

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Bone mass integrity is maintained by the balance between osteoblastic bone formation and osteoclastic bone resorption [1,2]. Bone forming osteoblasts are recruited from bone marrow-derived mesenchymal stem cells (BMSCs) via stages of cellular commitment, differentiation, osteoblast maturation and matrix mineralization [3,4]. Osteoclast, the multinucleated giant cells that resorb bone are derived from monocyte-macrophage lineage via a process, including cell proliferation, differentiation and fusion [5].
BMSCs are a group of non-hematopoietic stem cells that present in bone marrow niche and have the potential to differentiate into osteoblast, adipocyte, chondrocyte and other mesoderm-derived cell lineages [3,5-7]. Thus, identifying molecules that can direct the differentiation fate of BMSCs into osteogenic lineage will increase bone formation [8,9].
The Mediterranean diet is rich with a wide variety of phytochemicals that are known to have many pharmacological properties for improving chronic diseases including cardiovascular diseases, diabetes, inflammation, and cancer [10,11]. Carnosol (C20H26O4) is a phenolic diterpene phytochemical extracted mainly from Rosmarinus officinalis (rosemary), which representing the main constituents of antioxidant compounds of rosemary leaves [12,13]. Carnosol was reported for its wide range of bioactivities in vitro and in vivo, including antibacterial, anti-inflammatory, anticarcinogenic, neuroprotective and antioxidative effects [13-16]. Recently, carnosol was found to exert an antiaging effect in C. elegans by significantly inhibiting the reactive oxygen species accumulation under normal or oxidative stress condition [17].
In terms of bone biology, Carnosol was reported to inhibit the catabolic mediator’s gene expression of cartilage breakdown in chondrocytes via its anti-inflammatory effect to inhibit IL-6 production by osteoblast [18]. In addition, carnosol significantly inhibited the expression of pro-inflammatory cytokines and the nuclear translocation of the receptor activator of nuclear factor‐κB (NF-κB) in human primary chondrocytes stimulated by IL-1β [19,20], and to suppress the RANKL-induced osteoclast formation [21]. However, the potential role of carnosol in osteoblast differentiation of BMSCs was unknown.
In this study, we investigated the effect of carnosol on the differentiation of BMSCs into osteoblast lineage and adipocyte lineage. Our results demonstrated the dual effect of carnosol on stimulating osteogenesis and inhibiting adipogenesis via BMP-dependent mechanism.
METHODS
Cell cultures and reagents
Experiments were approved by the Standing Research Ethics Committee, King Faisal University, Saudi Arabia with approval number KFU-REC/2020-01-12 .
Primary mouse BMSCs were isolated and cultured from 8-weeks-old male C57BL/6J mice as previously reported by our group [22]. In brief, bone marrow cells were flushed out from mouse tibia and femur, and then filtrated through a 70-μm nylon mesh filter. Cells were cultured in RPMI-1640 medium supplemented with 12% FBS (Gibco Invitrogen, Grand Island, NY, USA), 12 μM L-glutamine (Gibco Invitrogen) and 1% penicillin/streptomycin (P/S) (Gibco Invitrogen). After 24 h, non-adherent cells were removed and cultured in 60 cm2. Medium was changed every 3–4 days and cells were washed and regularly sub-cultured.
Carnosol and LDN-193189 were purchased from (Sigma-Aldrich Chemie Gmbh, Munich, Germany).
Cell toxicity assay
Cell toxicity was determined by measuring cell viability using MTT cell proliferation assay kit after 3 days of treatment with carnosol (Sigma-Aldrich) according to the manufacturer's instructions kit. Cells were incubated with MTT solution to metabolize to formazan and absorbance was measured at a wavelength of 550 nm.
Osteoblast differentiation
mBMSCs were induced to differentiate into osteoblasts with osteogenic induction medium (OIM) consists of in α-minimum essential medium (α-MEM; Gibco) supplemented with 10% FBS, 100 U/ml of penicillin, 100 mg/ml of streptomycin, 10 mM β-glycerol-phosphate (Sigma-Aldrich ApS, Hamburg, Germany), and 50 mg/ml of vitamin C (Sigma-Aldrich ApS). Medium changed every third day during the differentiation course.
Adipocyte differentiation
Mouse bone marrow-derived mesenchymal stem cells (mBMSCs) were induced to differentiate into adipocytes with adipogenic-induction medium (AIM) consists of low glucose DMEM supplemented with 9% horse serum, 450 μM 1-methyl-3-isobutylxanthine (IBMX), 250 nM dexamethasone, 5 μg/ml insulin (Sigma-Aldrich) and 1 μM rosiglitazone (BRL 49653; Cayman Chemical, Ann Arbor, MI, USA). Medium changed every third day during the differentiation course.
Quantitative of alkaline phosphatase (ALP) activity
Cells were cultured in 96 well plate and induced with OIM as mentioned above. ALP activity was determined by incubating the cells with 1 mg/ml of P-nitro phenyl phosphate in 50 mM NAHCO3 and 1 mM MgCl2 buffer (pH 9.6) at 37°C for 20 min. The reaction was stopped by the addition of 3M NaOH. The absorbance of reaction was measured at 405 nm. Cell viability was determined using the CellTiter-Blue cell viability assay. ALP activity was represented after normalization to the value of cell viability and each sample was measured in 6 replicates.
