

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Neuropathic pain (NP) is a chronic pain caused by central or peripheral nerve damage, long-term diabetes, or chemotherapy drugs [1]. NP is often accompanied by decreased cognitive function, anxiety, depression, and insomnia; these symptoms may lead to a decline in the quality of life [2]. The comorbidity between pain and depression is considered the bottleneck of NP clinical treatment [3]. Accumulating evidence indicates that neuroinflammation is related to the pathophysiology of depression [4]. A study reported that proinflammatory cytokines were elevated in the hippocampus of patients with major depression [5]. Some studies showed that a variety of proinflammatory cytokines activated by hippocampal microglia are involved in the occurrence and maintenance of both anxiety and depression-like behaviors [6,7].
The toll-like receptor 4 (TLR4)/nuclear factor-kappa B (NF-κB) is a Ca2+-independent signaling pathway that activates microglia and enhances pain signal transduction. When peripheral nerves are injured, nociceptive transmitters combine with TLR4 to activate it, thereby activating microglia, which in turn activate NF-κB and downstream factors, promoting the synthesis and release of inflammatory mediators, such as tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6), and thus, inducing the production and enhancement of pain [8]. Cheng et al. [9] found that TLR4 and NF-κB were highly expressed in the brain tissue of mouse depression model. They found intense brain tissue inflammation and the severity of the inflammation was related to the degree of depression-like symptoms in mice. This suggests that the inflammatory cascade caused by the activation of the TLR4/NF-κB pathway may be associated with the induction of depression.
Paeoniflorin (PF) is the main bioactive component of Paeonia lactiflora pall [10]. PF can ameliorate central nervous system diseases, such as cognitive impairment, Parkinson’s disease, and depression [11-13]. It also possesses sedative, anti-inflammatory, and analgesic effects [14,15]. Studies have found that PF can inhibit the activation of spinal cord microglia and exert anti-inflammatory effects [16]. However, in the NP model, it has not been reported whether PF improves pain-related depression by regulating the TLR4/NF-κB signaling pathway in the hippocampus. Therefore, this study aimed to investigate the effect of PF on depression-like behaviors induced by NP, and its relationship with the TLR4/NF-κB signaling pathway in the hippocampus. This will further provide a new experimental basis for PF to treat the comorbidity between NP and depression.
Go to : 
METHODS
Drugs and reagents
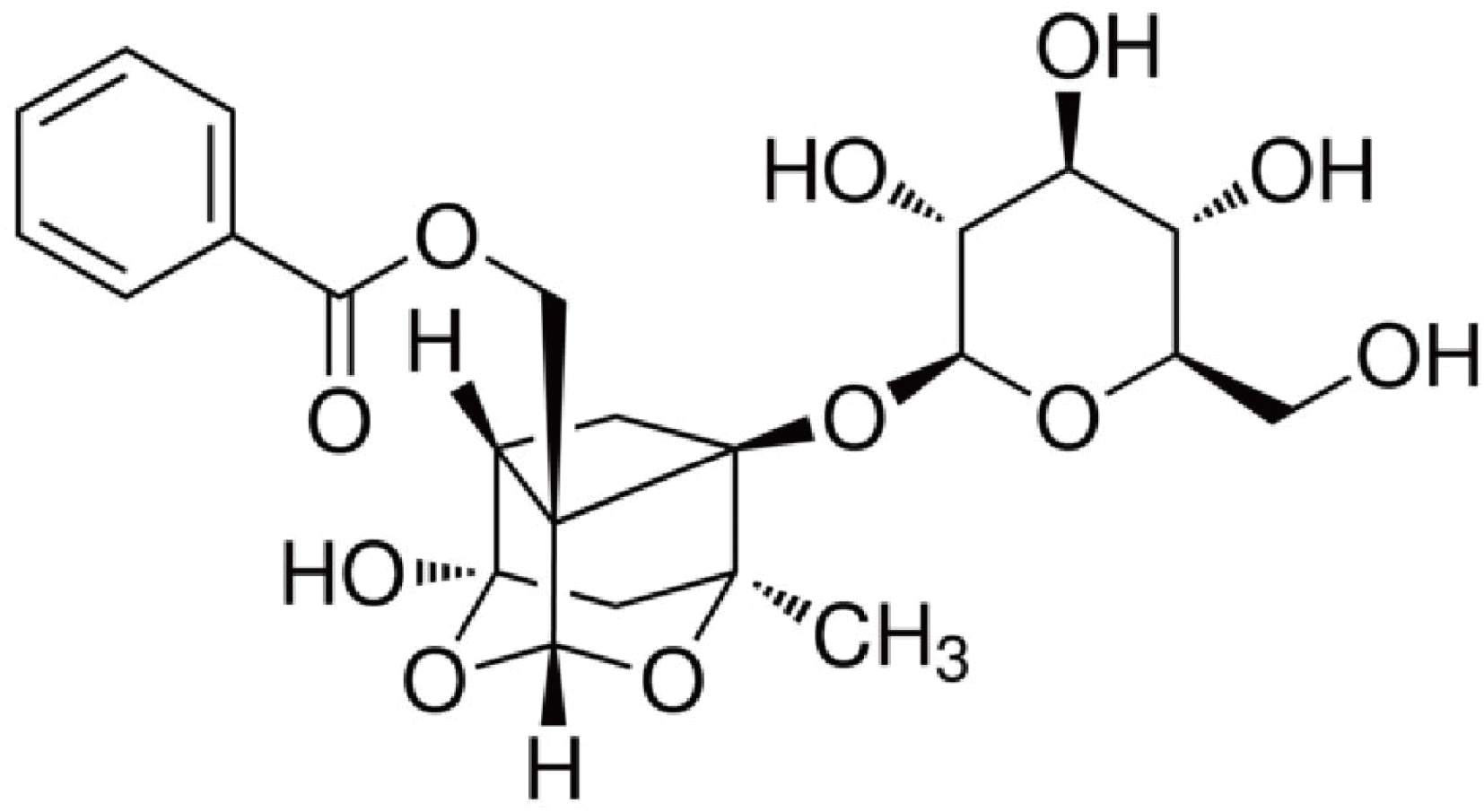
PF (purity ≥ 98%) was acquired from Sigma-Aldrich (Product Number: P0038 CAS Number: 23180-57-6, Saint Louis, MO, USA), the chemical structure of PF is shown in Fig. 1, it is a white powder, has a formula: C23H28O11 and formula weight: 480.46 g/mol, dissolved in methanol for use, the concentration is 1 mg/ml. TNF-α, IL-6, and IL-1β enzyme-linked immunosorbent assay (ELISA) kits were produced by Nanjing SenBeiJia Biological Technology Co. Ltd. (Nanjing, China). All antibodies were provided by Cell Signaling Technology (Danvers, MA, USA).
