current study, we confirmed the effects of administering Zj and Zj-Y in the AD mouse model. Our results in the behavioral tests indicate that treatment with Zj and Zj-Y improves the learning impairment and memory function. Moreover, oxidative stress is prevented by inhibiting lipid peroxidation and NO production. We further determined the cognitive improvement of Zj via antioxidative activity. Our study is in agreement with a previous report that Zj inhibits oxidative stress by decreasing the reactive oxygen species formation against oxidative injury in PC12 neuronal cells [58]. Zj is also known to improve cognitive abilities through various mechanisms. It has been reported that the seed of Zj attenuates Aβ-induced long-term potentiation deficit in the hippocampal tissue from mouse by regulating brain-derived neurotrophic factor (BDNF)/tropomyosin receptor kinase B signaling [59]. Saponin isolated from Zj reduced memory impairment induced by Aβ injection through amelioration of choline acetyltransferase and inflammation in a mouse model [60]. These effects of Zj indicate the potential as anti-AD agent against Aβ. Furthermore, Zj promotes neuronal differentiation that could be a means to protect neurodegenerative diseases such as AD. Previous studies have reported that treatment with Zj stimulates neurite outgrowth which results from overexpression of neurofilaments in cultured PC12 cells [61], and also up-regulates neurotrophic factors such as BDNF and nerve growth factor in astrocytes [62]. Simultaneously, increased markers for cell proliferation and neuroblast differentiation have been observed subsequent to administration of Zj, suggesting that Zj enhances neurogenesis in middle-aged mice [63]. Cyclic adenosine monophosphate, one of the bioactive components of Zj, exerts a role in neuronal differentiation [64]. These findings indicate that Zj and its bioactive components are potential candidates for developing health supplements and for the treatment of AD.
Our current study results indicate that Zj-Y imparts greater recovery of the working memory and stronger protective effect against oxidative stress than Zj. Previous studies have reported that fermentation of plant resources increases the total phenolic compound content, indicating that fermentation can be utilized as a processing method for increasing the physiological activity by increasing the absorption and bioavailability in the body [65]. For example, fermentation of Citrus grandis Osbeck peel by yeast increased the contents of an unknown compound and improved the antioxidant activities; this is thought to be due to conversion of flavonoid glycosides (such as naringin and neohesperidin) to other forms [66]. According to our previous study, the total polyphenol content of Zj was increased via fermentation. In particular, the content of total polyphenol increased 1.9-fold through fermentation using yeast, as compared to non-fermented Zj, which was the highest value obtained compared to other microorganisms [8]. Furthermore, Zj-Y showed strongest effect against oxidative stress and hair loss, suggesting that there is a positive correlation between the total polyphenol content and physiological activity [8]. Since Zj also has flavonoid glycosides (including quercetin 3-O-robinobioside and quercetin 3-O-rutinoside [rutin]), the increase in total polyphenol content and functionality by yeast fermentation is consistent with previous studies [20]. These observations with our findings suggest that Zj-Y has reinforced neuroprotective effects, which is associated with increased biologically active compounds due to fermentation. In conclusion, we propose that Zj, and especially Zj-Y, have the potential to improve Aβ-induced cognitive impairment via oxidative damage, and should be considered potential dietary agents for the prevention and treatment of AD.
Go to :
 XML Download
XML Download