ALP staining
Cells were induced to osteogenic lineage and fixed with acetone/citrate buffer pH 4.2 (1.5:1) for 5 min at room temperature. Cells were stained with Napthol-AS-TR-phosphate solution (Sigma-Aldrich ApS) for 1 h at room temperature. Napthol-AS-TR-phosphate solution consists of Napthol-AS-TR-phosphate diluted 1:5 in H2O and Fast Red TR (Sigma-Aldrich ApS) diluted 1:1.2 in 0.1 M Tris buffer, pH 9.0, after which both solutions were mixed 1:1 and applied to the cells.
Alizarin Red S staining and quantification
Cells were induced to differentiate into osteoblasts for 10–12 days and fixed with 70% ice-cold ethanol for 1 h at –20°C. Cells were stained with Alizarin Red (40 mM, pH = 4; Sigma-Aldrich ApS) for 10 min at room temperature. For quantification of calcium deposition, Alizarin Red S was eluted with 10% cetylpyridinium chloride (Sigma-Aldrich ApS) for 1 h at room temperature. The absorbance of the eluted dye was measured at 570 nm.
Oil Red O staining and quantification
Cells were induced to differentiate into adipocyte using AIM, then fixed in 4% paraformaldehyde for 10 min at room temperature, and fate droplets were stained with Oil Red O (0.5 g in 60% isopropanol) (Sigma-Aldrich) for 1 h. For lipids quantification, Oil Red O was eluted from the cells with isopropanol for 10 min at room temperature and absorbance of the extracted dye was detected at OD 490 nm. Oil Red O values were normalized to cell number (measured by number of viable cells).
Western blot analysis
Cells were treated without or with carnosol and collected at specific time points and lysed in cell lysis buffer (10 mM Tris–HCl, pH 7.4, 150 mM sodium chloride, 1% NP-40, 0.1% SDS, 1 mM EDTA, 1 mM phenyl-methylsulfonyl fluoride, 1 mM NaF, 1 mM Na3VO4), supplemented with protease inhibitor cocktail (Roche Diagnostics, Mannheim, Germany). 30 μg of protein was separated on 8%–12% NuPAGE Novex Bis-Tris gel systems (Thermo Fisher Scientific, Roskilde, Germany) followed by transfer to PVDF membrane (Millipore, Bedford, MA, USA). The membrane was blocked and probed with antibodies (dil 1:1,000) and incubated with secondary anti-rabbit horseradish peroxidase-conjugated antibody (Santa Cruz Biotechnology, Aarhus, Denmark). Antibodies for Smad1/5/8 (total or phosphor) and β-Actin purchased from Santa Cruz Biotechnology, Inc. (Heidelberg, Germany). Quantification of Western blots was performed with ImageJ program.
RNA extraction and real-time PCR analysis
Total RNA was extracted from cells using TRIzol single-step method (Thermo Fisher Scientific). cDNA was synthesized from 1 μg of total RNA using revertAid H minus first strand cDNA synthesis kit (Fermentas, St Leon-Rot, Germany) according to the manufacturer’s instructions. Quantitative real time PCR was performed using Applied Biosystems 7500 Real-Time system with Fast SYBR Green Master Mix (Applied Biosystems, Foster City, CA, USA). Primer sequences for target genes were presented in Supplementary Table 1. The expression of each target gene was normalization to β-Actin and Hprt mRNA expression as reference genes, using a comparative CT method [(1/(2delta-CT) formula, where delta-CT is the difference between CT-target and CT-reference] with Microsoft Excel 2007 [23].
Statistical analysis
All values are expressed as mean ± standard deviation (SD), of at least 3 independent experiments. The data analyzed statistically for 2-samples using unpaired Student’s t-test (2-tailed) assuming equal variation in the two groups. Differences were considered statistically significant at *p < 0.05, and **p < 0.005.
RESULTS
Carnosol promotes osteoblast differentiation of mBMSCs
We first studied the cytotoxicity of carnosol on primary culture of mBMSCs using cell viability MTT assay and cell counting of viable cells. Carnosol showed no effect on cell viability up to concentration of 10 μM as assessed by MMT assay and cell count after 3 days in culture (Fig. 1A, B). On the other hand, carnosol displayed cytotoxicity at concentration of 20 μM. Therefore, we used a range of carnosol concentration from 1 to 10 μM for all of our subsequent experiments.