Animals
Specific pathogen free grade male Balb/c mice (n = 40), weighing 18–22 g, were supplied by the Animal Science Experimental Center of Yanbian University (certificate number: SCXK(J)2011-0007). Animals were maintained in room temperature (22°C ± 2°C) and humidity, with food and water available ad libitum. This study was approved by the Laboratory Animal Ethics Committee of Yanbian University (study approval number: 20190712, Yanji, China).
Preparation of model and treatment
Generation of sciatic nerve cuffing (Cuff) mouse models: The mice were anesthetized by intraperitoneal injection of 5% chloral hydrate (0.1 ml/10 g). The method of exposing the sciatic nerve trunk was carried out according to Benbouzid et al. report [17]. The nerve trunk was carefully separated by a glass minute needle, and a 2 mm polyethylene tube (inside diameter = 0.38 mm, outside diameter = 1.09 mm; PE-20) with one opening inserted into the sciatic nerve trunk. The wound was sutured after treatment with penicillin. The mice in the Sham group received the same surgical procedure as above, but only the main sciatic nerve was isolated, and the cuff was not implanted.
The mice were randomly classified into four groups (n = 10/group): Sham surgery group (Sham), Cuff group (Cuff), Cuff + 50 mg/kg PF group (PF 50 mg/kg), and Cuff + 100 mg/kg PF group (PF 100 mg/kg). The dosages of PF were selected based on our preliminary data. Mice in the PF group were injected with 50 and 100 mg/kg PF intraperitoneally once a day for 14 consecutive days starting on the first day after surgery (the time of administration was 10–12 am). The Sham surgery and Cuff groups were injected with the same volume of normal saline.
Mechanical allodynia test and thermal hyperalgesia test
The method of measuring the paw withdrawal threshold (PWT) of the hind limbs was done according to the method published by Chaplan et al. [18]. After administration, the mice were placed in a plexiglass box in a quiet room to adapt for 20 min and to maintain immobility. We used von Frey filaments with different stimulation intensities to stimulate the soles of the mice. The stimulation intensity generally starts from 0.4 g. If a mouse did not lift its foot when the von Frey filaments were bent by more than 90°, it was considered unresponsive. At this time, the adjacent von Frey filaments with a higher stimulation intensity should be used; if there is a response, the adjacent von Frey filaments with a lower stimulation intensity should be used. The maximum stimulation intensity in this experiment was 4.0 g. We recorded the stimulus intensity value of the tested mice that caused the foot-lifting response 50% of the time, which was recorded as the PWT (g).
According to Wang et al.’s method [19], a pathway-cheps thermal pain stimulator (Beijing Hudoba Biotechnology Co., Ltd., Beijing, China) was used to assess thermal hyperalgesia. The mice were individually placed in a plexiglass box and were allowed to adapt for 30 min. Radiant heat was then applied to the surface of the sole of the hind paw on the surgical side. The paw withdrawal latency (PWL), defined as the time taken for the mice to lick or withdraw their paws was taken and recorded. In order to avoid tissue damage, we set an automatic delay cutoff time of 30 s, with an interval of 10 min between each measurement.
Depression-related behavior test
Sucrose preference test (SPT): The mice were trained to adapt drinking 1% sucrose solution 2 days before the test. The sucrose solution was then replaced with distilled water and left it for 24 h. After fasting for 23 h, the mice were each given one bottle of the pre-weighed 1% sucrose solution and distilled water. After 1 h, the consumption of sugar and pure water was measured. The sugar water preference (SP) was calculated according to the following formula: SP = sugar water intake (g)/[sugar water intake (g) + pure water intake (g)] × 100%.
Forced swimming test (FST): The mice were placed in a transparent bucket with a height of 25 cm and a diameter of 15 cm, with a water depth of 15 cm, so that no part of the mice limbs could touch the bottom of the tank. The water temperature was maintained at 24°C for 6 min, and the immobility time of the mice was recorded in the last 4 min of the test.
Tail suspension test (TST): The mice were suspended 15 cm from the ground with a tape fixed about 1 cm from the tail tip for 6 min. The time of immobility (s) was recorded in the last 4 min of the test.
Hematoxylin & eosin (H&E) staining
The brains of the animals in each group were fixed in 4% paraformaldehyde, dehydrated with 70%, 85%, 90%, 95%, and 100% ethanol. The brains were then embedded in paraffin and sliced at a thickness of 5 μm. H&E staining kit was used and H&E were used according to the instructions; the stained sections were dehydrated and sealed for preservation.