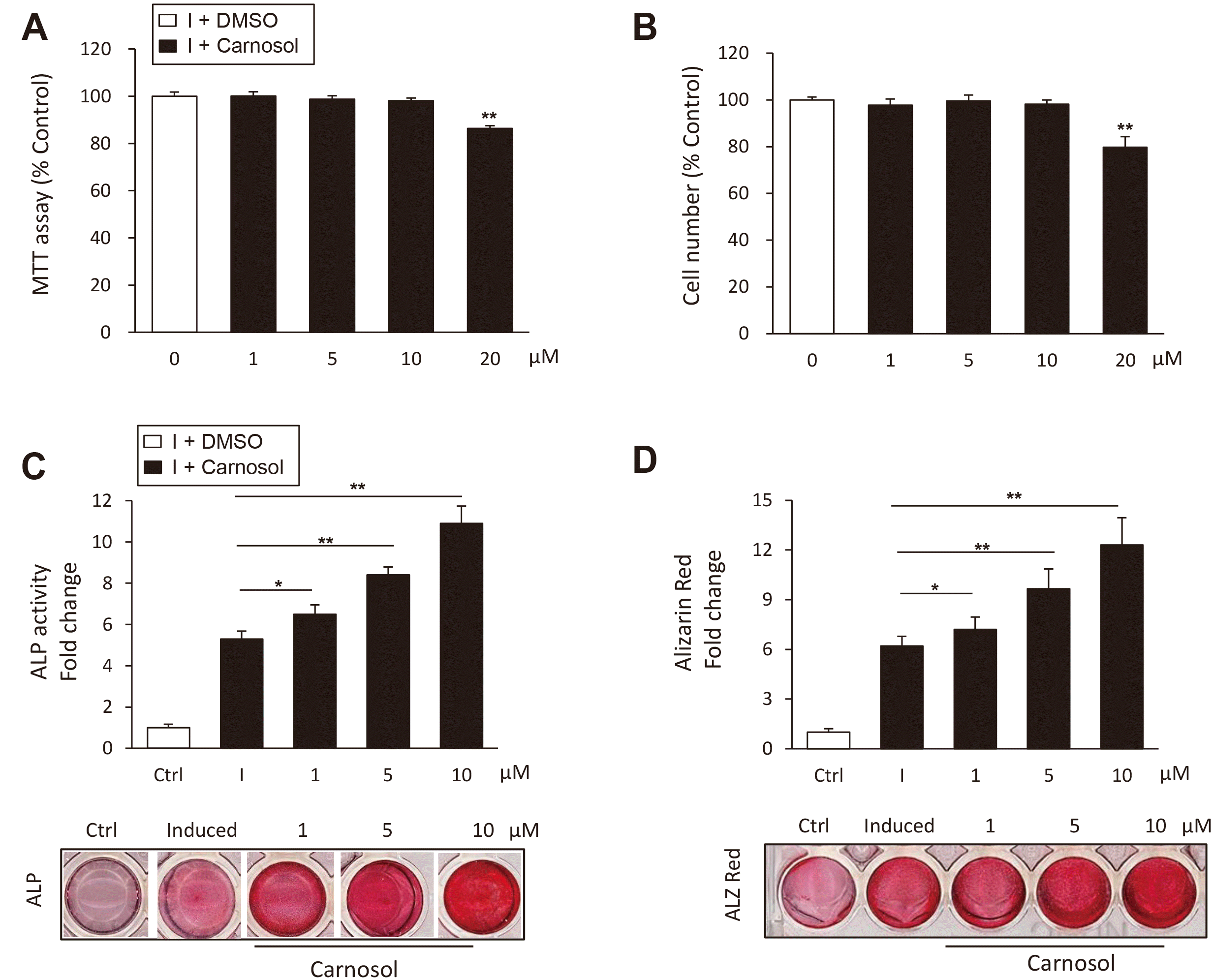
Fig. 1
Carnosol induces the differentiation of mouse bone marrow-derived mesenchymal stem cells (mBMSCs) into osteoblasts.
(A) Studying the cytotoxicity of carnosol on primary culture of mBMSCs using MTT cell viability assay. (B) Studying the effect of different concentrations of carnosol on mBMSCs cell proliferation as measured by counting cell number. Cells were either non-treated (0) or treated with different doses of carnosol for 3 days. (C) Stimulatory effect of carnosol on osteoblast differentiation of mBMSCs as assessed by ALP activity quantification and (D) Alizarin red staining quantification for matrix mineralization after 6 and 12 days of induction respectively. Stained images were shown under each graph. mBMSCs were either non-induced (control, ctrl), or induced with OIM in the absence (I) or the presence of different concentrations of carnosol. Values were shown as fold change over control. Values are mean ± standard deviation of three independent experiments (*p < 0.05, **p < 0.005 compared to Control [0] for [A and B] and compared to differentiated cells without carnosol [I] for panals [C and D]). ALP, alkaline phosphatase; OIM, Osteogenic-induction medium.
![]()
We examined the effect of carnosol on the osteoblast differentiation of mBMSCs. As shown in Fig. 1C, carnosol displayed dose-dependent stimulatory effect on ALP activity by mBMSCs during their differentiation into osteoblast lineage as compared to induced cells without carnosol (Fig. 1C). In addition, carnosol promoted matrix mineralization in mBMSCs during osteogenesis in dose-dependent manner as assessed by the quantification of Alizarin Red staining (Fig. 1D). We further studied the effect of carnosol on osteogenesis during the time course of 12 days of differentiation. Treatment of mBMSCs with carnosol (10 μM) showed to stimulate the ALP activity of mBMSCs during the whole differentiation course (Supplementary Fig. 1A).
Carnosol upregulates the expression of osteoblastic-related genes in mBMSCs
We studied further the effect of carnosol on osteogenesis at molecular level, by measuring its effect on osteogenic gene expression using qPCR. Treatment of mBMSCs during osteogenesis with carnosol showed to significantly upregulate the expression of both early (Runx2, Osx, Col1a1, and Alp) and late (Ocn and Opn) osteogenic markers (Fig. 2A). In addition, carnosol significantly upregulated the gene expression of Runx2, Alp and Ocn during the time course of osteoblast differentiation of mBMSCs (Supplementary Fig. 1B). QPCR-based osteogenic gene array analysis of carnosol-induced osteogenesis in mBMSCs revealed significant up-regulation of the expression of genes related to osteoblast differentiation and ossification and matrix formation by 31.8% and 27.3% respectively (Fig. 2B, Table 1).
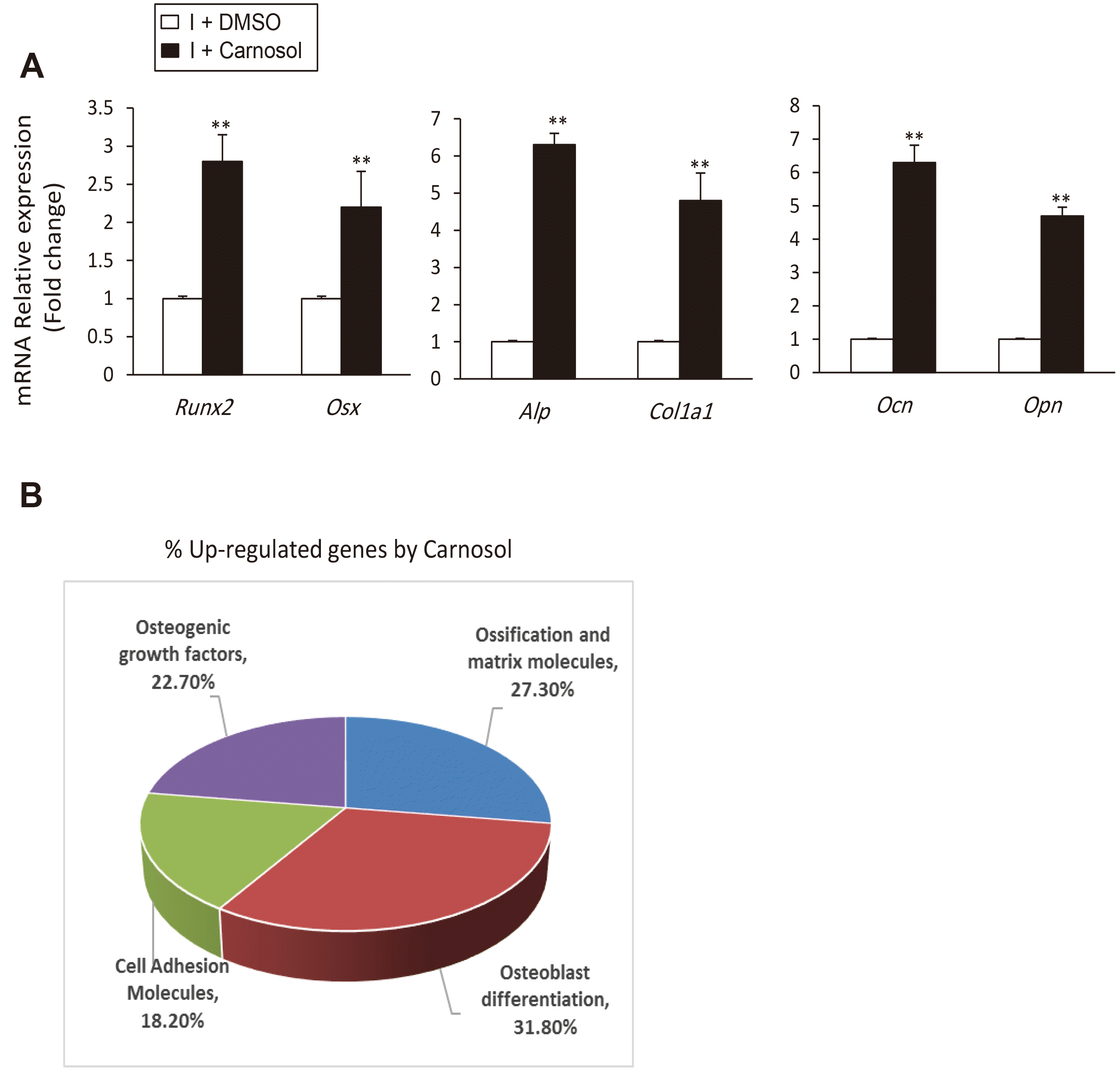
Fig. 2
Carnosol induces the expression of osteoblast-related genes.
(A) Carnosol upregulated the gene expression of early (Runx2, Osx, Alp, and Col1a1) and late osteogenic factors (Ocn and Opn) in mouse bone marrow-derived mesenchymal stem cells (mBMSCs) during osteogenesis. mBMSCs were induced to osteoblast differentiation using OIM in the absence (I+DMSO) or the presence of 10 µM carnosol (I+Carnosol) for 12 days. Gene expression values were normalization to reference genes and represented as fold change over induced cells without carnosol. (B) Analysis of carnosol-induced osteoblast-related genes in mBMSCs during osteogenesis using qPCR-based osteogenic gene array assay. The pie chart showed the percentage of upregulated genes as categorized by their osteogenic functions according to the data in Table 1. Values are mean ± standard deviation of three independent experiments (**p < 0.005 compared to I+DMSO). OIM, Osteogenic-induction medium.
![]()
Table 1
Upregulated osteogenic genes by carnosol during osteogenic differentiation of mBMSCs
Cells were induced to differentiate into osteoblast in the absence (control) or the presence of carnosol (10 µM). After 6 days of induction, total RNA were extracted and Mouse osteogenesis RT2 Profiler PCR array was performed was performed using quantitative PCR method. Up-regulated genes by carnosol were normalized to reference genes and represented as fold change over control cells without carnosol. Values are mean of 3 independent experiments. mBMSCs, mouse bone marrow-derived mesenchymal stem cells.
![]()
Carnosol inhibits the differentiation of mBMSCs into adipocytes
To examine, whether carnosol has the potential to direct the differentiation of mBMSCs into osteoblast versus adipocyte, we study the effect of carnosol on adipogenesis of mBMSCs. Carnosol showed to exert dose-dependent inhibitory effect on the differentiation of mBMSCs into adipocyte as assessed by for lipid accumulation and its quantification (Fig. 3A, B). In addition, Carnosol inhibited the capacity of mBMSCs to differentiate into adipocyte at different time points during the whole course of adipogenesis as assessed by quantitative Oil Red O staining (Supplementary Fig. 2A). At the molecular level, carnosol significantly down-regulated the mRNA expression of both early (Pparγ2 and C/ebpα) and late (aP2 and Lpl) adipogenic genes in mBMSCs as assessed by qPCR analysis (Fig. 3C). Furthermore, carnosol down-regulated both Pparγ2 and aP2 at different time points during the course of adipogenesis in mBMSCs (Supplementary Fig. 2B).