Immunological staining
Sections were dried, deparaffinized, and hydrated. The antigen unmasking solution was heated at high temperature and the sections were incubated for 20 min. The solution was then allowed to stand to return to room temperature. Endogenous peroxidase blocker was added and incubated at room temperature for 10 min. The sections were incubated with Iba-1 (0.5% Blocking buffer, 1:500 dilution) primary antibody at 4°C overnight, reheated for 30 min, and then incubated with goat anti-rabbit secondary antibody (0.5% BSAT, 1:400 dilution) at room temperature for 30 min. They were washed with 0.5% PBST (Phosphate Buffered Saline with 0.1% Tween-20). Freshly prepared DAB (3,3’-diaminobenzidine) color developing solution was used for 2–10 min to patch and cover film. The photomicrograph system (Leica Camera AG, Solms, Germany) system was used to collect images, and the image-Proplus software (Media Cybernetics Image Technology Company, Orlando, FL, USA) was used to draw the cell body area and analyze the statistics.
ELISA
The hippocampal tissue was weighed to make a 10% tissue homogenate. The supernatant was collected and stored in a freezer at –80°C for later use. The assay was performed according to the instructions of the ELISA kit, and the optical density value of the final reaction product at 450 nm in a microplate reader was plotted using GraphPad Prism 8.4.2 (GraphPad Software, San Diego, CA, USA). The concentrations of TNF-α, IL-6, and IL-1β were calculated based on the absorbance readings of TNF-α, IL-6, and IL-1β standard curve.
Western blot analysis
The total protein extraction kit (Millipore, Billerica, MA, USA) was used to extract hippocampal tissue proteins and to determine protein concentration. The proteins were separated on SDS-PAGE gel (50 μg/lane) and were transferred to a polyvinylidene difluoride membrane. After blocking with 5% skim milk for 1 h, the TLR4, MyD88, IκBα, p-NF-κBp65, and NF-κBp65 antibodies (dilution 1:1,000) were incubated overnight at 4°C. The membranes were washed and probed with corresponding horseradish peroxidase-conjugated secondary antibody, and finally, were imaged with the Electro-Chemi-Luminescence kit (Kaiji Bio Co., Nanjing, China). ImageJ software (http://rsbweb.nih.gov) was used for semi-quantitative analysis of western blot bands.
Statistical analysis
ImageJ software was used to process the pictures collected above as required. GraphPad Prism 8 software was used to perform the statistical analysis on the data. All data were expressed as mean ± standard deviation. The comparison between groups was performed by one-way analysis of variance. A p-value of < 0.05 means that the difference is statistically significant.
Go to :
RESULTS
Effect of PF on Cuff-induced hyperalgesia
The mechanical pain threshold and thermal withdrawal latency of all the groups were detected 1 day before model creation and at days 2, 5, 7, 10, and 14 after model creation. The results are shown in Fig. 2. The PWT and PWL in the Cuff group were significantly lower than those in the Sham group (PWT, p < 0.001; PWL, p < 0.001). After PF treatment, the PWT and PWL of mice were significantly increased (PWT, p < 0.001; PWL, p < 0.001). In addition, it was positively correlated with increasing concentration of PF. The results suggest that the Cuff model successfully induced NP in mice. The PF reduced the mechanical hyperalgesia and thermal hyperalgesia in a dose-dependent manner and has a strong analgesic effect on NP.
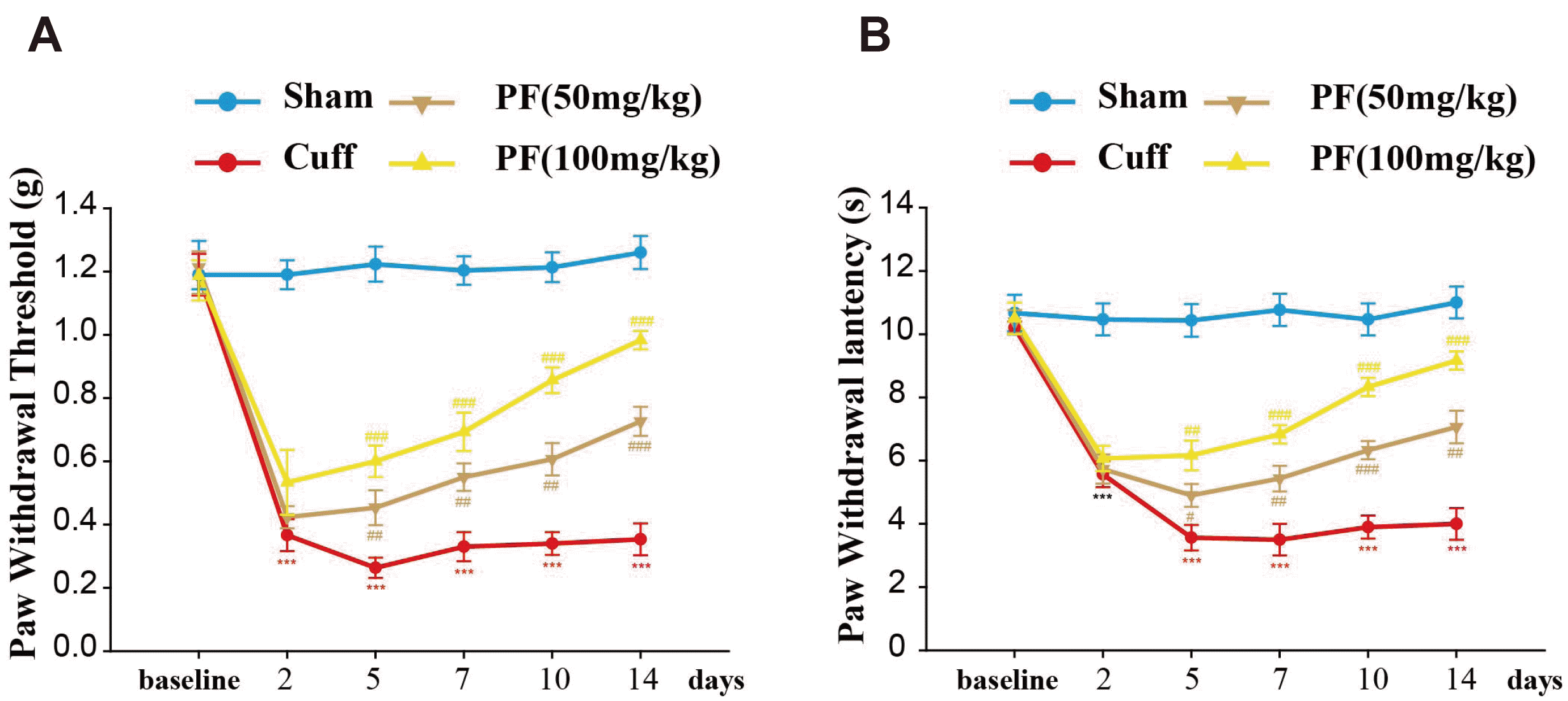 | Fig. 2PF treatment alleviates Cuff-induced hypersensitivity to pain.Effects of an intraperitoneal injection of PF (50, 100 mg/kg) on mechanical allodynia (A) and thermal hyperalgesia (B) in Cuff model mice. All values are expressed as mean ± standard deviation. n = 10. PF, paeoniflorin. ***p < 0.001 vs. Sham group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. Cuff group.
|
Effect of PF on depressive-like behavior of NP mice
The SPT is a test to check the response of the mice to reward stimuli, and a lack of interest in reward stimuli is an important manifestation of depression. As shown in Fig. 3A, there were significant differences seen in the rate of sucrose consumption in the four groups of mice in SPT. The mice in the Cuff group showed a statistically significant decrease (p < 0.01) in sugar preference 14 days after surgery, compared with the Sham group. The sugar preference of the PF 50 mg/kg and PF 100 mg/kg groups was significantly increased (p < 0.05, p < 0.01) compared with the Cuff group. Results suggested that PF could inhibit the anhedonia successfully induced by the Cuff model in mice, indicating its antidepressant effects.
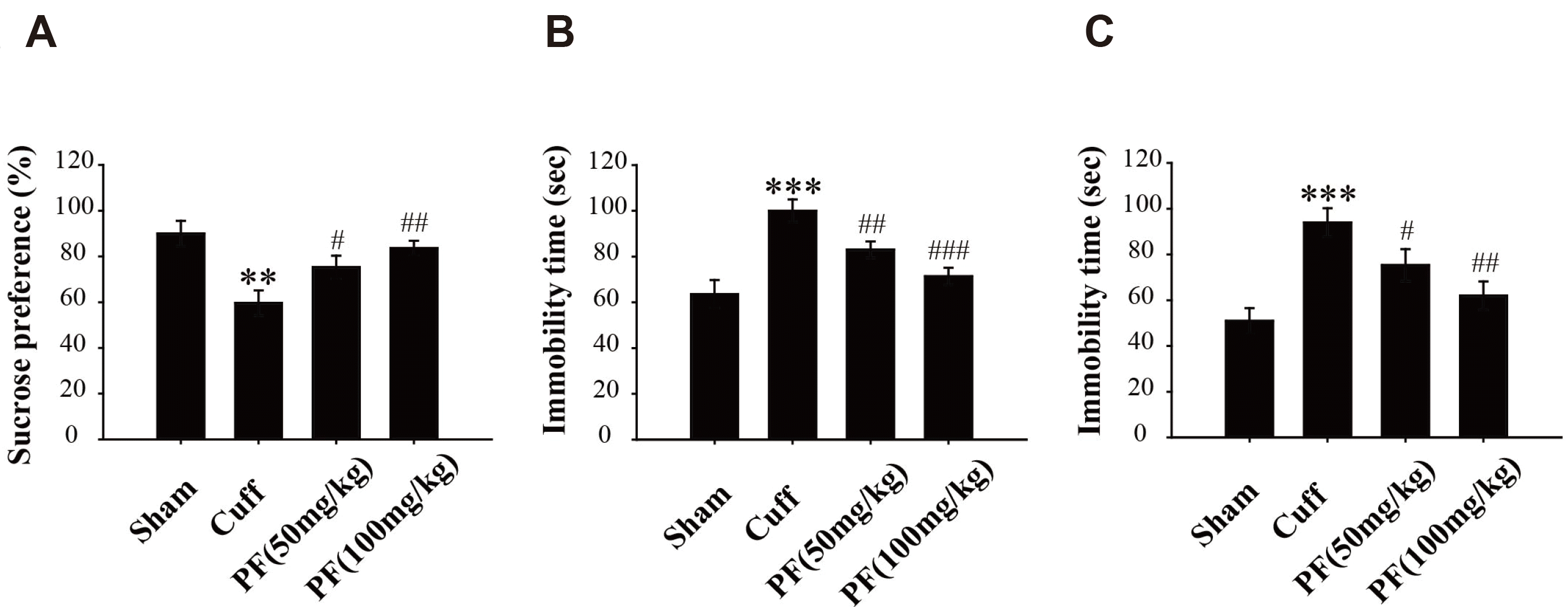 | Fig. 3Effects of PF on the depressive behavior of NP model mice.Effects of PF (50, 100 mg/kg) on sucrose preference test (A), forced swimming test (B), and tail suspension test (C) in Cuff model mice. Results are presented as mean ± standard deviation. n = 10. PF, paeoniflorin; NP, neuropathic pain. **p < 0.01, ***p < 0.001 vs. Sham group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. Cuff group.
|
The increased immobility time of mice in the FST and TST is considered to be a manifestation of hopelessness. As shown in Fig. 3B, C, there were significant differences in the immobility time in FST and TST between the four groups of mice. The immobility time of mice in the Cuff group was significantly increased compared with that in the Sham group (FST, p < 0.001; TST, p < 0.001). The treatment with PF shortened the time of Cuff mice (FST, p < 0.01, p < 0.001; TST, p < 0.05, p < 0.01), and the response was found to be dose dependent. The results suggest that the Cuff model successfully enhanced emotions of despair in mice, and PF can significantly improve despair-like emotions.