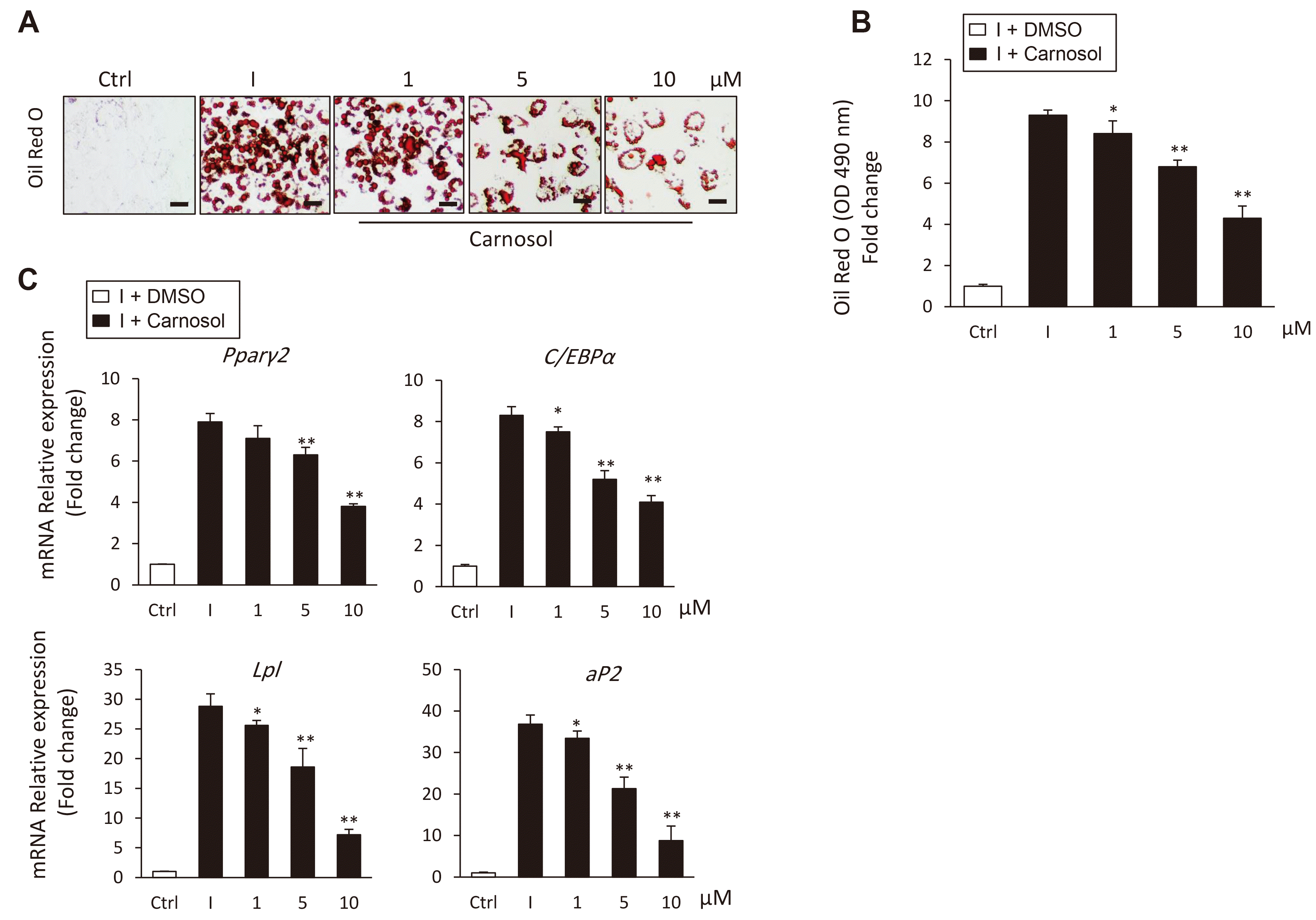
Fig. 3
Carnosol inhibits the differentiation of mouse bone marrow-derived mesenchymal stem cells (mBMSCs) into adipocytes.
(A) Inhibitory effect of carnosol on adipogenesis of mBMSCs as measured by Oil Red O staining for fat droplets formation. mBMSCs were induced to differentiated into adipocytes either in the presence of DMSO (I) as control or different concentrations of carnosol. Scale bars = 100 µm. (B) Quantification of Oil Red O staining after 12 days of differentiation. (C) Quantitative real-time PCR analysis of the adipogenic specific genes expression in the differentiated mBMSCs into adipocytes in the presence of different concentrations of carnosol. Gene expression values were normalized to reference genes and represented as fold inhibition to induced cells without carnosol. Values are mean ± standard deviation of three independent experiments (*p < 0.05, **p < 0.005, compared to [I]).