Effect of PF on morphological changes in hippocampal CA3 structure in NP mice
H&E staining results are displayed in Fig. 4A. The pyramidal cells in the CA3 region of the Sham group were arranged neatly and tightly, without vacuoles and obvious nucleoli. In the Cuff group, the pyramidal cells in the CA3 region were arranged loosely, with vacuolar changes and inflammatory cell infiltration. The number of necrotic and inflammatory cells were significantly reduced and the cell arrangement was neat in the PF (50 mg/kg) group. After 14 days of treatment with PF (100 mg/kg), the morphology of the pyramidal cells in the CA3 region normalized, and the arrangement was also basically normal, with no significant difference compared with the Sham group. These results suggest that PF can reduce pyramidal cell damage and inhibit the inflammatory response in the CA3 region of the hippocampus.
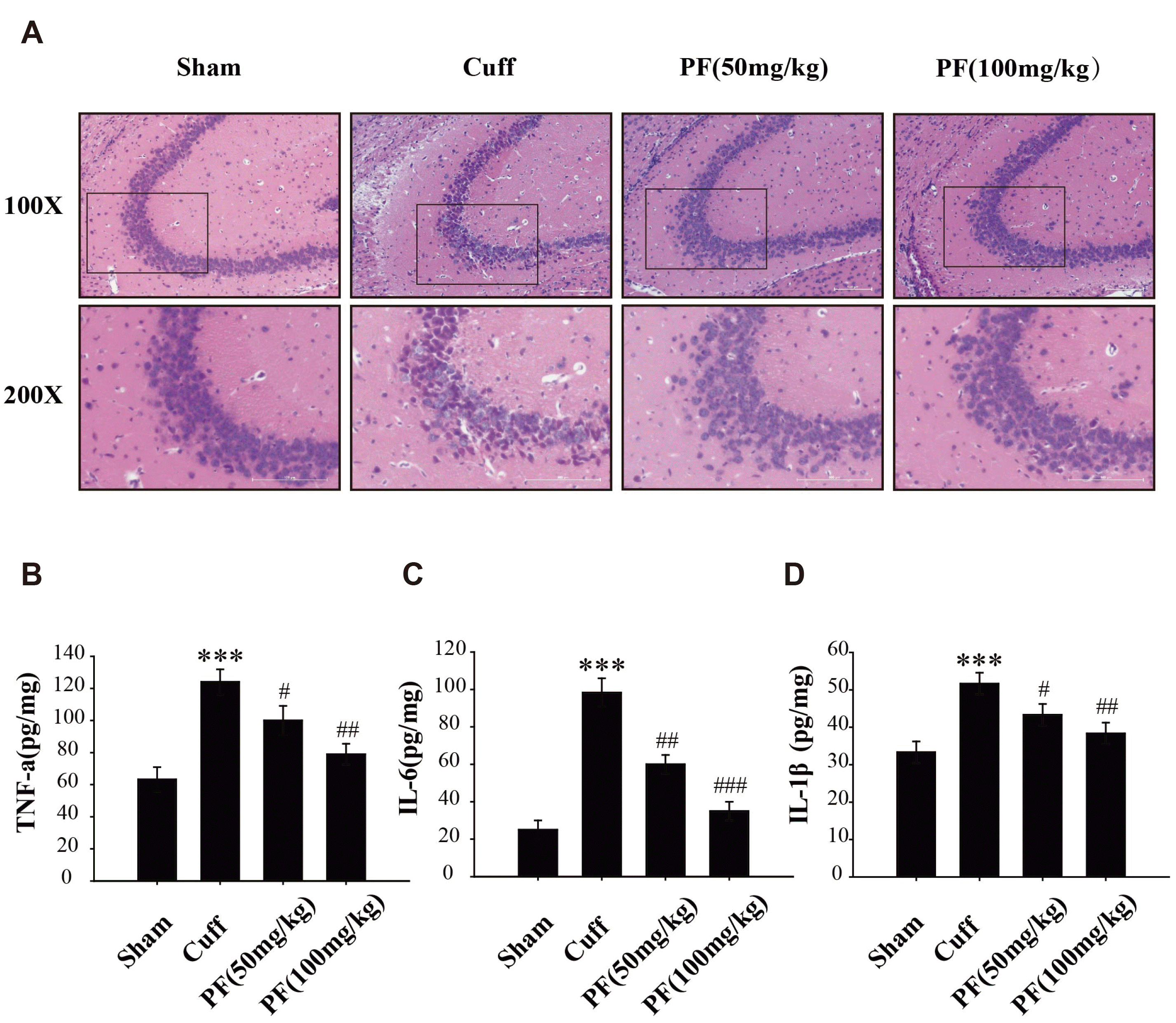 | Fig. 4PF improves the inflammatory infiltration of hippocampal pyramidal cells and reduces the expression of pro-inflammatory cytokines.Representative photomicrographs of H&E staining in the CA3 (Bar = 100 μm) region of hippocampus in different groups (A). ELISA showed that PF treatment decreased the expression of TNF-α (B), IL-6 (C), and IL-8 (D) caused by Cuff. The data represent the mean ± standard deviation. n = 4. PF, paeoniflorin. ***p < 0.001 vs. Sham group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. Cuff group.
|
Effect of PF on the expression of pro-inflammatory cytokines in the hippocampus of NP mice
We used ELISA to detect the levels of pro-inflammatory cytokines TNF-α, IL-6, and IL-1β in the hippocampus of mice. As shown in Fig. 4B–D, compared with the Sham group, the Cuff group caused a significant increase in the levels of TNF-α, IL-6, and IL-1β in the hippocampus (p < 0.001). After PF treatment, the levels of TNF-α, IL-1β, and IL-6 were significantly reduced (TNF-α, p < 0.05, p < 0.01; IL-6, p < 0.01, p < 0.001; IL-1β, p < 0.05, p < 0.01). These results suggest that the abnormal increase of pro-inflammatory cytokines in the hippocampus may be involved in the influence of NP-related depression, and PF can inhibit the expression of pro-inflammatory cytokines in the hippocampus.
Effect of PF on the activation of microglia induced by NP mice
We used Iba-1 as a microglia activation marker to observe whether PF played a role in the activation of hippocampal microglia in NP mice induced by the Cuff model. The Iba-1 positive cells were scattered in the hippocampal CA1 and DG regions of the Sham group (Fig. 5A). The cell bodies were elongated with dendritic protrusions, indicating a resting state. Compared with the Sham group, the Iba-1 positive cell bodies of the mice in the Cuff group became larger, the protrusions were short and thick, and the branch retraction was reduced, showing an activated state. After treatment with PF, the activated morphology of Iba-1 positive cells in mice was significantly improved compared with the Cuff group, leaning to a resting morphology.
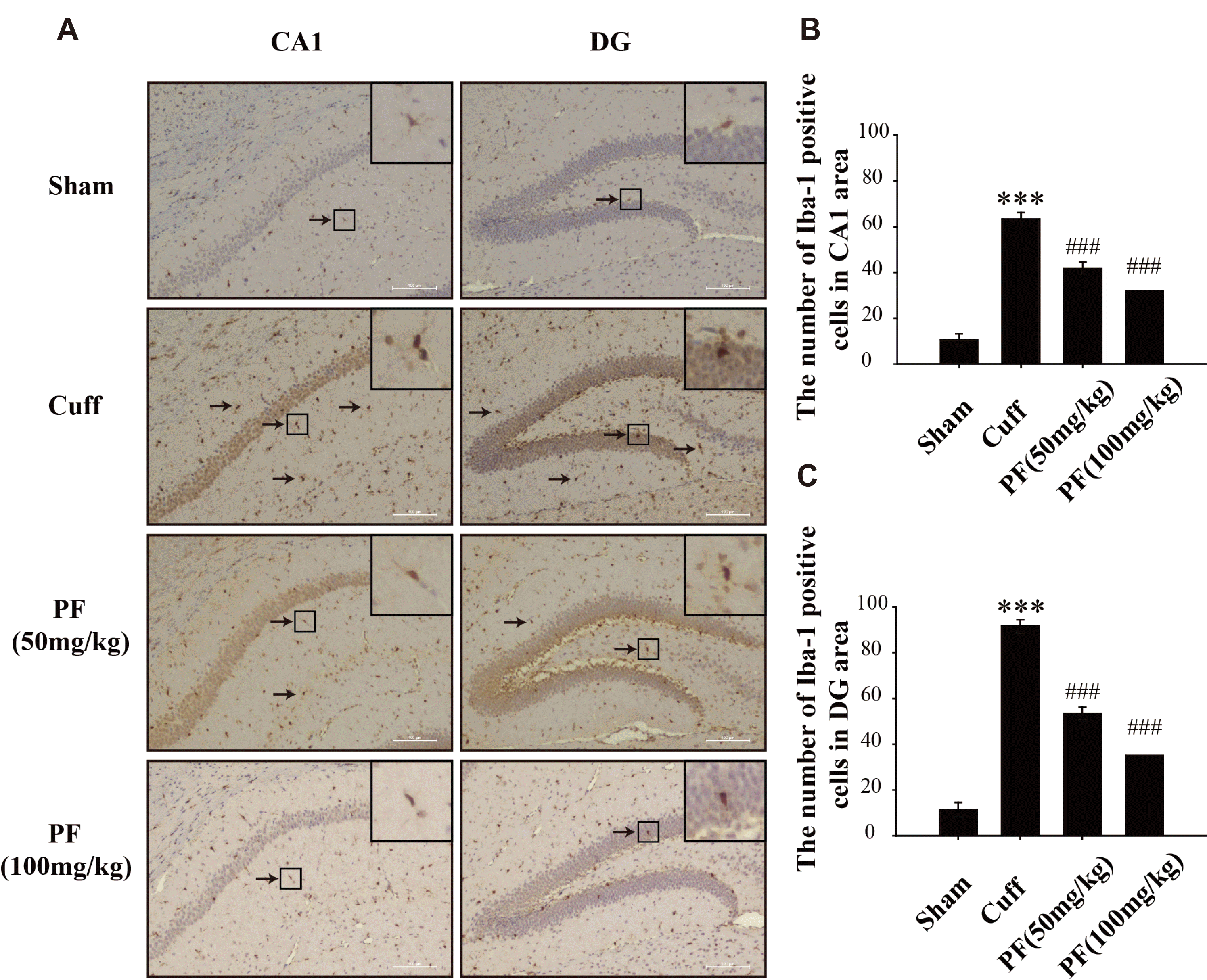 | Fig. 5PF treatment suppressed overactivation of microglia in the CA1 and DG induced by Cuff.Representative images of CA1 and DG (Bar = 100 μm) region sections stained with Iba-1 antibodies (A; black arrows indicate positive microglia). Effect of PF on the number of positive cells for Iba-1 in CA1 (B) and DG (C) regions. Mean ± standard deviation of three experiments. n = 4. PF, paeoniflorin. ***p < 0.001 vs. Sham group; ###p < 0.001 vs. Cuff group.
|
The results of immunohistochemistry were analyzed using Image-ProPlus software. The expression level of Iba-1 in the CA1 and DG regions of the Cuff group significantly increased (p < 0.001) compared with the Sham group (Fig. 5B, C). The expression of Iba-1 was significantly reduced after PF treatment in a dose-dependent manner (p < 0.001, p < 0.001). These results suggest that the activation of hippocampal microglia may be involved in NP-related depression, and PF can inhibit the activation of hippocampal microglia.