![]()
Carnosol stimulates BMP2-induced osteogenesis in mBMSCs
To understand the mechanism underlying the stimulatory effect of carnosol on osteogenesis, we studied the effect of carnosol on signaling molecules known to induce osteogenesis and bone formation (including TGFβ1, BMP2, IGF-1, PDGF, and bFGF). Interestingly, carnosol significantly stimulated BMP2-induced osteogenesis compared to other factors, as assessed by ALP activity measurements (Fig. 4A). This finding was also confirmed for other BMP members (BMP4 and 7) as assessed by significant increased matrix mineralization in treated mBMSCs with BMPs in the presence on carnosol versus cells treated without carnosol (Fig. 4B). In addition, treatment of mBMSCs with carnosol showed to significantly upregulate the BMP2-induced osteoblastic target genes (Alp, Ocn, Col1a1, Runx2, Dlx5, and Msx2) as compared to non-treated cells (Fig. 4C).
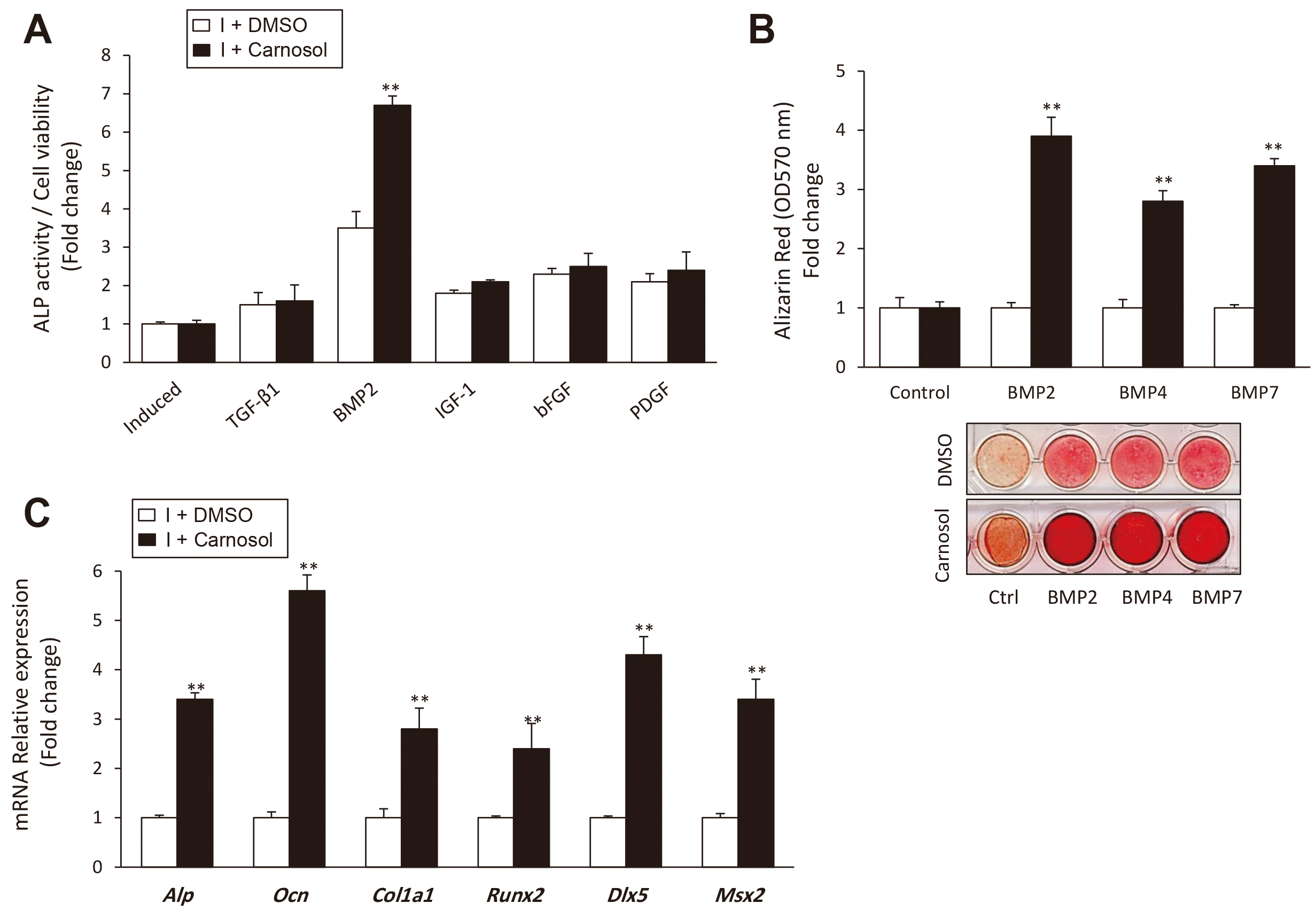
Fig. 4
Carnosol promotes bone morphogenetic protein (BMP)-induced osteogenesis in mouse bone marrow-derived mesenchymal stem cells (mBMSCs).
(A) Studying the effect of carnosol on promoting the osteogenic induction of different osteogenic growth factors in mBMSCs. Cells were induced for osteogenesis without (I+DMSO) or with TGFβ1 (10 ng/ml), BMP2 (100 ng/ml), IGF1 (100 ng/ml), bFGF (100 ng/ml) or PDGF (100 ng/ml) for 6 days in the absence or the presence of 10 µM carnosol for 6 days. (B) Effect of carnosol on BMP2, 4, and 7-induced osteogenesis in mBMSCs as measured by quantitative Alizarin Red staining for matrix mineralization after 12 days of induction. Representative images of Alizarin Red staining were shown. (C) Effect of carnosol on inducing the expression of BMP2-upregulated osteogenic gene expression. Gene expression values were normalization to reference genes and represented as fold change over induced cells without carnosol. Values are mean ± standard deviation of three independent experiments (**p < 0.005, compared to induced cells without carnosol [I+DMSO]).