Effect of PF on the TLR4/NF-κB pathway in NP mice
Western blot was used to analyze the expression levels of TLR4, MyD88, IκBα, p-NF-κBp65, and NF-κBp65 in the hippocampus. As shown in Fig. 6A–C, the expression levels of TLR4 and MyD88 in Cuff group mice were significantly increased (TLR4, p < 0.001; MyD88, p < 0.001). PF treatment downregulated the expression of (TLR4, p < 0.01, p < 0.01; MyD88, p < 0.01, p < 0.001). NF-κBp65 is a nuclear protein related to the NF-κB signaling pathway. As shown in Fig. 6D, PF inhibited the increase of p-NF-κBp65 expression caused by the Cuff model (p < 0.01, p < 0.001). IκBα is known to inhibit the activation and nuclear translocation of NF-κB molecules, and the production of NF-κBp65 molecules is usually accompanied by the degradation of IκBα molecules [20]. As shown in Fig. 6E, the expression of IκBα was reduced in the Cuff group of mice (p < 0.001). The expression of IκBα was significantly upregulated after PF intervention (p < 0.01, p < 0.001). These results suggest that PF has an inhibitory effect on the TLR4/NF-κB signaling pathway, and the TLR4/NF-κB signaling pathway may be related to the antihyperalgesic and antidepressant‐like effects of PF.
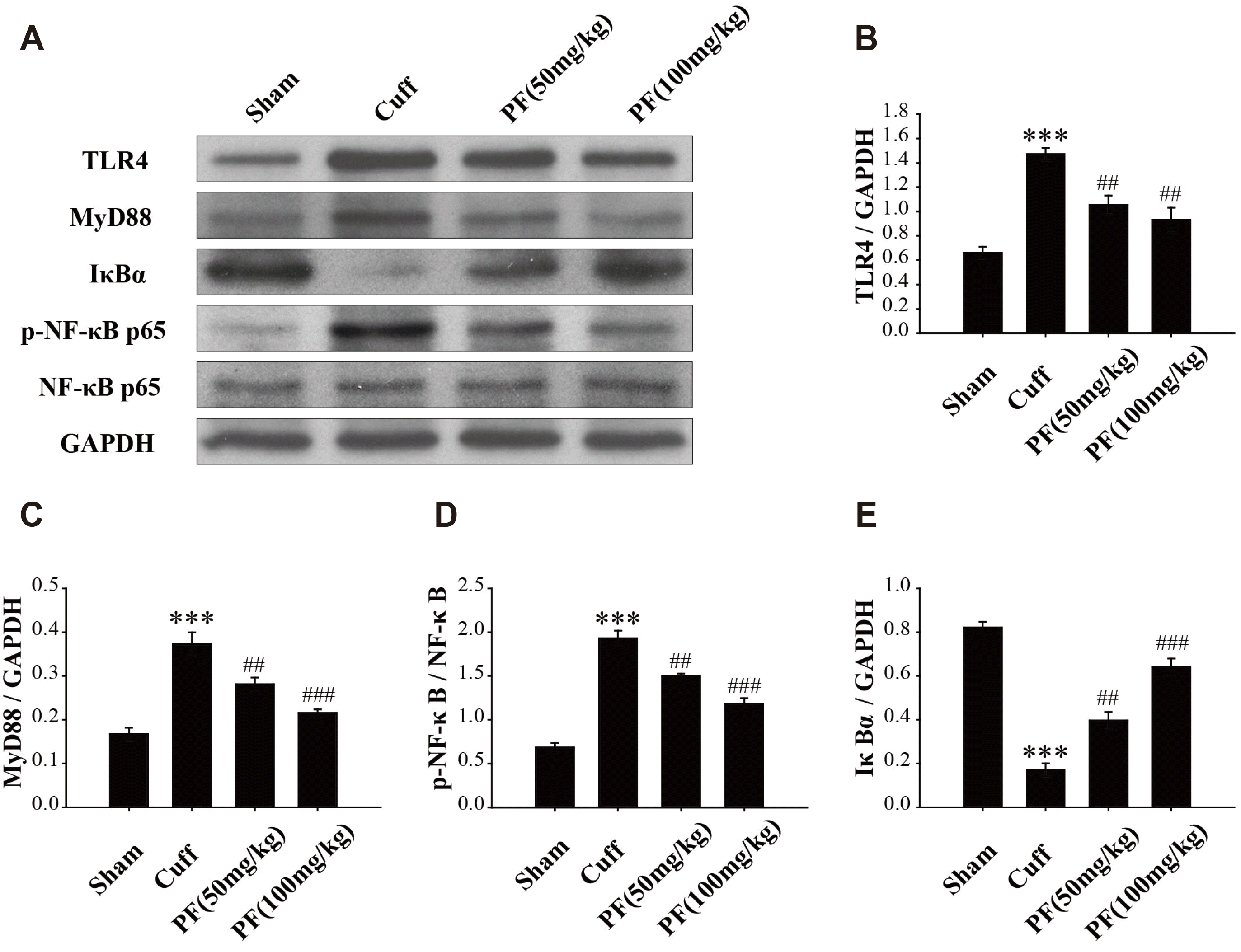 | Fig. 6PF inhibits the increase in expression of TLR4/NF-κB pathway induced by Cuff.The expression of TLR4, MyD88, IκBα, p-NF-κBp65, and NF-κBp65 were determined by western blot analysis in the hippocampus (A). Statistical analysis of relative levels of TLR4 (B), MyD88 (C), p-NF-κBp65 (D), and IκBα (E). Quantitative results are expressed as mean ± standard deviation. n = 3. PF, paeoniflorin. ***p < 0.001 vs. Sham group; ##p < 0.01, ###p < 0.001 vs. Cuff group.