![]()
The mechanism of carnosol-induced osteogenesis is mediated by BMP signaling pathway
To investigate, whether the stimulatory effect of carnosol on osteogenesis is mediated via BMP signaling, we first studied the possible activation of BMP2 signaling in mBMSCs by carnosol. Carnosol displayed does-dependent activation of Smad 1/5/8 phosphorylation as assessed by Western blot analysis (Fig. 5A). Furthermore, inhibition of BMP signaling in mBMSCs with a specific BMP1R inhibitor, LDN-193189, showed to significantly inhibit the stimulatory effect of carnosol on BMP-induced Smad 1/5/8 phosphorylation as assessed by western blot analysis (Fig. 5B). Consequently, treatment of mBMSCs with LDN-193189 in the presence of carnosol, showed to attenuate the stimulatory effect of carnosol on osteogenesis by 62% compared to treated cells without inhibitor as measured by the quantitative ALP activity assay (Fig. 5C).
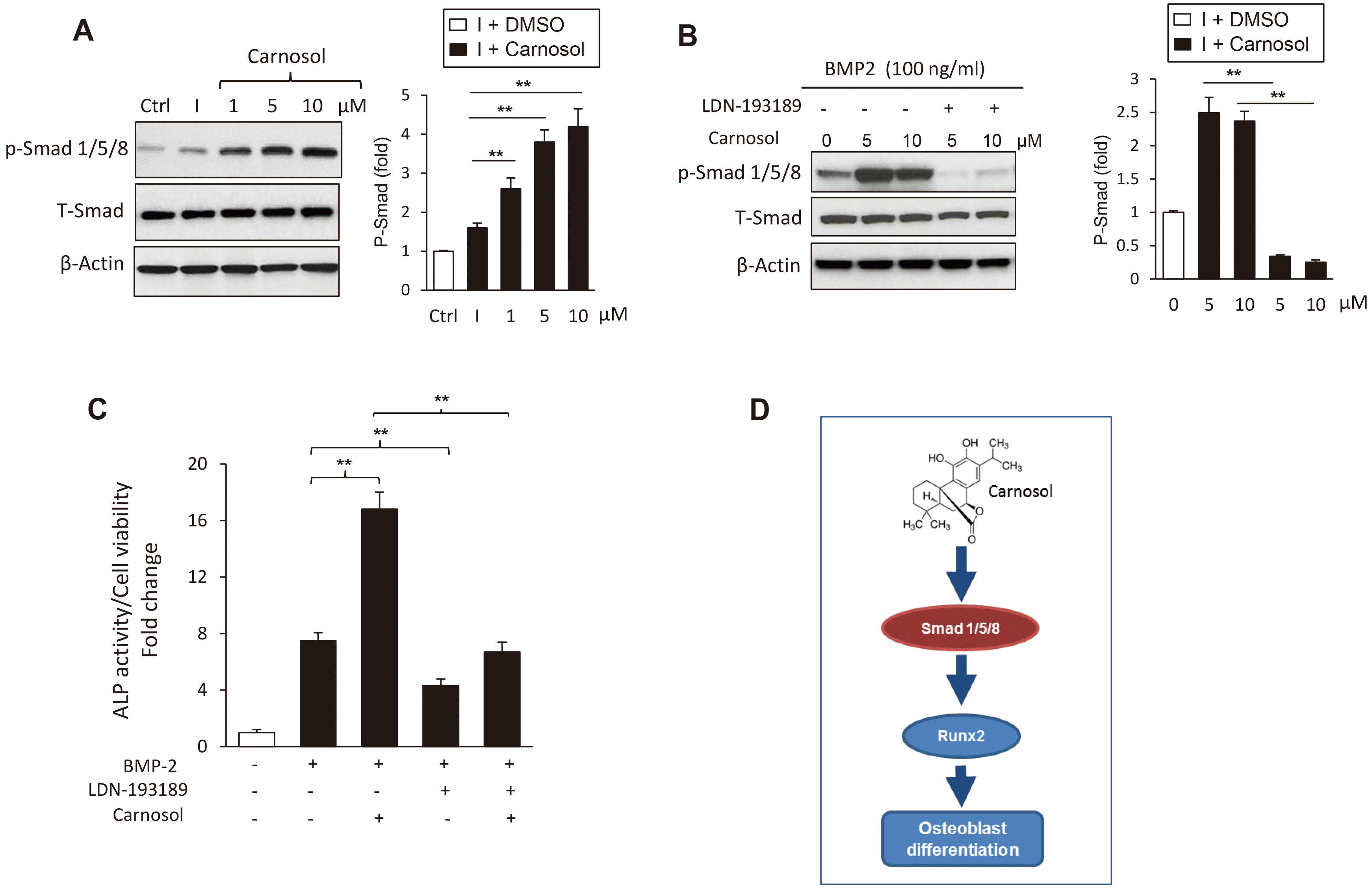
Fig. 5
Carnosol promotes osteoblast differentiation of mouse bone marrow-derived mesenchymal stem cells (mBMSCs) through the activation of bone morphogenetic protein (BMP) signaling pathway.
(A) Western blot analysis showing the dose-dependent stimulatory effect of carnosol (10 µM) on the activation of Smad1/5/8 phosphorylation in mBMSCs after 10 min. (B) Western blot analysis demonstrating the ability of BMP1R inhibitor (LDN-193189, 10 µM) to inhibit the stimulatory effect of carnosol on BMP-induced Smad1/5/8 phosphorylation in mBMSCs after 10 min of treatment. (C) BMP1R inhibitor (LDN-193189, 10 µM) suppressed the stimulatory effect of carnosol (10 µM) on BMP2-induced osteogenesis in mBMSCs as measured by quantitative ALP activity. Cells were pre-treated with LDN-193189, 10 µM and induced with BMP2 in the absence or the presence of carnosol (10 µM) for 6 days. (D) Schematic diagram illustrating the mechanism of the stimulatory effect of carnosol to on osteogenesis via activating canonical BMP signalling. Values are mean ± standard deviation of three independent experiments (**p < 0.005). ALP, alkaline phosphatase.
![]()
DISCUSSION
Carnosic acid and its major oxidative derivatives, carnosol are major antioxidant components (> 90%) of rosemary (Rosmarinus officinalis) that showed many applications in field of nutrition, and health [24,25].
In this study, we identified the medicinally important phytochemical, carnosol as a novel osteogenic factor that promotes the differentiation of BMSCs into osteoblasts, while inhibits their differentiation into adipocytes. Our data revealed that the stimulatory effect of carnosol on osteogenesis is mediated via BMP-signaling pathway.
Our data demonstrated that carnosol promoted osteogenic differentiation at the early and late stages by increasing ALP activity and matrix mineralization respectively. In consistent, at the molecular level, carnosol upregulated the expression of early and late osteogenic markers, suggesting the involvement of carnosol in directing the early commitment of BMSCs into osteoblastic cell lineage and promoting mature osteoblast formation.
BMP signaling is an essential pathway that involved in osteoblast differentiation and bone formation. Binding of the osteogenic inducers, BMPs to its serine–threonine receptors (BMP type I receptor and type II receptors) leads to activate the phosphorylation of Smad1/5/8 (phosphorylated R-Smad). Subsequently, phosphorylated R-Smad binds to SMAD4 and co-translocates into the nucleus to activate the expression of main osteogenic transcription factor Runx2 and osteogenic target genes, i.e., Ocn, Dlx5, and Msx2 [26-28]. Our data, identified carnosol as a novel inducer for BMP-induced osteogenesis, via canonical-Smad-dependent pathway in a dose-dependent manner. In addition, the stimulatory effect of carnosol on osteogenesis in mBMSCs was abolished upon the inhibition of BMP signaling by specific BMP1R inhibitor. In this context, the biological effects of carnosol were found to be mediated by several signaling molecules. Carnosol was reported to inhibit cartilage breakdown in osteoarthritic chondrocytes via the inhibition of pro‐inflammatory factors [18] and to suppress osteoclast formation and activity by inhibiting NF‐κB ligand (RANKL)‐induced osteoclast [21]. Carnosol exerts long term antioxidant effect by activating phosphatidylinositol 3-kinase (PI3K)/Akt pathway and HO‐1 pathways [29]. Furthermore, many signaling molecules are implicated in the anticancer properties of carnosol including molecules associated with cell cycles including PI3K/Akt, mammalian target of rapamycin (mTOR), and 5’-AMP-activated protein kinase (AMPK) and androgen and estrogen receptors [25,30,31] .
Our data demonstrated for the first time that carnosol promotes osteogenesis by activating BMP-signaling and upregulating its downstream osteogenic target genes.
However, the mechanism of how carnosol can activate BMP signaling is still unknown. Further studies are required to investigate, whether carnosol can bind to BMP receptors or acting downstream intracellularly to activate Smad4 protein. In regards to the affinity of carnosol to some cell receptors, carnosol showed to inhibit both prostate and breast cancers by binding and antagonizing estrogen and androgen receptors [32-34].
Our data demonstrated the inhibitory effect of carnosol on adipogenesis by suppressing both early and late stages of adipocyte differentiation of BMSCs. In consistent, carnosol inhibits the differentiation of mouse pre-adipocyte 3T3-L1 cells into mature adipocyte by stimulating glutathione metabolism [35]. The antiadipogenic effect of carnosol was revealed by its capacity to protect lipids from oxidation in vitro [24], and to reduce lipid absorption in mice by inhibiting pancreatic lipase(s) [36]. In addition, administration of carnosol to diabetic rats showed to excrete antidiabetic effects by reducing blood glucose, triglyceride and total cholesterol in association with reduced plasma levels of oxidative stress [37].
The lineage specific differentiation of BMSCs into osteoblasts or adipocytes is controlled by several signaling molecules including Wnt, Notch, BMPs and TGF-β1 [3,8,38,39]. In this context, we recently reported that the adipocytes in bone marrow microenvironment exerts paracrine inhibitory effects on osteogenesis via suppressing BMP-induced osteogenesis [40]. Thus, the inhibitory effect of carnosol on adipogenesis of BMSCs is beneficiary for the stimulatory effect of carnosol on BMP-signaling activation and subsequently promoting osteoblast differentiation.
Phytochemicals are renewable source of compounds with therapeutic potential in many metabolic diseases. In this study, we identified carnosol as a novel osteogenic phytochemical with a high potential to direct the differentiation of BMSCs into osteoblastic cell lineage versus adipocytic cell lineage. The stimulatory effect of carnosol on osteogenesis showed to be mediated via BMP signaling-dependent mechanism (Fig. 5D). Thus, our data provide carnosol as a novel renewable source of phytochemical that plausibly can be used directly as an osteoanabolic drug or as raw material to develop drug for enhancing bone formation in many bone-loss related diseases.
SUPPLEMENTARY MATERIALS
Supplementary data including one table and two figures can be found with this article online at https://doi.org/10.4196/kjpp.2021.25.3.197.
 XML Download
XML Download