|
Go to :
DISCUSSION
According to epidemiological reports, the prevalence of depression among patients with NP is approximately 34% [21]. Although many drugs have been developed to treat pain and depression, most drugs have multiple adverse effects [22]. Previous studies have shown that PF, as a natural medicine, has the advantage of multiple targets and multiple pathways, and has significant effects in pain relief and anti-depression [23-26]. In this study, intraperitoneal injection of PF for 14 days could attenuate hyperalgesia in mice, suggesting that PF has analgesic effects. After modeling, the chronic pain of mice may interfere with the results of the FST and TST; however, it has little effect on the increase in immobility time in the experiment, whether it was in the SPT, FST, or TST, it was concluded that the depression of mice increased. It should be noted that PF treatment can improve depression-like behavior.
The hippocampus is an important anatomical structure where NP is found to be associated with depression. Pathological studies have shown that hippocampal atrophy is detected in both NP and depressed patients [27-29]. H&E staining of the mouse hippocampus showed a large amount of inflammatory infiltration in the pyramidal cells of the CA3 region after Cuff modeling, and some cells were atrophied or lost. We speculate that this is precisely due to the neuroinflammation that caused changes in the hippocampus after Cuff modeling, which in turn increased depression in mice. PF can improve the damage in the hippocampal structure of the Cuff mice, suggesting that PF has neuroprotective and antidepressant-like effects.
The mechanism of neuroinflammation has been reported in studies of NP and depression [4,30], and the occurrence and development of neuroinflammation are vital in the involvement of pro-inflammatory cytokines. Several clinical studies have found that patients with pain and depression have significantly higher concentrations of pro-inflammatory factors, such as TNF-α and IL-6 in the peripheral blood than those not in pain and non-depressed patients [31,32]. In addition, it has also been found that animal models can manifest depression-like behaviors after receiving intraventricular injection of pro-inflammatory cytokines [33]. The injection of an antagonist to interfere with the inflammatory pathway can simultaneously alleviate hyperalgesia and depressive symptoms in the pain model [34]. Thus, the reduction of pro-inflammatory cytokines may have a therapeutic effect on NP and depression, which allowed us to evaluate the expression of several inflammation-related factors in the hippocampus. We found that PF significantly inhibited the overexpression of TNF-α, IL-6, and IL-1β in the hippocampus of the Cuff group in a dose-dependent manner. We concluded that the analgesic and antidepressant effects of PF may be mediated, at least in part, by inhibiting hippocampal inflammation.
Microglia are widely distributed in the central nervous system and can mediate a series of changes in the process of pain signal transmission. Their role has been widely recognized in mediating pain. Under normal circumstances, microglia are considered to be dormant cells of the central nervous system, which activate rapidly after the nervous system is subjected to noxious stimulation. Activated microglia can release a large amount of neuroactive substances and pro-inflammatory cytokines [35]. These substances and cytokines can act on neurons to enhance the release of nociceptive neurotransmitters from the primary afferent nerve endings and can further activate the glial cells to release more cytokines, forming a positive feedback effect, which causes persistent NP. Previous studies have found that in the NP model, microglia activation participates and promotes pain hypersensitivity [36]. The activation of microglia has also been reported in human studies on depression [37]. It can be seen that abnormal activation of microglia is involved in the pathogenesis of NP and depression. Our results show that hippocampal microglia in Cuff mice are in an activated state, and PF can significantly inhibit the activation of hippocampal microglia.
The TLR4/NF-κB signaling pathway is an important target for inhibiting microglial activation [38]. There are reports of high expression of TLR4 and NF-κB in both pain and depression models [8,9]. First, we detected the expression of TLR4 and its downstream MyD88 in the hippocampus by western blotting; PF inhibited the expression of TLR4 and MyD88 in a dose-dependent manner. IκBα inhibits the transcription factor activity of NF-κB by binding to NF-κB [39]. Once stimulated, IκBα kinase participates in the phosphorylation and ubiquitin-mediated degradation of NF-κB through the proteasome pathway, so that NF-κB can be released and enter the nucleus to perform transcriptional activation, thereby activating the release of various inflammatory factors [40,41]. We continue to explore whether PF inhibits NF-κB phosphorylation and IκBα degradation. Our research shows that PF blocks IκBα degradation in a dose-dependent manner. It effectively inhibits NF-κB phosphorylation, thereby inhibiting the release of inflammatory factors and improving NP and its related depression-like behaviors. In summary, our research proved that PF can reduce hippocampal inflammation by inhibiting the TLR4/NF-κB signaling pathway in the hippocampus and alleviate Cuff-induced NP and depression-like symptoms. This study suggests that PF is a potentially effective treatment option for patients with NP and depression and is worthy of further clinical research.
We believe that neuralgia causes increased expression of TLR4/NF-κB pathway-related proteins, and activation of hippocampal microglia to release pro-inflammatory cytokines which causes hippocampal inflammation leading to the maintenance of NP and production of depression-like behaviors. This study shows that PF can inhibit the hippocampal inflammatory response through the TLR4/NF-κB pathway and display an analgesic and antidepressant effect.
Go to :
 XML Download
XML Download